
Lost in Translation: Exploring microRNA Biogenesis and Messenger RNA Fate in Anoxia-Tolerant Turtles


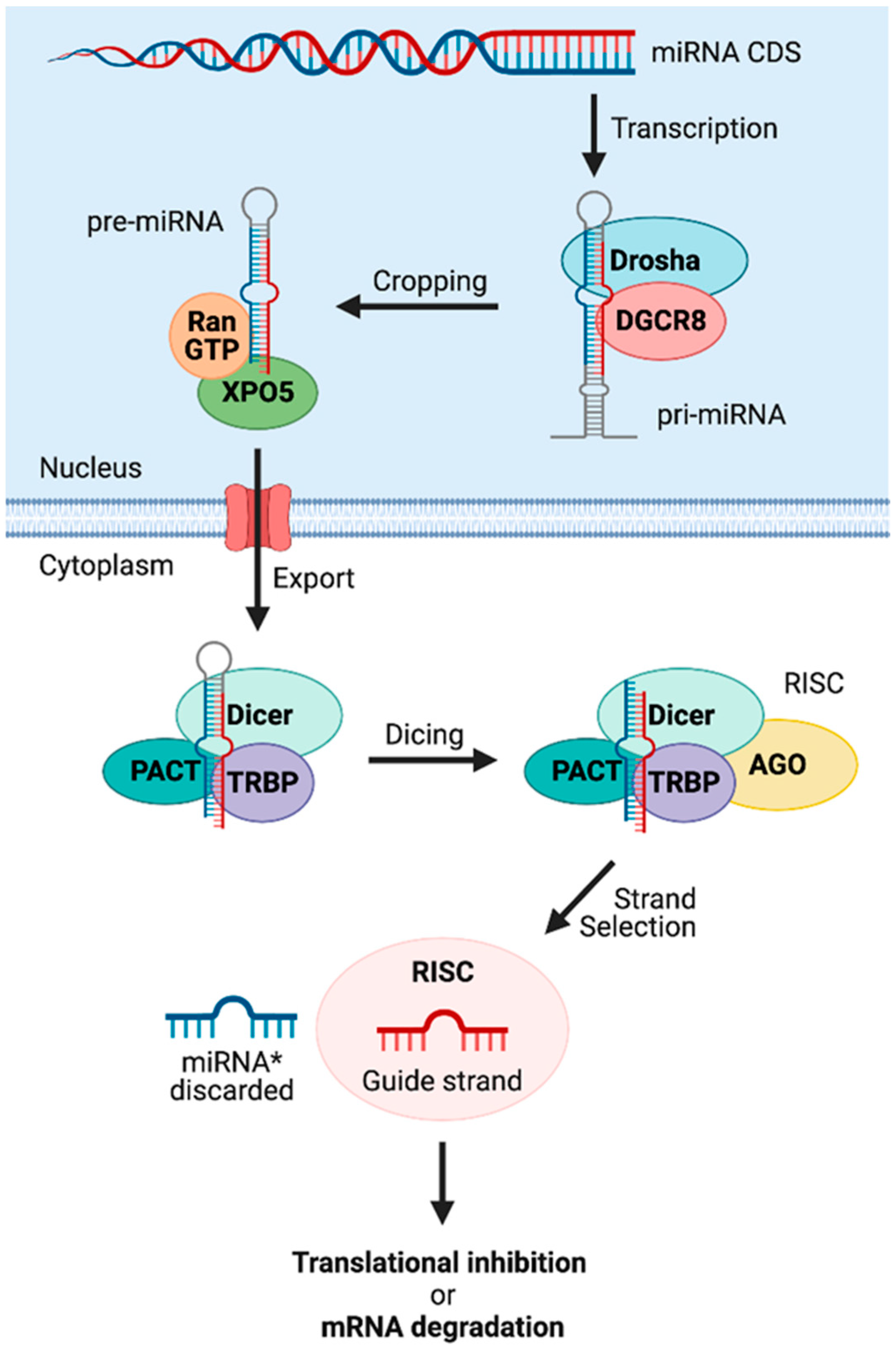{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Total Protein Extraction
2.3. Immunoblotting
2.4. Data Quantification and Statistics
3. Results
3.1. Liver microRNA Biogenesis Protein Expression
3.2. Muscle microRNA Biogenesis Protein Expression
3.3. Liver P-Body and Stress Granule Protein Expression
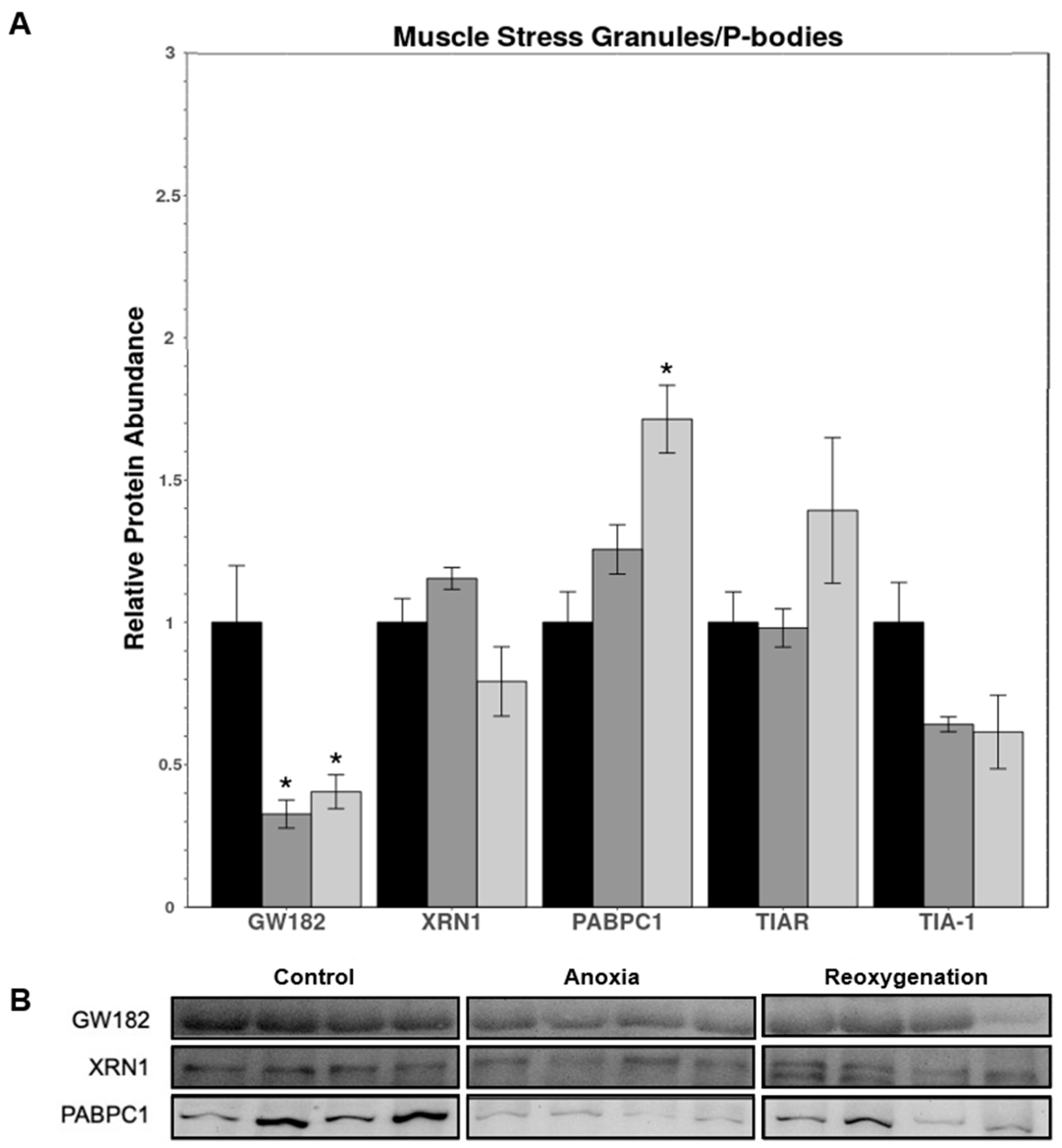
3.4. Muscle P-Body and Stress Granule Protein Expression
4. Discussion
4.1. Regulation of microRNA Biogenesis
4.2. Messenger RNA Fate in P-Bodies and Stress Granules
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, D.C.; Ultsch, G.R. Physiology of hibernation under the ice by turtles and frogs. J. Exp. Zoöl. Part A Ecol. Genet. Physiol. 2010, 313A, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B. Anoxia tolerance in turtles: Metabolic regulation and gene expression. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.C. Metabolic depression and oxygen depletion in the diving turtle. J. Appl. Physiol. 1968, 24, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Ultsch, G.R. Ecology and physiology of hibernation and overwintering among freshwater fishes, turtles, and snakes. Biol. Rev.—Camb. Philos. Soc. 1989, 64, 435–516. [Google Scholar] [CrossRef]
- Breedon, S.A.; Hadj-Moussa, H.; Storey, K.B. Nrf2 activates antioxidant enzymes in the anoxia-tolerant red-eared slider turtle, Trachemys scripta elegans. J. Exp. Zoöl. Part A Ecol. Integr. Physiol. 2021, 335, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Bundgaard, A.; James, A.M.; Joyce, W.; Murphy, M.P.; Fago, A. Suppression of reactive oxygen species generation in heart mitochondria from anoxic turtles: The role of complex I S-nitrosation. J. Exp. Biol. 2018, 221, jeb174391. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.C.; Jackson, D.C. Lactate uptake by skeletal bone in anoxic turtles, Trachemys scripta. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 299–304. [Google Scholar] [CrossRef]
- Kesaraju, S.; Schmidt-Kastner, R.; Prentice, H.M.; Milton, S.L. Modulation of stress proteins and apoptotic regulators in the anoxia tolerant turtle brain. J. Neurochem. 2009, 109, 1413–1426. [Google Scholar] [CrossRef] [Green Version]
- Kesaraju, S.; Nayak, G.; Prentice, H.M.; Milton, S.L. Upregulation of Hsp72 mediates anoxia/reoxygenation neuroprotection in the freshwater turtle via modulation of ROS. Brain Res. 2014, 1582, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Krivoruchko, A.; Storey, K.B. Regulation of the heat shock response under anoxia in the turtle, Trachemys scripta elegans. J. Comp. Physiol. B 2010, 180, 403–414. [Google Scholar] [CrossRef]
