
Humidity and Light Modulate Oxygen-Induced Viability Loss in Dehydrated Haematococcus lacustris Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae Cultivation, Dehydration and Rehydration
2.2. Differential Scanning Calorimetry
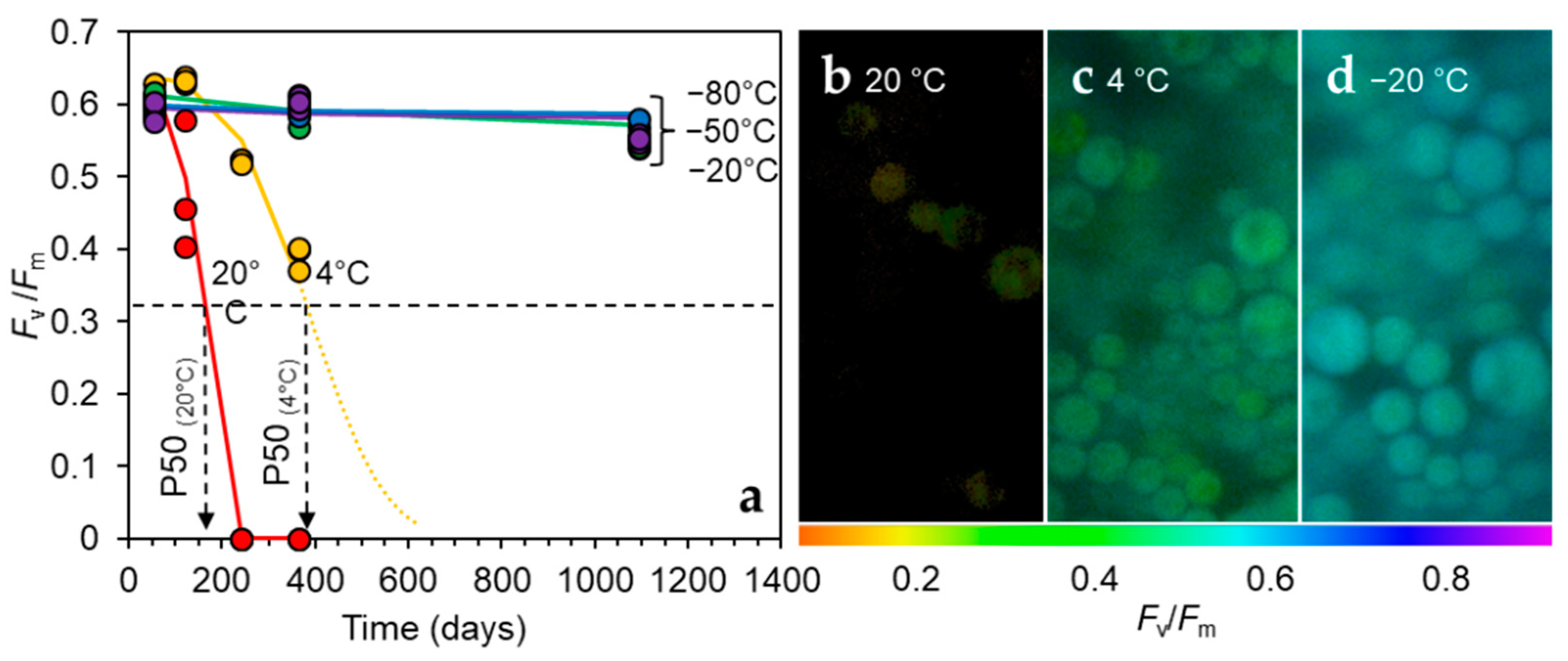
2.3. Long-Term Storage at Various Temperatures and 50% RH in the Dark
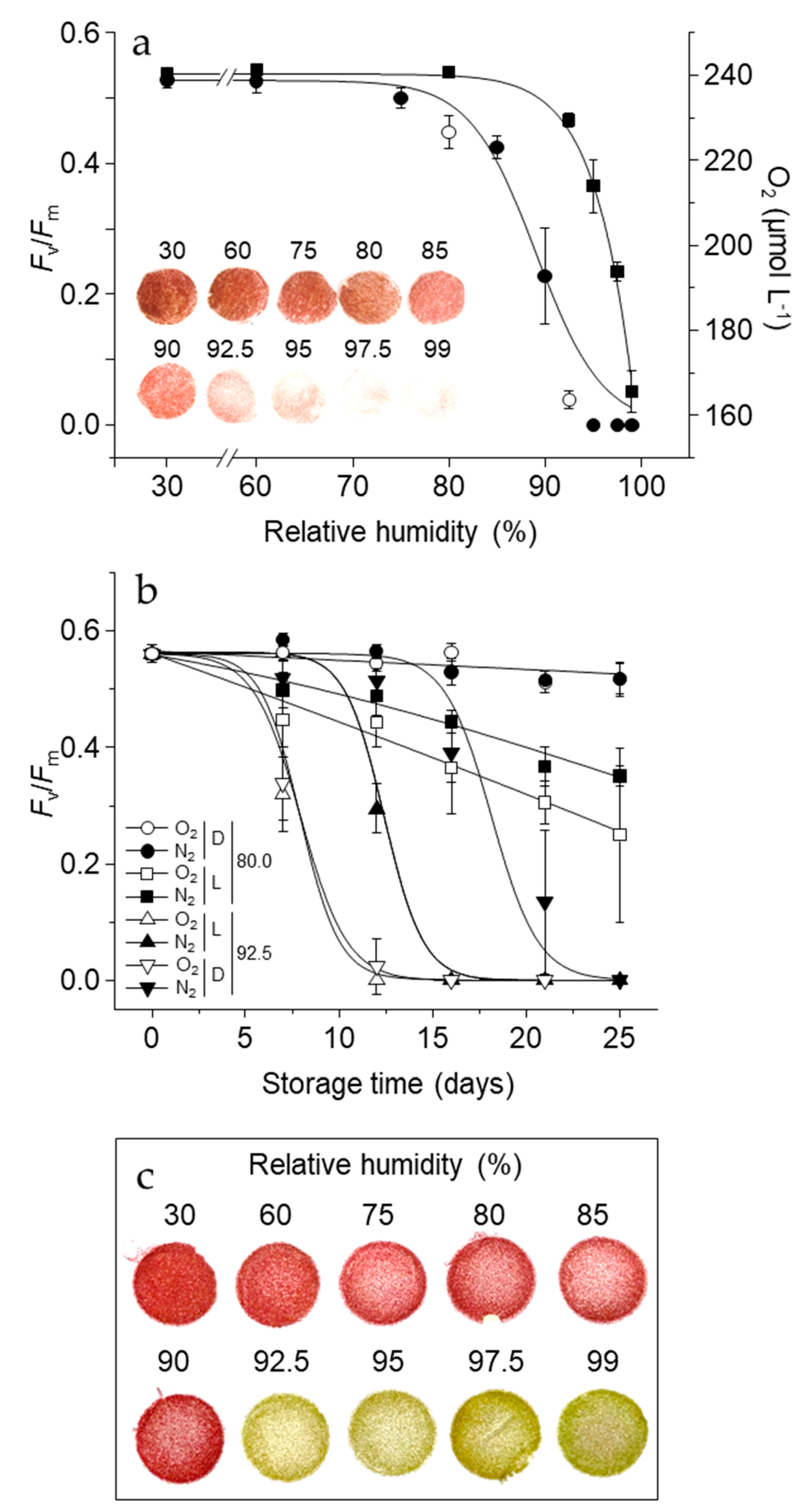
2.4. Short-Term Storage at 21 °C, Various RH, in Light or Dark and Contrasting Oxygen Tension
2.5. Sample Preparation for Biochemical Analyses
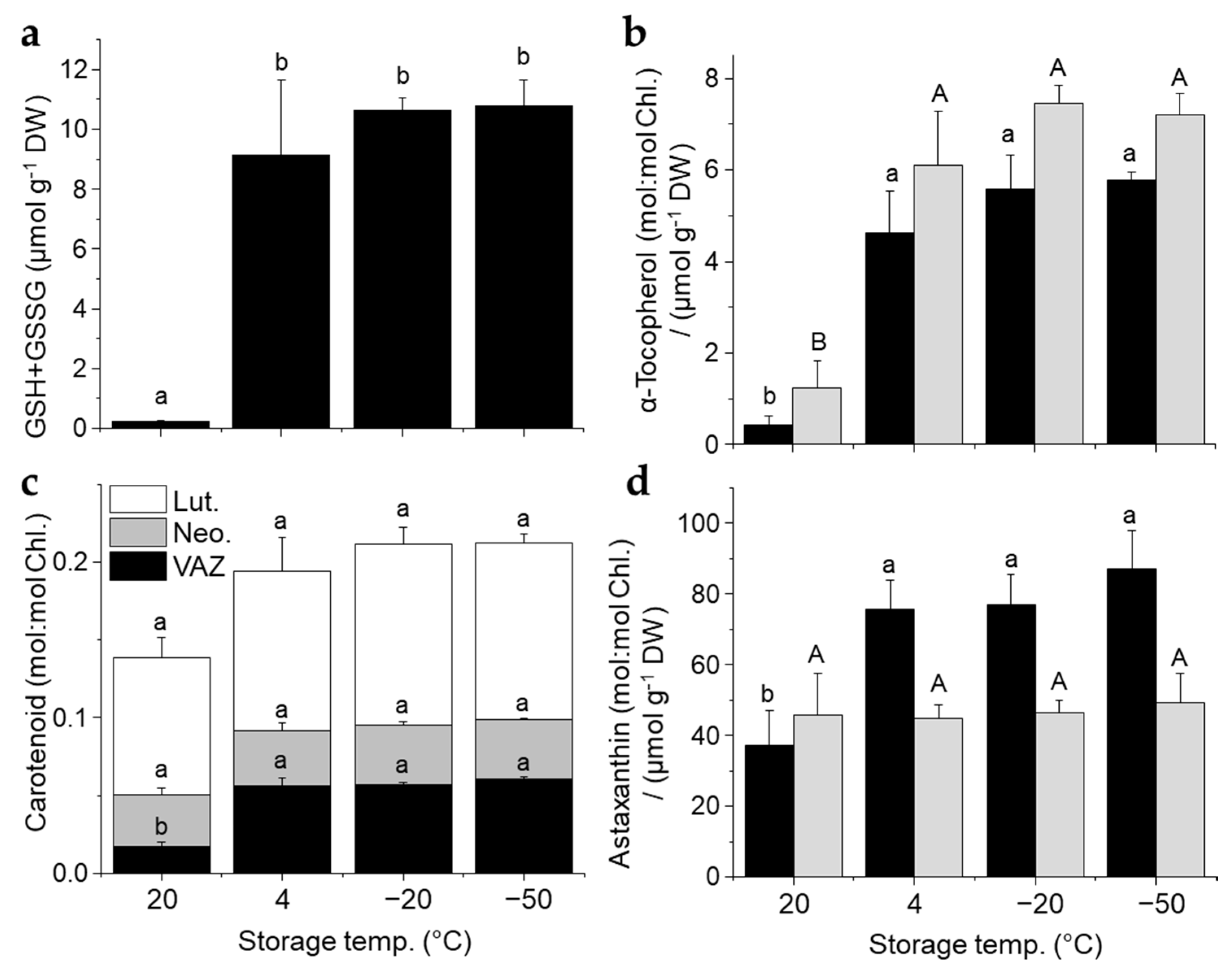
2.6. HPLC Analyses of Pigments, Tocopherols and Glutathione
2.7. Statistical Analyses of Biochemical Measurements
2.8. Chlorophyll Fluorescence Analyses
3. Results
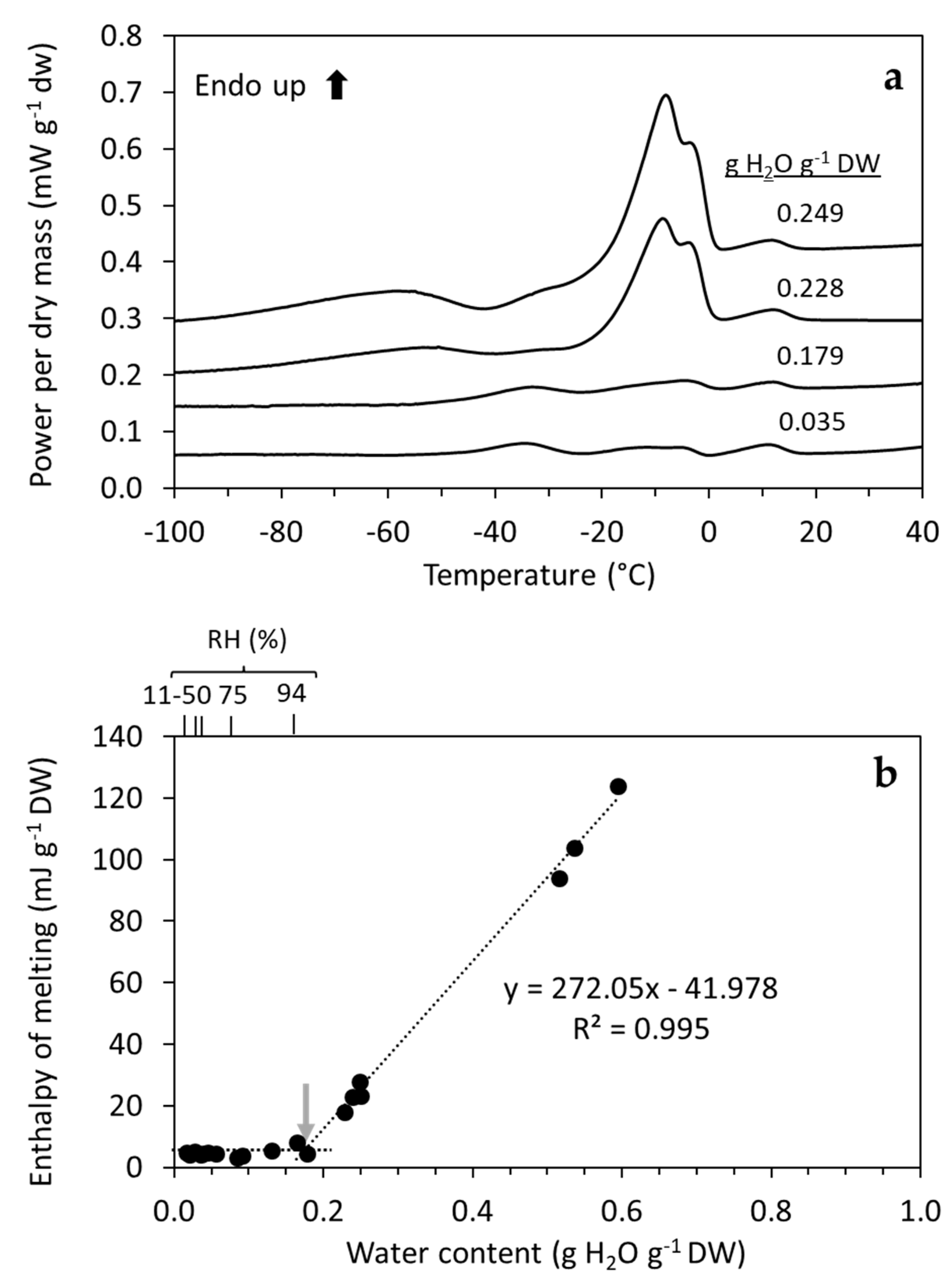
3.1. Physicochemical Status of H. lacustris Cells Based on DSC
3.2. Temperature Dependency of Ageing Reactions in Desiccated H. lacustris Cells
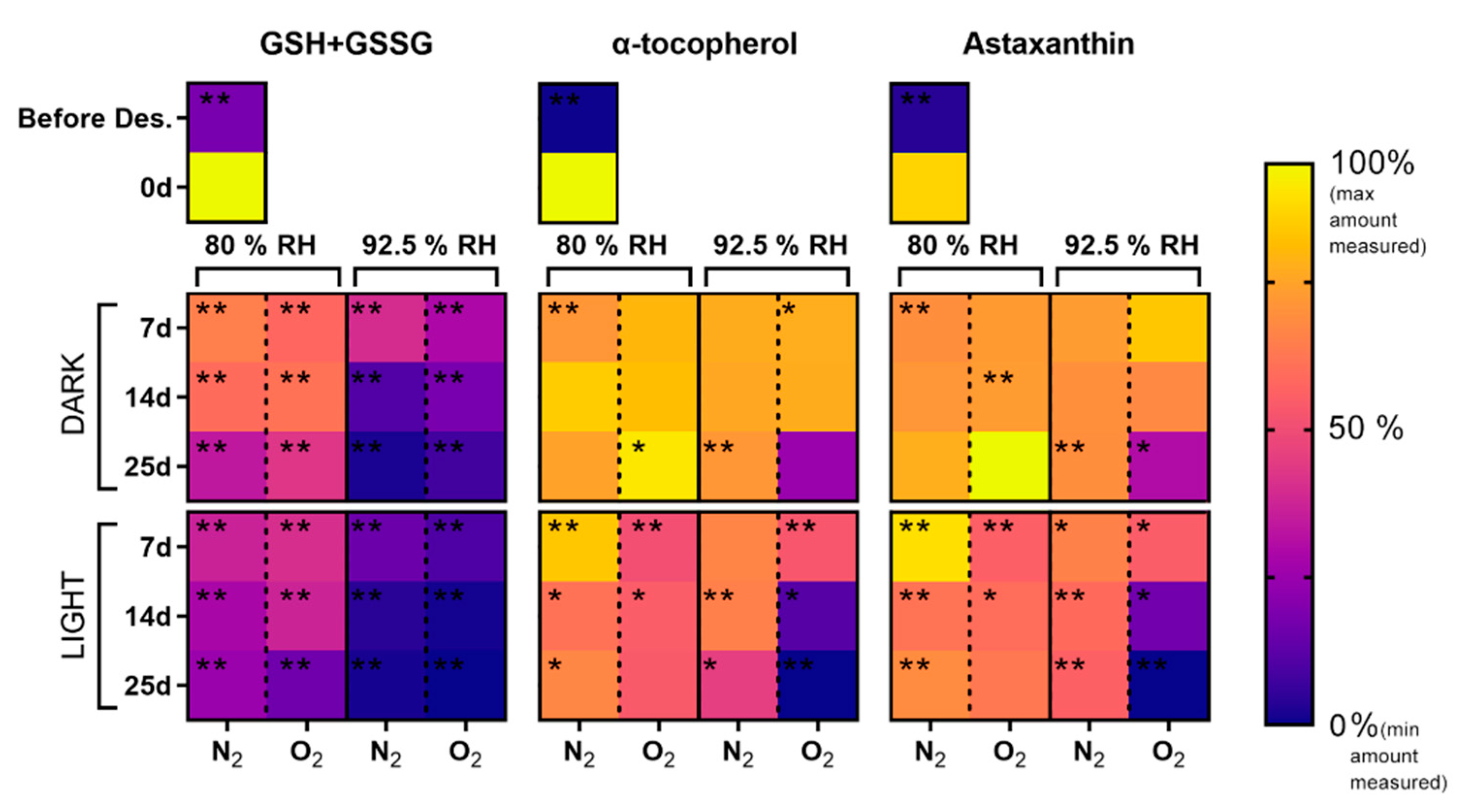
3.3. Oxygen and Light-Dependent Effects on Viability Loss, Pigment Bleaching, and Antioxidant Metabolism at High RH and Contrasting Physicochemical Status
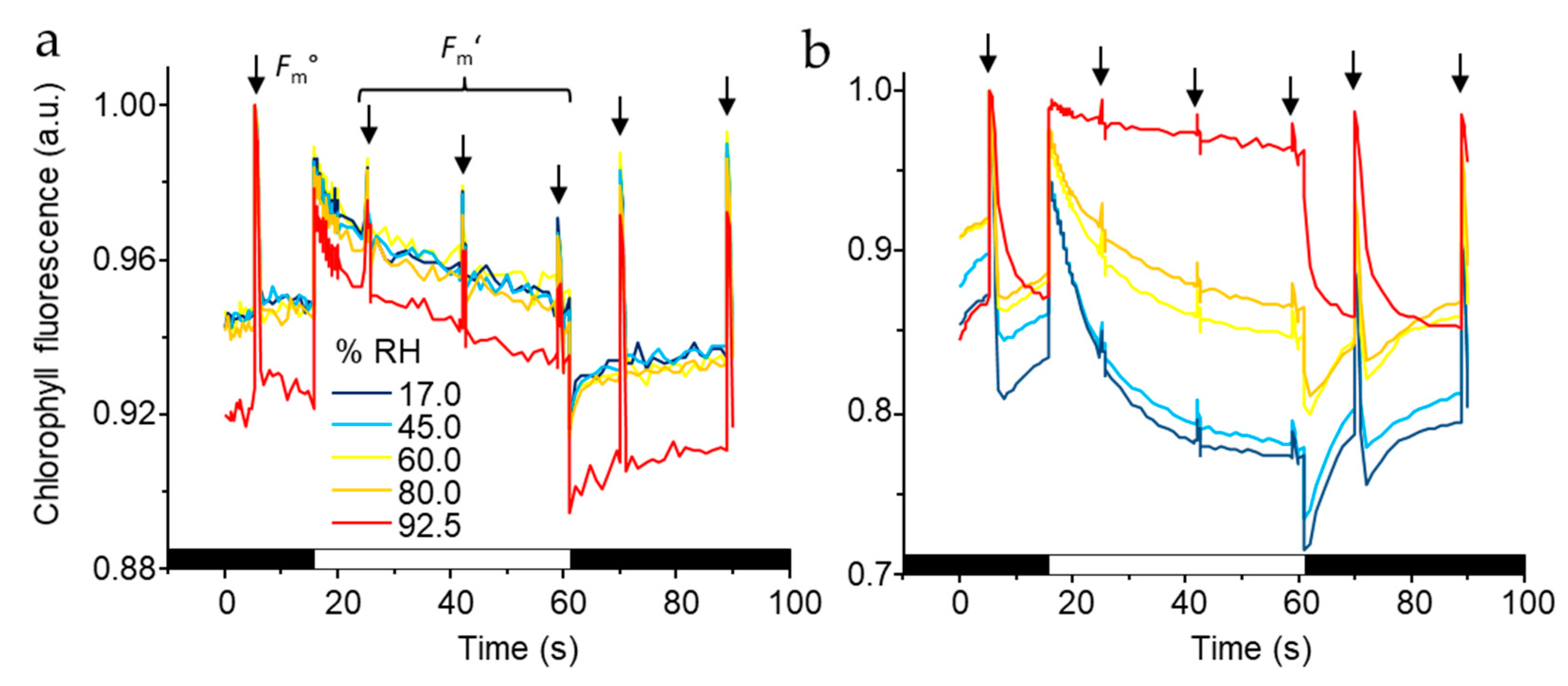
3.4. PSII Activity in Dehydrated Cells Equilibrated to Contrasting RH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellissent-Funel, M.C.; Hassanali, A.; Havenith, M.; Henchman, R.; Pohl, P.; Sterpone, F.; van der Spoel, D.; Xu, Y.; Garcia, A.E. Water determines the structure and dynamics of proteins. Chem. Rev. 2016, 116, 7673–7697. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, D.; Walters, C. Detailed characterization of mechanical properties and molecular mobility within dry seed glasses: Relevance to the physiology of dry biological systems. Plant J. 2011, 68, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Marin, B.; Kranner, I.; San Sebastian, M.; Artetxe, U.; Laza, J.M.; Vilas, J.L.; Pritchard, H.W.; Nadajaran, J.; Miguez, F.; Becerril, J.M.; et al. Evidence for the absence of enzymatic reactions in the glassy state. A case study of xanthophyll cycle pigments in the desiccation-tolerant moss Syntrichia ruralis. J. Exp. Bot. 2013, 64, 3033–3043. