
Utilisation of Defined Media towards Evaluating Brewing Ale Yeast Fermentation in Small Scale Batches †

 ,
, 
Abstract
:1. Introduction
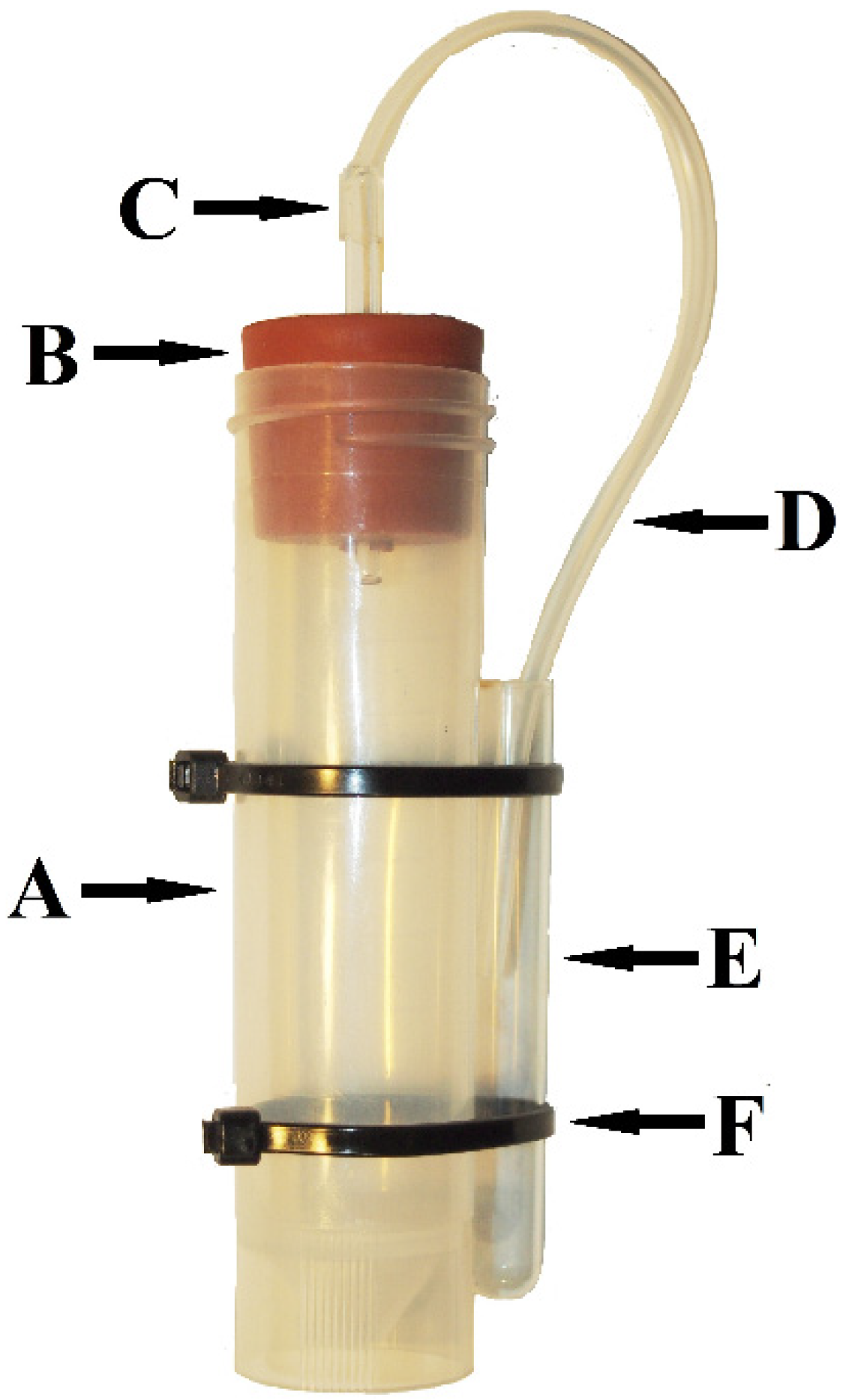
2. Methods
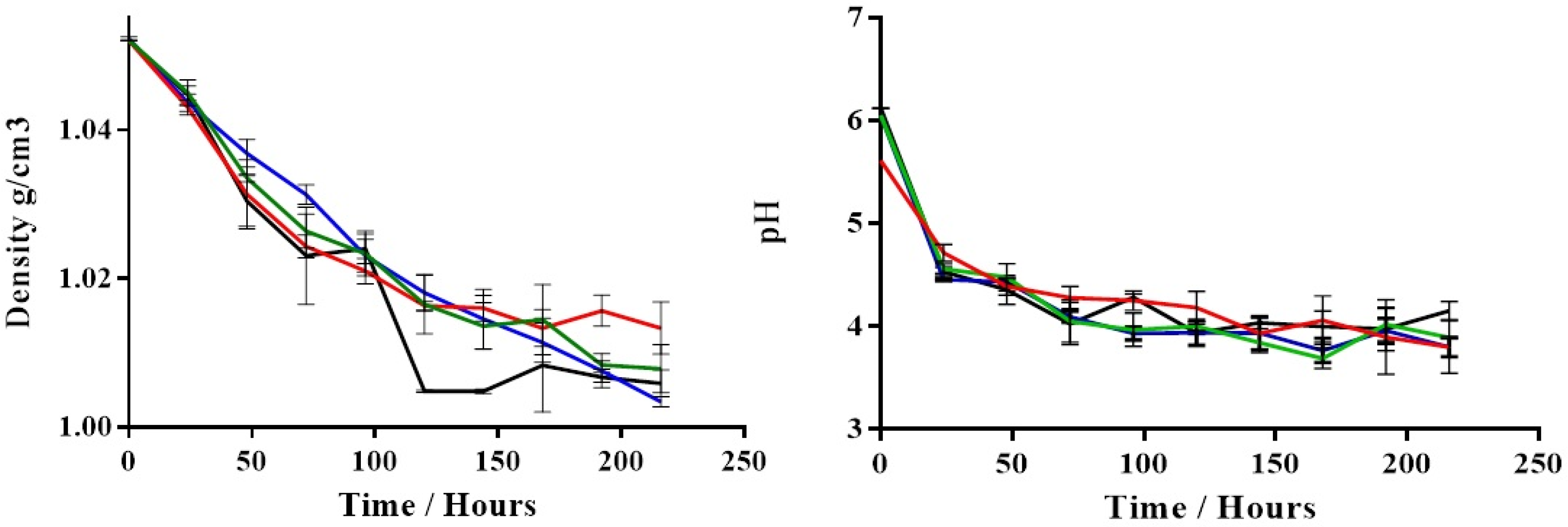
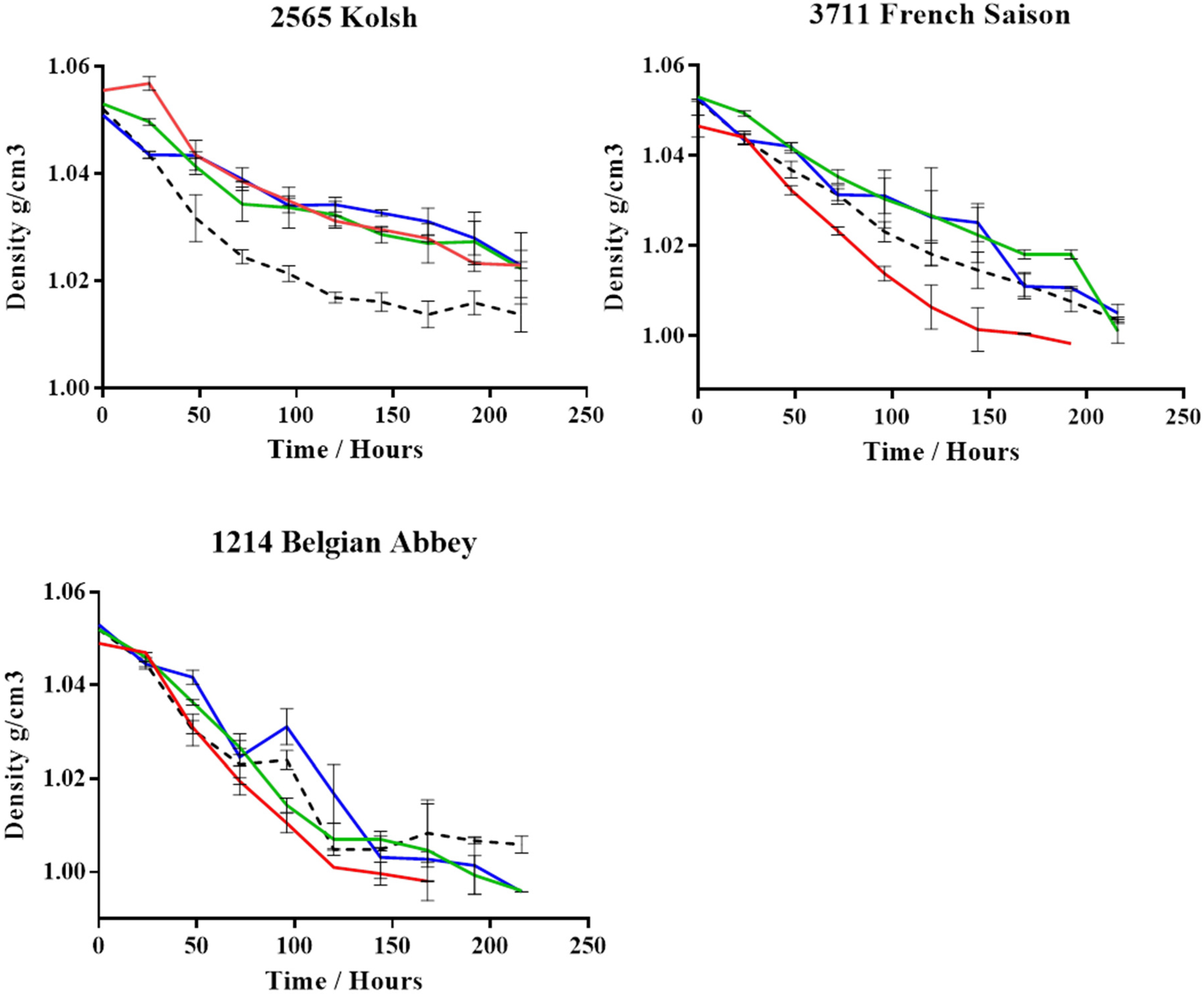
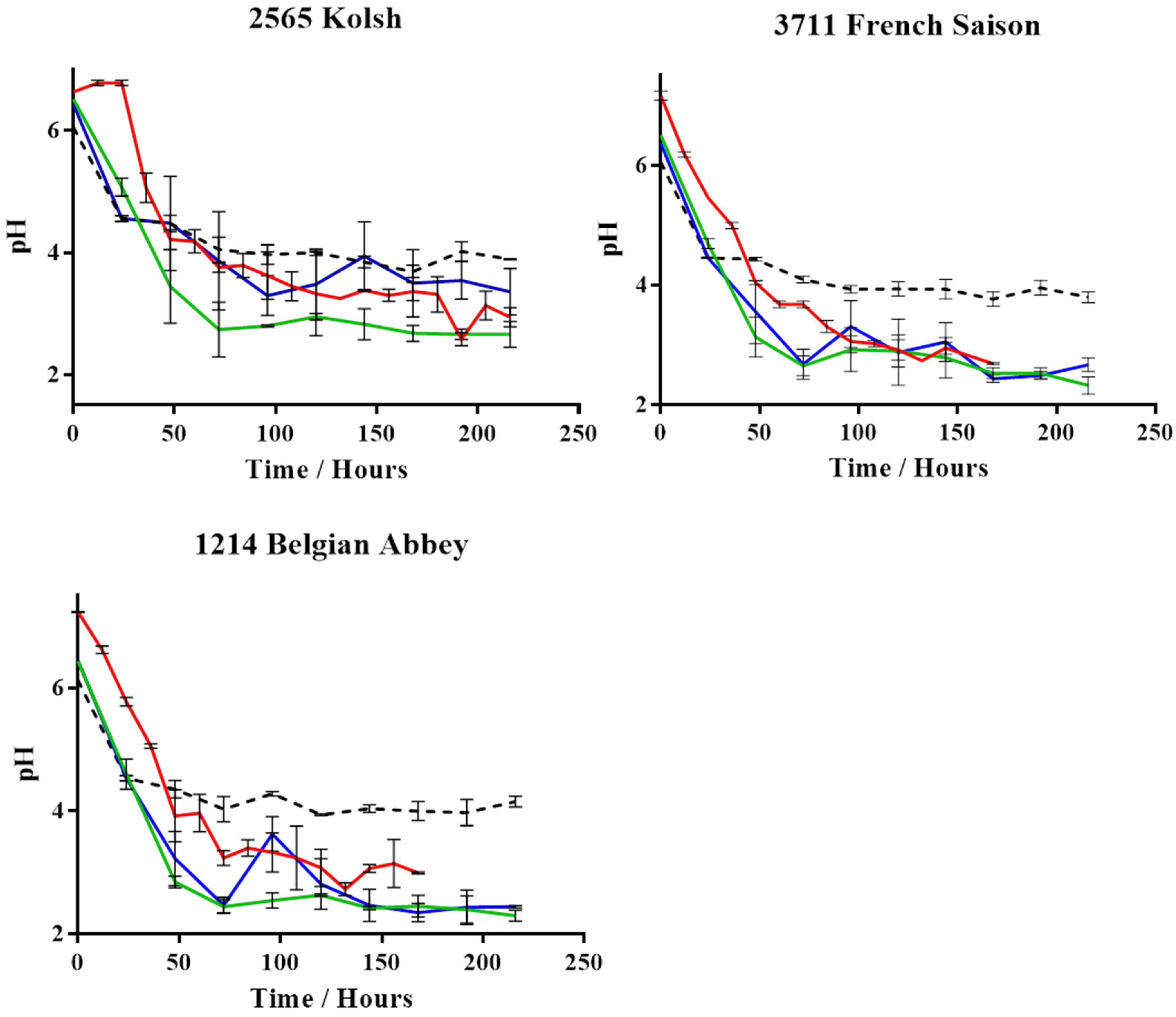
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Recek, N.; Zhou, R.; Zhou, R.; Te’o, V.S.J.; Speight, R.E.; Mozetič, M.; Vesel, A.; Cvelbar, U.; Bazaka, K.; Ostrikov, K.K. Improved fermentation efficiency of S. cerevisiae by changing glycolytic metabolic pathways with plasma agitation. Sci. Rep. 2018, 8, 8252. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, C. Current perspectives on the role of enzymes in brewing. J. Cereal Sci. 2009, 50, 353–357. [Google Scholar] [CrossRef]
- Bamforth, C. Barley and malt Starch in brewing: A general review. Master Brew. Assoc. Am. 2003, 4, 89–97. [Google Scholar]
- MacIntosh, A.J. Carbon Dioxide Generation, Transport and Release during the Fermentation of Barley Malt. Ph.D. Thesis, Dalhousie University, Halifax, NS, Canada, 2013. [Google Scholar]
- Lainioti, G. New Approaches to the Kinetic Study of Alcoholic Fermentation by Chromatographic Techniques. J. Chromatogr. Sci. 2013, 51, 764–779. [Google Scholar] [CrossRef] [PubMed]
- Dutton, J. FV control with real time SG monitoring. Brew. Dist. Int 1990, 21, 20–21. [Google Scholar]
- Leedham, P.A.; Tubb, R.S. Controlling alcohol Fermentations. U.S. Patent US4652451A, 24 March 1987. [Google Scholar]
- Stassi, P.; Goetzke, G.; Fehring, J. Evalution of an insertion thermal mass flowmeter to monitor carbon dioxide evolution rate in large scale fermentations. Tech. Q.-Master Brew. Assoc. Am. 1991, 28, 84–88. [Google Scholar]