- Reiterer, M.; Milton, S.L. Induction of foxo3a protects turtle neurons against oxidative stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 243, 110671. [Google Scholar] [CrossRef] [PubMed]
- Stecyk, J.A.W.; Couturier, C.S.; Fagernes, C.E.; Ellefsen, S.; Nilsson, G.E. Quantification of heat shock protein mRNA expression in warm and cold anoxic turtles (Trachemys scripta) using an external RNA control for normalization. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 7, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Willmore, W.G.; Storey, K. Antioxidant systems and anoxia tolerance in a freshwater turtle Trachemys scripta elegans. Mol. Cell. Biochem. 1997, 170, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W. Metabolic suppression and oxygen availability. Can. J. Zoöl. 1988, 66, 152–158. [Google Scholar] [CrossRef]
- Storey, K.B. Adventures in oxygen metabolism. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.P.; Stecyk, J.A.W. The heart as a working model to explore themes and strategies for anoxic survival in ectothermic vertebrates. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 300–312. [Google Scholar] [CrossRef]
- Biggar, K.K.; Storey, K.B. Evidence for cell cycle suppression and microRNA regulation of cyclin D1 during anoxia exposure in turtles. Cell Cycle 2012, 11, 1705–1713. [Google Scholar] [CrossRef] [Green Version]
- Rider, M.H.; Hussain, N.; Dilworth, S.M.; Storey, K.B. Phosphorylation of translation factors in response to anoxia in turtles, Trachemys scripta elegans: Role of the AMP-activated protein kinase and target of rapamycin signalling pathways. Mol. Cell. Biochem. 2009, 332, 207–213. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Metabolic rate depression in animals: Transcriptional and translational controls. Biol. Rev. Camb. Philos. Soc. 2004, 79, 207–233. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Hadj-Moussa, H.; Zhang, J.; Pifferi, F.; Perret, M.; Storey, K.B. Profiling torpor-responsive microRNAs in muscles of the hibernating primate Microcebus murinus. Biochim. Biophys. Acta—Gene Regul. Mech 2020, 1863, 194473. [Google Scholar] [CrossRef]
- Hadj-Moussa, H.; Pamenter, M.E.; Storey, K.B. Hypoxic naked mole–rat brains use microRNA to coordinate hypometabolic fuels and neuroprotective defenses. J. Cell. Physiol. 2021, 236, 5080–5097. [Google Scholar] [CrossRef] [PubMed]
- Hoyeck, M.P.; Hadj-Moussa, H.; Storey, K.B. Estivation-responsive microRNAs in a hypometabolic terrestrial snail. PeerJ 2019, 7, e6515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, B.E.; Lefai, E.; Giroud, S.; Swenson, J.E.; Chazarin, B.; Gauquelin-Koch, G.; Arnemo, J.M.; Evans, A.L.; Bertile, F.; Storey, K.B. MicroRNAs facilitate skeletal muscle maintenance and metabolic suppression in hibernating brown bears. J. Cell. Physiol. 2020, 235, 3984–3993. [Google Scholar] [CrossRef]