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation tolerance: Avoiding cellular damage during drying and rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, C.; Hill, L.M.; Wheeler, L.J. Dying while dry: Kinetics and mechanisms of deterioration in desiccated organisms. Integr. Comp. Biol. 2005, 45, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Leprince, O.; Harren, F.J.; Buitink, J.; Alberda, M.; Hoekstra, F.A. Metabolic dysfunction and unabated respiration precede the loss of membrane integrity during dehydration of germinating radicles. Plant Physiol. 2000, 122, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Buitink, J.; Claessens, M.M.; Hemminga, M.A.; Hoekstra, F.A. Influence of water content and temperature on molecular mobility and intracellular glasses in seeds and pollen. Plant Physiol. 1998, 118, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Kranner, I.; Birtic, S. A modulating role for antioxidants in desiccation tolerance. Integr. Comp. Biol. 2005, 45, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Gerna, D.; Ballesteros, D.; Arc, E.; Stöggl, W.; Seal, C.E.; Marami-Zonouz, N.; Na, C.S.; Kranner, I.; Roach, T. Does oxygen affect ageing mechanisms of Pinus densiflora seeds? A matter of cytoplasmic physical state. J. Exp. Bot. 2022, 73, 2631–2649. [Google Scholar] [CrossRef]
- Roach, T.; Krieger-Liszkay, A. Photosynthetic regulatory mechanisms for efficiency and prevention of photo-oxidative stress. In Annual Plant Reviews Online; Roberts, J.A., Ed.; Wiley Publishing: Hoboken, NJ, USA, 2019; pp. 273–306. [Google Scholar]