- Pfisterer, E.; Krynicki, C.; Steer, J.; Hagg, W. On-line control of ethanol and carbon dioxide in high gravity brewing. Tech. Q.-Master Brew. Assoc. Am. 1988, 25, 1–5. [Google Scholar]
- Evans, D.E.; Hamet, M.A.G. The Selection of a Dried Yeast Strain for Use in the Apparent Attenuation Limit Malt Analysis (AAL) Procedure. J. Inst. Brew. 2005, 111, 209–214. [Google Scholar] [CrossRef]
- Bathgate, G.N. The determination of fermentable extract in brewing worts. J. Inst. Brew. 1981, 87, 301–302. [Google Scholar] [CrossRef]
- Parulekar, S.J.; Birol, G.; Cinar, A.; Undey, C. Batch Fermentation: Modeling Monitoring and Control; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Bamforth, C. Wort Composition and Beer Quality. In Brewing Yeast Fermentation Performance; Smart, K., Ed.; Blackwell Science Ltd.: Hoboken, NJ, USA, 2008; pp. 77–84. [Google Scholar]
- Smart, K. Brewing Yeast Fermentation Performance; Blackwell: Oxford, UK, 2003. [Google Scholar]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort composition and its impact on the flavour-active higher alcohol and ester formation of beer—A review. J. Inst. Brew. 2014, 120, 157–163. [Google Scholar] [CrossRef]
- Bisson, L.F.; Fan, Q.; Walker, G.A. Sugar and Glycerol Transport in Saccharomyces cerevisiae. In Yeast Membrane Transport; Ramos, J., Sychrová, H., Kschischo, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 125–168. [Google Scholar]
- Goldenthal, M.J.; Vanoni, M.; Buchferer, B.; Marmur, J. Regulation of MAL gene expression in yeast: Gene dosage effects. Mol. Gen. Genet. 1987, 209, 508–517. [Google Scholar] [CrossRef]