- Biggar, K.K.; Storey, K.B. Exploration of low temperature microRNA function in an anoxia tolerant vertebrate ectotherm, the red eared slider turtle (Trachemys scripta elegans). J. Therm. Biol. 2017, 68, 139–146. [Google Scholar] [CrossRef]
- Zhang, J.; Biggar, K.K.; Storey, K.B. Regulation of p53 by reversible post-transcriptional and post-translational mechanisms in liver and skeletal muscle of an anoxia tolerant turtle, Trachemys scripta elegans. Gene 2013, 513, 147–155. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Finnegan, E.F.; Pasquinelli, A.E. MicroRNA biogenesis: Regulating the regulators. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 51–68. [Google Scholar] [CrossRef] [Green Version]
- Koscianska, E.; Starega-Roslan, J.; Krzyzosiak, W.J. The Role of Dicer Protein Partners in the Processing of MicroRNA Precursors. PLoS ONE 2011, 6, e28548. [Google Scholar] [CrossRef] [Green Version]
- Leung, A.K.L.; Sharp, P.A. Function and Localization of MicroRNAs in Mammalian Cells. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Kedersha, N. Stress granules: The Tao of RNA triage. Trends Biochem. Sci. 2008, 33, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.; Stoecklin, G.; Ayodele, M.; Yacono, P.; Lykke-Andersen, J.; Fritzler, M.J.; Scheuner, D.; Kaufman, R.J.; Golan, D.E.; Anderson, P. Stress granules and processing bodies are dynamically linked sites of mRNP remodeling. J. Cell Biol. 2005, 169, 871–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standart, N.; Weil, D. P-Bodies: Cytosolic Droplets for Coordinated mRNA Storage. Trends Genet. 2018, 34, 612–626. [Google Scholar] [CrossRef]
- Li, Y.R.; King, O.D.; Shorter, J.; Gitler, A.D. Stress granules as crucibles of ALS pathogenesis. J. Cell Biol. 2013, 201, 361–372. [Google Scholar] [CrossRef]
- Buchan, J.R. MRNP Granules Assembly, Function, and Connections with Disease. RNA Biol. 2014, 11, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Buchan, J.R.; Parker, R. Eukaryotic Stress Granules: The Ins and Outs of Translation. Mol. Cell 2009, 36, 932–941. [Google Scholar] [CrossRef] [Green Version]
- Parker, R.; Sheth, U. P Bodies and the Control of mRNA Translation and Degradation. Mol. Cell 2007, 25, 635–646. [Google Scholar] [CrossRef]
- Parker, R. RNA Degradation in Saccharomyces cerevisae. Genetics 2012, 191, 671–702. [Google Scholar] [CrossRef] [Green Version]
- Kedersha, N.; Anderson, P. Mammalian Stress Granules and Processing Bodies. Methods Enzymol. 2007, 431, 61–81. [Google Scholar] [CrossRef]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational Regulation of Gene Expression during Conditions of Cell Stress. Mol. Cell 2010, 40, 228–237. [Google Scholar] [CrossRef]