- Heber, U.; Bilger, W.; Türk, R.; Lange, O.L. Photoprotection of reaction centres in photosynthetic organisms: Mechanisms of thermal energy dissipation in desiccated thalli of the lichen Lobaria pulmonaria. New Phytol. 2010, 185, 459–470. [Google Scholar] [CrossRef]
- Challabathula, D.; Zhang, Q.; Bartels, D. Protection of photosynthesis in desiccation-tolerant resurrection plants. J. Plant Physiol. 2018, 227, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Farrant, J.M.; Vander Willigen, C.; Loffell, D.A.; Bartsch, S.; Whittaker, A. An investigation into the role of light during desiccation of three angiosperm resurrection plants. Plant Cell Environ. 2003, 26, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Holzinger, A.; Karsten, U. Desiccation stress and tolerance in green algae: Consequences for ultrastructure, physiological and molecular mechanisms. Front. Plant Sci. 2013, 4, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roqueiro, G.; Facorro, G.B.; Huarte, M.G.; Rubin de Celis, E.; Garcia, F.; Maldonado, S.; Maroder, H. Effects of photooxidation on membrane integrity in Salix nigra seeds. Ann. Bot. 2010, 105, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros, D.; Narayan, S.; Varghese, B.; Sershen. Photo-oxidation modulates green fern spore longevity during dry storage. Plant Cell Tissue Organ Cult. 2018, 133, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.M.; Hendry, G.A.F.; Atherton, N.M.; Vertucci-Walters, C.W. Free radical accumulation and lipid peroxidation in testas of rapidly aged soybean seeds: A light-promoted process. Seed Sci. Res. 1996, 6, 101–107. [Google Scholar] [CrossRef]
- Farrant, J.M. A comparison of mechanisms of desiccation tolerance among three angiosperm resurrection plant species. Plant Ecol. 2000, 151, 29–39. [Google Scholar] [CrossRef]
- Rebecchi, L. Dry up and survive: The role of antioxidant defences in anhydrobiotic organisms. J. Limnol. 2013, 72, e8. [Google Scholar] [CrossRef] [Green Version]