- Priest, F.; Stewart, G. Handbook of Brewing; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Yu, Z.M.; Zhao, M.M.; Li, H.P.; Zhao, H.F.; Zhang, Q.L.; Wan, C.Y.; Li, H.P. A comparative study on physiological activities of lager and ale brewing yeasts under different gravity conditions. Biotechnol. Bioproc. Eng. 2012, 17, 818–826. [Google Scholar] [CrossRef]
- Casey, G.P.; Magnus, C.A.; Ingledew, W.M. High-Gravity Brewing: Effects of Nutrition on Yeast Composition, Fermentative Ability, and Alcohol Production. Appl. Environ. Microbiol. 1984, 48, 639–646. [Google Scholar] [CrossRef]
- Stewart, G.G. High-Gravity Brewing and Distilling—Past Experiences and Future Prospects. J. Am. Soc. Brew. Chem. 2010, 68, 1–9. [Google Scholar] [CrossRef]
- Saerens, S.; Verbelen, P.; Vanbeneden, N.; Thevelein, J.; Delvaux, F. Monitoring the influence of high-gravity brewing and fermentation temperature on flavour formation by analysis of gene expression levels in brewing yeast. Appl. Microbiol. Biotechnol. 2008, 80, 1039–1051. [Google Scholar] [CrossRef]
- Cooper, D.; Stewart, G.; Bryce, J. Some reasons why high gravity brewing has a negative effect on head retention. J. Inst. Brew. 1998, 104, 83–87. [Google Scholar] [CrossRef]
- Cruber, M.A. The flavor contributions of kilned and roasted products to finished beer styles. Tech. Q.-Master Brew. Assoc. Am. 2001, 38, 223–227. [Google Scholar]
- Psota, V.; Vejrazka, K.; Famera, O.; Hrcka, M. Relationship between grain hardness and malting quality of barley (Hordeum vulgare L.). J. Inst. Brew. 2007, 113, 80–86. [Google Scholar] [CrossRef]
- Wainwright, T.; Slack, P.T.; Long, D.E. N-nitrosodimethylamine precursors in malt. IARC Sci. Publ. 1982, 41, 71–80. [Google Scholar]
- Slack, P.T.; Baxter, E.D.; Wainwright, T. Inhibition by Hordein of Starch Degradation. J. Inst. Brew. 1979, 85, 112–114. [Google Scholar] [CrossRef]