- Gilks, N.; Kedersha, N.; Ayodele, M.; Shen, L.; Stoecklin, G.; Dember, L.M.; Anderson, P. Stress Granule Assembly Is Mediated by Prion-like Aggregation of TIA-1. Mol. Biol. Cell 2004, 15, 5383–5398. [Google Scholar] [CrossRef] [Green Version]
- Kedersha, N.L.; Gupta, M.; Li, W.; Miller, I.; Anderson, P. RNA-Binding Proteins Tia-1 and Tiar Link the Phosphorylation of Eif-2α to the Assembly of Mammalian Stress Granules. J. Cell Biol. 1999, 147, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Tourrière, H.; Chebli, K.; Zekri, L.; Courselaud, B.; Blanchard, J.M.; Bertrand, E.; Tazi, J. The RasGAP-associated endoribonuclease G3BP assembles stress granules. J. Cell Biol. 2003, 160, 823–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakymiw, A.; Lian, S.; Eystathioy, T.; Li, S.; Satoh, M.; Hamel, J.C.; Fritzler, M.J.; Chan, E.K.L. Disruption of GW bodies impairs mammalian RNA interference. Nat. Cell Biol. 2005, 7, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Eystathioy, T.; Jakymiw, A.; Chan, E.K.L.; Séraphin, B.; Cougot, N.; Fritzler, M.J. The GW182 protein colocalizes with mRNA degradation associated proteins hDcp1 and hLSm4 in cytoplasmic GW bodies: FIGURE 1. RNA 2003, 9, 1171–1173. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Parker, R. The Discovery and Analysis of P Bodies. Adv. Exp. Med. Biol. 2013, 768, 23–43. [Google Scholar] [CrossRef]
- Christie, M.; Boland, A.; Huntzinger, E.; Weichenrieder, O.; Izaurralde, E. Structure of the PAN3 Pseudokinase Reveals the Basis for Interactions with the PAN2 Deadenylase and the GW182 Proteins. Mol. Cell 2013, 51, 360–373. [Google Scholar] [CrossRef] [Green Version]
- Jonas, S.; Izaurralde, E. Towards a Molecular Understanding of MicroRNA-Mediated Gene Silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef]
- Hoyle, N.; Castelli, L.M.; Campbell, S.G.; Holmes, L.E.; Ashe, M.P. Stress-dependent relocalization of translationally primed mRNPs to cytoplasmic granules that are kinetically and spatially distinct from P-bodies. J. Cell Biol. 2007, 179, 65–74. [Google Scholar] [CrossRef]
- Dang, Y.; Kedersha, N.; Low, W.-K.; Romo, D.; Gorospe, M.; Kaufman, R.; Anderson, P.; Liu, J.O. Eukaryotic Initiation Factor 2α-independent Pathway of Stress Granule Induction by the Natural Product Pateamine A. J. Biol. Chem. 2006, 281, 32870–32878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazroui, R.; Sukarieh, R.; Bordeleau, M.-E.; Kaufman, R.J.; Northcote, P.; Tanaka, J.; Gallouzi, I.; Pelletier, J. Inhibition of Ribosome Recruitment Induces Stress Granule Formation Independently of Eukaryotic Initiation Factor 2α Phosphorylation. Mol. Biol. Cell 2006, 17, 4212–4219. [Google Scholar] [CrossRef] [PubMed]