- França, M.B.; Panek, A.D.; Eleutherio, E.C.A. Oxidative stress and its effects during dehydration. Comp. Biochem. Physiol.-Part A Mol. Integr. Physiol. 2007, 146, 621–631. [Google Scholar] [CrossRef]
- Erkut, C.; Vasilj, A.; Boland, S.; Habermann, B.; Shevchenko, A.; Kurzchalia, T.V. Molecular strategies of the caenorhabditis Elegans dauer larva to survive extreme desiccation. PLoS ONE 2013, 8, e82473. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Antioxidative response of ascorbate-glutathione pathway enzymes and metabolites to desiccation of recalcitrant Acer saccharinum seeds. J. Plant Physiol. 2006, 163, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Böck, N.; Rittmeier, N.; Arc, E.; Kranner, I.; Holzinger, A. Acquisition of desiccation tolerance in Haematococcus pluvialis requires photosynthesis and coincides with lipid and astaxanthin accumulation. Algal Res. 2022, 64, 102699. [Google Scholar] [CrossRef]
- Kranner, I.; Birtic, S.; Anderson, K.M.; Pritchard, H.W. Glutathione half-cell reduction potential: A universal stress marker and modulator of programmed cell death? Free Radic. Biol. Med. 2006, 40, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Beckett, R.P.; Minibayeva, F.V.; Colville, L.; Whitaker, C.; Chen, H.; Bailly, C.; Kranner, I. Extracellular superoxide production, viability and redox poise in response to desiccation in recalcitrant Castanea sativa seeds. Plant Cell Environ. 2010, 33, 59–75. [Google Scholar] [PubMed]
- Stegner, M.; Wagner, J.; Roach, T. Antioxidant depletion during seed storage under ambient conditions. Seed Sci. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Roach, T.; Nagel, M.; Börner, A.; Eberle, C.; Kranner, I. Changes in tocochromanols and glutathione reveal differences in the mechanisms of seed ageing under seedbank conditions and controlled deterioration in barley. Environ. Exp. Bot. 2018, 156, 8–15. [Google Scholar] [CrossRef]