- Chandra, G.S.; Proudlove, M.O.; Baxter, E.D. The structure of barley endosperm—An important determinant of malt modification. J. Sci. Food Agric. 1999, 79, 37–46. [Google Scholar] [CrossRef]
- Gupta, M.; Abu-Ghannam, N.; Gallaghar, E. Barley for Brewing: Characteristic Changes during Malting, Brewing and Applications of its By-Products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 318–328. [Google Scholar] [CrossRef]
- Taidi, B.; Mina, M.; Hodgson, J. Development of an artificial wort for yeast fermentation studies. In Proceedings of the ASBC Meeting, Victoria, BC, Canada, 24–27 June 2001. [Google Scholar]
- Taidi, B.; Kennedy, A.; Hodgson, J. Wort Substitutes and Yeast Nutrition. In Brewing Yeast Fermentation Performance, 2nd ed.; Smart, K., Ed.; Wiley: New York, NY, USA, 2003; pp. 86–95. [Google Scholar]
- Lake, J.C.; Speers, R.A.; Porter, A.V.; Gill, T.A. Miniaturizing the fermentatation assay: Effects of fermentor size and fermentation kinetics on detection of premature yeast flocculation. J. Am. Soc. Brew. Chem. 2008, 66, 94–102. [Google Scholar] [CrossRef]
- ASBC. Methods of Analysis Yeast-14 Miniature Fermentation Assay; ASBC: Saint Paul, MN, USA, 2011. [Google Scholar]
- Piddocke, M.P. Physiological characterization of brewer’s yeast in high-gravity beer fermentations with glucose or maltose syrups as adjuncts. Appl. Microbiol. Biotechnol. 2009, 84, 453–465. [Google Scholar] [CrossRef]
- Gąsior, J.; Kawa-Rygielska, J.; Kucharska, A.Z. Carbohydrates profile, polyphenols content and antioxidative properties of beer worts produced with different dark malts varieties or roasted barley grains. Molecules 2020, 25, 3882. [Google Scholar] [CrossRef]