- Mollet, S.; Cougot, N.; Wilczynska, A.; Dautry, F.; Kress, M.; Bertrand, E.; Weil, D. Translationally Repressed mRNA Transiently Cycles through Stress Granules during Stress. Mol. Biol. Cell 2008, 19, 4469–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadj-Moussa, H.; Storey, K.B. Micromanaging freeze tolerance: The biogenesis and regulation of neuroprotective microRNAs in frozen brains. Cell. Mol. Life Sci. 2018, 75, 3635–3647. [Google Scholar] [CrossRef] [PubMed]
- Keiver, K.M.; Hochachka, P.W. Catecholamine stimulation of hepatic glycogenolysis during anoxia in the turtle Chrysemys picta. Am. J. Physiol. 1991, 261, 1341–1345. [Google Scholar] [CrossRef] [PubMed]
- Mehrani, H.; Storey, K.B. Enzymatic control of glycogenolysis during anoxic submergence in the freshwater turtle Trachemys scripta. Int. J. Biochem. Cell Biol. 1995, 27, 821–830. [Google Scholar] [CrossRef]
- Warren, D.E.; Reese, S.A.; Jackson, D.C. Tissue Glycogen and Extracellular Buffering Limit the Survival of Red-Eared Slider Turtles during Anoxic Submergence at 3 °C. Physiol. Biochem. Zoöl. 2006, 79, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.G.; Wang, T.; Madsen, P.T. Hypoxic turtles keep their cool. Temperature 2015, 2, 40–41. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, G.E.; Lutz, P.L. Anoxia Tolerant Brains. J. Cereb. Blood Flow Metab. 2004, 24, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Wijenayake, S.; Hawkins, L.; Storey, K.B. Dynamic regulation of six histone H3 lysine (K) methyltransferases in response to prolonged anoxia exposure in a freshwater turtle. Gene 2018, 649, 50–57. [Google Scholar] [CrossRef]
- Zhang, J.; Storey, K.B. RBioplot: An easy-to-use R pipeline for automated statistical analysis and data visualization in molecular biology and biochemistry. PeerJ 2016, 4, e2436. [Google Scholar] [CrossRef] [PubMed]
- Biggar, K.K.; Zhang, J.; Storey, K.B. Navigating oxygen deprivation: Liver transcriptomic responses of the red eared slider turtle to environmental anoxia. PeerJ 2019, 7, e8144. [Google Scholar] [CrossRef] [PubMed]
- Couturier, C.S.; Stecyk, J.A.W.; Ellefsen, S.; Sandvik, G.K.; Milton, S.L.; Prentice, H.M.; Nilsson, G.E. The expression of genes involved in excitatory and inhibitory neurotransmission in turtle (Trachemys scripta) brain during anoxic submergence at 21 °C and 5 °C reveals the importance of cold as a preparatory cue for anoxia survival. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Douglas, D.N.; Giband, M.; Altosaar, I.; Storey, K.B. Anoxia induces changes in translatable mRNA populations in turtle organs: A possible adaptive strategy for anaerobiosis. J. Comp. Physiol. B 1994, 164, 405–414. [Google Scholar] [CrossRef]