- Groot, S.P.C.; van Litsenburg, M.-J.; Kodde, J.; Hall, R.D.; de Vos, R.C.H.; Mumm, R. Analyses of metabolic activity in peanuts under hermetic storage at different relative humidity levels. Food Chem. 2022, 373, 131020. [Google Scholar] [CrossRef] [PubMed]
- Morscher, F.; Kranner, I.; Arc, E.; Bailly, C.; Roach, T. Glutathione redox state, tocochromanols, fatty acids, antioxidant enzymes and protein carbonylation in sunflower seed embryos associated with after-ripening and ageing. Ann. Bot. 2015, 116, 669–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schausberger, C.; Roach, T.; Stoggl, W.; Arc, E.; Finch-Savage, W.E.; Kranner, I. Abscisic acid-determined seed vigour differences do not influence redox regulation during ageing. Biochem. J. 2019, 476, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Seal, C.E.; Zammit, R.; Scott, P.; Flowers, T.J.; Kranner, I. Glutathione half-cell reduction potential and α-tocopherol as viability markers during the prolonged storage of Suaeda maritima seeds. Seed Sci. Res. 2010, 20, 47–53. [Google Scholar] [CrossRef]
- Stewart, R.R.C.; Bewley, J.D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Dickie, J.B.; Ellis, R.H.; Kraak, H.L.; Ryder, K.; Tompsett, P.B. Temperature and seed storage longevity. Ann. Bot. 1990, 65, 197–204. [Google Scholar] [CrossRef]
- Ballesteros, D.; Hill, L.M.; Lynch, R.T.; Pritchard, H.W.; Walters, C. Longevity of preserved germplasm: The temperature dependency of aging reactions in glassy matrices of dried fern spores. Plant Cell Physiol. 2019, 60, 376–392. [Google Scholar] [CrossRef]
- Walters, C. Orthodoxy, recalcitrance and in-between: Describing variation in seed storage characteristics using threshold responses to water loss. Planta 2015, 242, 397–406. [Google Scholar] [CrossRef]