- Rakete, S.; Glomb, M.A. A Novel Approach for the Quantitation of Carbohydrates in Mash, Wort, and Beer with RP-HPLC Using 1-Naphthylamine for Precolumn Derivatization. J. Agric. Food Chem. 2013, 61, 3828–3833. [Google Scholar] [CrossRef]
- Garza-Ulloa, H.; Cantú, R.G.; Gajá, A.M.C. Determination of Amino Acids in Wort and Beer by Reverse-Phase High-Performance Liquid Chromatography. J. Am. Soc. Brew. Chem. 1986, 44, 47–51. [Google Scholar] [CrossRef]
- Punčochářová, L.; Pořízka, J.; Diviš, P.; Štursa, V. Study of the influence of brewing water on selected analytes in beer. Potravin. Slovak J. Food Sci. 2019, 13, 507–514. [Google Scholar] [CrossRef]
- MacWilliam, I.C. Wort composition—A review. J. Inst. Brew. 1968, 74, 38–54. [Google Scholar] [CrossRef]
- Graham, R.K.; Skurray, G.R.; Caiger, P. Nutritional studies on yeasts during batch and continuous fermentation. I. Changes in vitamin concentrations. J. Inst. Brew. 1970, 76, 366–371. [Google Scholar] [CrossRef]
- EBC. Hot Water Extract of Malt: Constant Temperature Mash—Section 4 Malt Method 4.6. In Analytica EBC; BrewUp: Brussels, Belgium, 1997. [Google Scholar]
- Soares, E.V. Flocculation in Saccharomyces cerevisiae: A review. J. Appl. Microbiol. 2011, 110, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, C. pH in brewing: An overview. Tech. Q.-Master Brew. Assoc. Am. 2001, 38, 1–9. [Google Scholar]
- Salmon, J.M. Effect of Sugar Transport Inactivation in Saccharomyces cerevisiae on Sluggish and Stuck Enological Fermentations. Appl. Environ. Microbiol. 1989, 55, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Salmon, J.M.; Barre, P. Assimilable nitrogen addition and hexose transport system activity during enological fermentation. J. Inst. Brew. 1994, 100, 279–282. [Google Scholar] [CrossRef]