- Keenan, S.W.; Hill, C.A.; Kandoth, C.; Buck, L.T.; Warren, D.E. Transcriptomic Responses of the Heart and Brain to Anoxia in the Western Painted Turtle. PLoS ONE 2015, 10, e0131669. [Google Scholar] [CrossRef]
- Melleby, A.O.; Sandvik, G.K.; Couturier, C.S.; Nilsson, G.E.; Stecyk, J.A.W. H2S-producing enzymes in anoxia-tolerant vertebrates: Effects of cold acclimation, anoxia exposure and reoxygenation on gene and protein expression. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 243–244, 110430. [Google Scholar] [CrossRef]
- Sparks, K.; Couturier, C.S.; Buskirk, J.; Flores, A.; Hoeferle, A.; Hoffman, J.; Stecyk, J.A.W. Gene expression of hypoxia-inducible factor (HIF), HIF regulators, and putative HIF targets in ventricle and telencephalon of Trachemys scripta acclimated to 21 °C or 5 °C and exposed to normoxia, anoxia or reoxygenation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 267, 111167. [Google Scholar] [CrossRef]
- Leung, A.; Sharp, P.A. MicroRNA Functions in Stress Responses. Mol. Cell 2010, 40, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Zlotorynski, E. Insights into the kinetics of microRNA biogenesis and turnover. Nat. Rev. Mol. Cell Biol. 2019, 20, 511. [Google Scholar] [CrossRef]
- Ma, E.; MacRae, I.J.; Kirsch, J.F.; Doudna, J.A. Autoinhibition of Human Dicer by Its Internal Helicase Domain. J. Mol. Biol. 2008, 380, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Bortolamiol-Becet, D.; Hu, F.; Jee, D.; Wen, J.; Okamura, K.; Lin, C.-J.; Ameres, S.L.; Lai, E.C. Selective Suppression of the Splicing-Mediated MicroRNA Pathway by the Terminal Uridyltransferase Tailor. Mol. Cell 2015, 59, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichholf, B.; Herzog, V.A.; Fasching, N.; Manzenreither, R.A.; Sowemimo, I.; Ameres, S.L. Time-Resolved Small RNA Sequencing Unravels the Molecular Principles of MicroRNA Homeostasis. Mol. Cell 2019, 75, 756–768.e7. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.R.; Ignatova, V.; Burkard, T.; Hung, J.-H.; Manzenreither, R.A.; Sowemimo, I.; Herzog, V.A.; Reichholf, B.; Fariña-Lopez, S.; Ameres, S.L. Uridylation of RNA Hairpins by Tailor Confines the Emergence of MicroRNAs in Drosophila. Mol. Cell 2015, 59, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, Z.; O’Loughlin, E.; Lee, T.; Houel, S.; O’Carroll, D.; Tarakhovsky, A.; Ahn, N.G.; Yi, R. Quantitative functions of Argonaute proteins in mammalian development. Genes Dev. 2012, 26, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: MicroRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef]