- Crane, J.; Kovach, D.; Gardner, C.; Walters, C. Triacylglycerol phase and ‘intermediate’seed storage physiology: A study of cuphea carthagenensis. Planta 2006, 223, 1081–1089. [Google Scholar] [CrossRef]
- Hamilton, K.N.; Ashmore, S.E.; Pritchard, H.W. Thermal analysis and cryopreservation of seeds of Australian wild citrus species (rutaceae): Citrus australasica, C. inodora and C. garrawayi. CryoLetters 2009, 30, 268–279. [Google Scholar] [PubMed]
- Mira, S.; Nadarajan, J.; Liu, U.; González-Benito, M.E.; Pritchard, H.W. Lipid thermal fingerprints of long-term stored seeds of brassicaceae. Plants 2019, 8, 414. [Google Scholar] [CrossRef] [Green Version]
- Boussiba, S. Carotenogenesis in the green alga haematococcus pluvialis: Cellular physiology and stress response. Physiol. Plant. 2000, 108, 111–117. [Google Scholar] [CrossRef]
- Goto, S.; Kogure, K.; Abe, K.; Kimata, Y.; Kitahama, K.; Yamashita, E.; Terada, H. Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochim. Biophys. Acta-Biomembr. 2001, 1512, 251–258. [Google Scholar] [CrossRef]
- Woodall, A.A.; Lee, S.W.M.; Weesie, R.J.; Jackson, M.J.; Britton, G. Oxidation of carotenoids by free radicals: Relationship between structure and reactivity. Biochim. Biophys. Acta-Gen. Subj. 1997, 1336, 33–42. [Google Scholar] [CrossRef]
- Ballesteros, D.; Walters, C. Calorimetric properties of water and triacylglycerols in fern spores relating to storage at cryogenic temperatures. Cryobiology 2007, 55, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Walters-Vertucci, C.; Hoekstra, F.A.; Leprince, O. Calorimetric properties of dehydrating pollen (analysis of a desiccation-tolerant and an intolerant species). Plant Physiol. 1996, 111, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, C. Principles for preserving germplasm in gene banks. In Ex Situ Plant Conservation: Supporting Species Survival in the Wild; Guerrant, E.O., Haven, K., Maunder, M., Raven, T.H., Eds.; Island Press: Washington, DC, USA, 2004. [Google Scholar]