- Nikulin, J.; Vidgren, V.; Krogerus, K.; Magalhães, F.; Valkeemäki, S.; Kangas-Heiska, T.; Gibson, B. Brewing potential of the wild yeast species Saccharomyces paradoxus. Eur. Food Res. Technol. 2020, 246, 2283–2297. [Google Scholar] [CrossRef]
- Fox, R.L.; Logue, S.J.; Eglinton, J.K. Fermentable sugar profile as an alternative to apparent attenuation limit for selection in barley breeding. In Proceedings of the 10th Australian Barley Technical Symposium, Canberra, Australia, 16–20 September 2001. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration | Concentration Used by Taidi et al. [31] | Typical Range in Natural Wort |
|---|---|---|---|
| Carbohydrates | |||
| Glucose (g L−1) | 12 | 10.4 | 0.47–6.48 |
| Fructose (g L−1) | 8 | 4.6 | ND |
| Sucrose (g L−1) | 0.4 | 3.5 | ND |
| Maltose (g L−1) | 115 | 115.5 | 34.74–51.69 |
| Maltotriose (g L−1) | 5.75 | Not specified * | 9.13–13.87 |
| Dextrin (g L−1) | 0 | 0 | 18.03–39.85 |
| Buffer | |||
| Potassium phosphate dibasic (K2HPO4) (g L−1) | 2 g L−1 | 0 (used 0.625 g L−1 citric acid & 0.215 g L−1 CaSO4∙2H2O) | ND |
| pH | 6.2 | 5.2 | 5.2–5.4 |
| Yeast Nitrogen Base | |||
| Ammonium sulphate (g L−1) | 5 | 0.1307 | ND |
| Monopotassium phosphate (g L−1) | 1 | 0 | ND |
| Magnesium sulphate (g L−1) | 0.5 | 0 | ND |
| Sodium chloride (g L−1) | 0.1 | 0 | ND |
| Calcium chloride (g L−1) | 0.1 | 0 | ND |
| Boric acid (µg L−1) | 400 | 0 | ND |
| Manganese sulphate (µg L−1) | 400 | 0 | ND |
| Zinc sulphate (µg L−1) | 400 | 0 | ND |
| Ferric chloride (µg L−1) | 200 | 0 | ND |
| Sodium molybdate (µg L−1) | 200 | 0 | ND |
| Potassium iodide (µg L−1) | 100 | 0 | ND |
| Copper sulphate (µg L−1) | 40 | 0 | ND |
| L-histidine monohydrochloride (mg L−1) | 10 | 0 | ND |