- Horman, S.R.; Janas, M.M.; Litterst, C.; Wang, B.; MacRae, I.J.; Sever, M.J.; Morrissey, D.V.; Graves, P.; Luo, B.; Umesalma, S.; et al. Akt-Mediated Phosphorylation of Argonaute 2 Downregulates Cleavage and Upregulates Translational Repression of MicroRNA Targets. Mol. Cell 2013, 50, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Xia, W.; Khotskaya, Y.B.; Huo, L.; Nakanishi, K.; Lim, S.-O.; Du, Y.; Wang, Y.; Chang, W.-C.; Chen, C.-H.; et al. EGFR modulates microRNA maturation in response to hypoxia through phosphorylation of AGO2. Nature 2013, 497, 383–387. [Google Scholar] [CrossRef]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Lainé, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a regulator of cellular PKR and HIV-1 virus expression, interacts with Dicer and functions in RNA silencing. EMBO Rep. 2005, 6, 961–967. [Google Scholar] [CrossRef]
- Lee, Y.; Hur, I.; Park, S.-Y.; Kim, Y.-K.K.; Suh, M.R.; Kim, V.N.; Mi, R.S.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J. 2006, 25, 522–532. [Google Scholar] [CrossRef]
- Liu, J.; Rivas, F.V.; Wohlschlegel, J.; Yates, J.R.; Parker, R.; Hannon, G.J. A role for the P-body component GW182 in microRNA function. Nat. Cell Biol. 2005, 7, 1161–1166. [Google Scholar] [CrossRef]
- Yao, B.; Li, S.; Chan, E.K.L. Function of GW182 and GW Bodies in siRNA and miRNA Pathways. Adv. Exp. Med. Biol. 2013, 768, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Tessier, S.N.; Audas, T.E.; Wu, C.-W.; Lee, S.; Storey, K.B. The involvement of mRNA processing factors TIA-1, TIAR, and PABP-1 during mammalian hibernation. Cell Stress Chaperon. 2014, 19, 813–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begley, V.; Corzo, D.; Jordán-Pla, A.; Cuevas-Bermúdez, A.; De Miguel-Jiménez, L.; Pérez-Aguado, D.; Machuca-Ostos, M.; Navarro, F.; Chávez, M.J.; Pérez-Ortín, J.E.; et al. The mRNA degradation factor Xrn1 regulates transcription elongation in parallel to Ccr4. Nucleic Acids Res. 2019, 47, 9524–9541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasco-Moreno, B.; De Campos-Mata, L.; Böttcher, R.; Garcia-Martinez, J.; Jungfleisch, J.; Nedialkova, D.D.; Chattopadhyay, S.; Gas, M.-E.; Oliva, B.; Pérez-Ortín, J.E.; et al. The exonuclease Xrn1 activates transcription and translation of mRNAs encoding membrane proteins. Nat. Commun. 2019, 10, 1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breedon, S.A.; Storey, K.B. Lost in Translation: Exploring microRNA Biogenesis and Messenger RNA Fate in Anoxia-Tolerant Turtles. Oxygen 2022, 2, 227-245. https://doi.org/10.3390/oxygen2020017
Breedon SA, Storey KB. Lost in Translation: Exploring microRNA Biogenesis and Messenger RNA Fate in Anoxia-Tolerant Turtles. Oxygen. 2022; 2(2):227-245. https://doi.org/10.3390/oxygen2020017
Chicago/Turabian StyleBreedon, Sarah A., and Kenneth B. Storey. 2022. "Lost in Translation: Exploring microRNA Biogenesis and Messenger RNA Fate in Anoxia-Tolerant Turtles" Oxygen 2, no. 2: 227-245. https://doi.org/10.3390/oxygen2020017