- Buitink, J.; Hemminga, M.A.; Hoekstra, F.A. Characterization of molecular mobility in seed tissues: An electron paramagnetic resonance spin probe study. Biophys. J. 1999, 76, 3315–3322. [Google Scholar] [CrossRef] [Green Version]
- Walters, C.; Wheeler, L.; Stanwood, P.C. Longevity of cryogenically stored seeds. Cryobiology 2004, 48, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Small, D. The physical chemistry of lipids from alkanes to phospholipids. Handb. Lipid Res. 1986, 4, 1–672. [Google Scholar]
- Crane, J.; Miller, A.L.; Van Roekel, J.W.; Walters, C. Triacylglycerols determine the unusual storage physiology of Cuphea seed. Planta 2003, 217, 699–708. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Z.; Hu, Q.; Sommerfeld, M.; Lu, Y.; Han, D. Cellular capacities for high-light acclimation and changing lipid profiles across life cycle stages of the green alga Haematococcus pluvialis. PLoS ONE 2014, 9, e106679. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, D.; Pritchard, H.W.; Walters, C. Dry architecture: Towards the understanding of the variation of longevity in desiccation-tolerant germplasm. Seed Sci. Res. 2020, 30, 142–155. [Google Scholar] [CrossRef]
- Nagel, M.; Kranner, I.; Neumann, K.; Rolletschek, H.; Seal, C.E.; Colville, L.; Fernández-Marín, B.; Börner, A. Genome-wide association mapping and biochemical markers reveal that seed ageing and longevity are intricately affected by genetic background and developmental and environmental conditions in barley. Plant Cell Environ. 2015, 38, 1011–1022. [Google Scholar] [CrossRef]