| L-D-methionine (mg L−1) | 20 | 0 | ND |
| L-D-tryptophan (mg L−1) | 2 | 0 | ND |
| Inositol (mg L−1) | 0.5 | 0 | 18–60 |
| Niacin (mg L−1) | 0.4 | 0 | 8–41.36 |
| Pyridoxine HCl (µg L−1) | 400 | 0 | 580–1005 |
| Thiamine HCl (µg L−1) | 400 | 0 | 280–600 |
| Calcium pantothenate (µg L−1) | 400 | 0 | 450–980 |
| Riboflavin (µg L−1) | 400 | 0 | 400–930 |
| p-aminobenzoic acid (µg L−1) | 200 | 0 | 19–96 |
| Folic acid (µg L−1) | 2 | 0 | 100–130 |
| Biotin (µg L−1) | 2 | 0 | 0.85–1.15 |
| Yeast Strain | Media | AAL (%) | RAL (%) |
|---|---|---|---|
| 2565 Kolsh | Wort | 81 ± 7 | 58 ± 7 |
| Defined | 58 ± 7 | 47 ± 3 | |
| T-test | NS | NS | |
| 3711 French Saison | Wort | 98 ± 7 | 80 ± 5 |
| Defined | 98 ± 6 | 80 ± 6 | |
| T-test | NS | NS | |
| 1214 Belgian Abbey | Wort | 92 ± 2 | 76 ± 2 |
| Defined | 107 ± 3 | 88 ± 2 | |
| T-test | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, J.B.; Portman, D.; Batley, R.; Lal, P.; Bean, D.; Aldred, P.; Naiker, M. Utilisation of Defined Media towards Evaluating Brewing Ale Yeast Fermentation in Small Scale Batches. Biol. Life Sci. Forum 2022, 18, 3. https://doi.org/10.3390/Foods2022-12990
Johnson JB, Portman D, Batley R, Lal P, Bean D, Aldred P, Naiker M. Utilisation of Defined Media towards Evaluating Brewing Ale Yeast Fermentation in Small Scale Batches. Biology and Life Sciences Forum. 2022; 18(1):3. https://doi.org/10.3390/Foods2022-12990
Chicago/Turabian StyleJohnson, Joel B., Drew Portman, Ryan Batley, Pawan Lal, David Bean, Peter Aldred, and Mani Naiker. 2022. "Utilisation of Defined Media towards Evaluating Brewing Ale Yeast Fermentation in Small Scale Batches" Biology and Life Sciences Forum 18, no. 1: 3. https://doi.org/10.3390/Foods2022-12990