- Pospíšil, P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. Biochim. Biophys. Acta 2012, 1817, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, T.; Endo, T.; Hashimoto, T.; Yamada, Y.; Asada, K.; Yokota, A. Directed disruption of the tobacco ndhb gene impairs cyclic electron flow around photosystem i. Proc. Natl. Acad. Sci. USA 1998, 95, 9705–9709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamori, W.; Shikanai, T.; Makino, A. Photosystem I cyclic electron flow via chloroplast NADH dehydrogenase-like complex performs a physiological role for photosynthesis at low light. Sci. Rep. 2015, 5, 13908. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Baur, T.; Stoggl, W.; Krieger-Liszkay, A. Chlamydomonas reinhardtii responding to high light: A role for 2-propenal (acrolein). Physiol. Plant 2017, 161, 75–87. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | RH (%) | O2 (%) | Light | P50 (Days) | P50 (Years) |
|---|---|---|---|---|---|
| 20 | 50 | 21 | Dark | 148 | 0.4 |
| 4 | 50 | 21 | Dark | 383 | 1.1 |
| −20 | 50 | 21 | Dark | 299,885 * | 821.6 * |
| −50 | 50 | 21 | Dark | 3.8 × 1012 * | 10 × 109 * |
| −80 | 50 | 21 | Dark | 1.7 × 1012 * | 4.7 × 109 * |
| 21 | 80 | 90 | Dark | 41.8 | <0.1 |
| 21 | 80 | 1 | Dark | 40.2 | <0.1 |
| 21 | 80 | 90 | Light | 21.4 | <0.1 |
| 21 | 80 | 1 | Light | 37.9 | <0.1 |
| 21 | 92.5 | 90 | Dark | 7.3 | <0.1 |
| 21 | 92.5 | 1 | Dark | 17.2 | <0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roach, T.; Fambri, A.; Ballesteros, D. Humidity and Light Modulate Oxygen-Induced Viability Loss in Dehydrated Haematococcus lacustris Cells. Oxygen 2022, 2, 503-517. https://doi.org/10.3390/oxygen2040033
Roach T, Fambri A, Ballesteros D. Humidity and Light Modulate Oxygen-Induced Viability Loss in Dehydrated Haematococcus lacustris Cells. Oxygen. 2022; 2(4):503-517. https://doi.org/10.3390/oxygen2040033
Chicago/Turabian StyleRoach, Thomas, Alessandro Fambri, and Daniel Ballesteros. 2022. "Humidity and Light Modulate Oxygen-Induced Viability Loss in Dehydrated Haematococcus lacustris Cells" Oxygen 2, no. 4: 503-517. https://doi.org/10.3390/oxygen2040033