
Effects of Dietary Sodium Propionate on Growth, Digestive Enzyme Activity, and Expression of Immune System Genes in Juveniles of Tropical Gar (Atractosteus tropicus)
 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Design
2.3. Experimental Diets
2.4. Growth and Feed Indexes
2.5. Biological Samples and Somatic Indexes
2.6. Digestive Enzyme Activity
2.7. RNA Extraction and qPCR

{kind=link}
{kind=link}
| Gen | Primer Sequence (5′-3′) | Size (bp) | Amplification Efficiency (%) | Reference | Temperature (°C) |
|---|---|---|---|---|---|
| 1Ribosomal protein | F: TGTGCTGCCTGGAAGAGAAG R: TTTCGGGGTTGTGGGAGATG | 90 | 99.82 | [27] | 60 |
| 2Occludin | F: TGACGAATACCACAGACTGAAG R: CGATCATAGTCGCTGACCATC | 123 | [5] | 64 | |
| 3Mucin | F: GGCCTCCTCAAGAGCACGGTG R: TCTGCACGCTGGAGCACTCAATG | 100 | 90.94 | [4] | 70 |
| 4Nucleotide-binding oligomerization domain-2 | F: GTAGTGAACAAGGAGGCGGAC R: TGAGCTCATCCAGGCCATCG | 295 | [5] | 68 |
2.8. Statistical Analysis
3. Results
3.1. Growth and Survival Rates
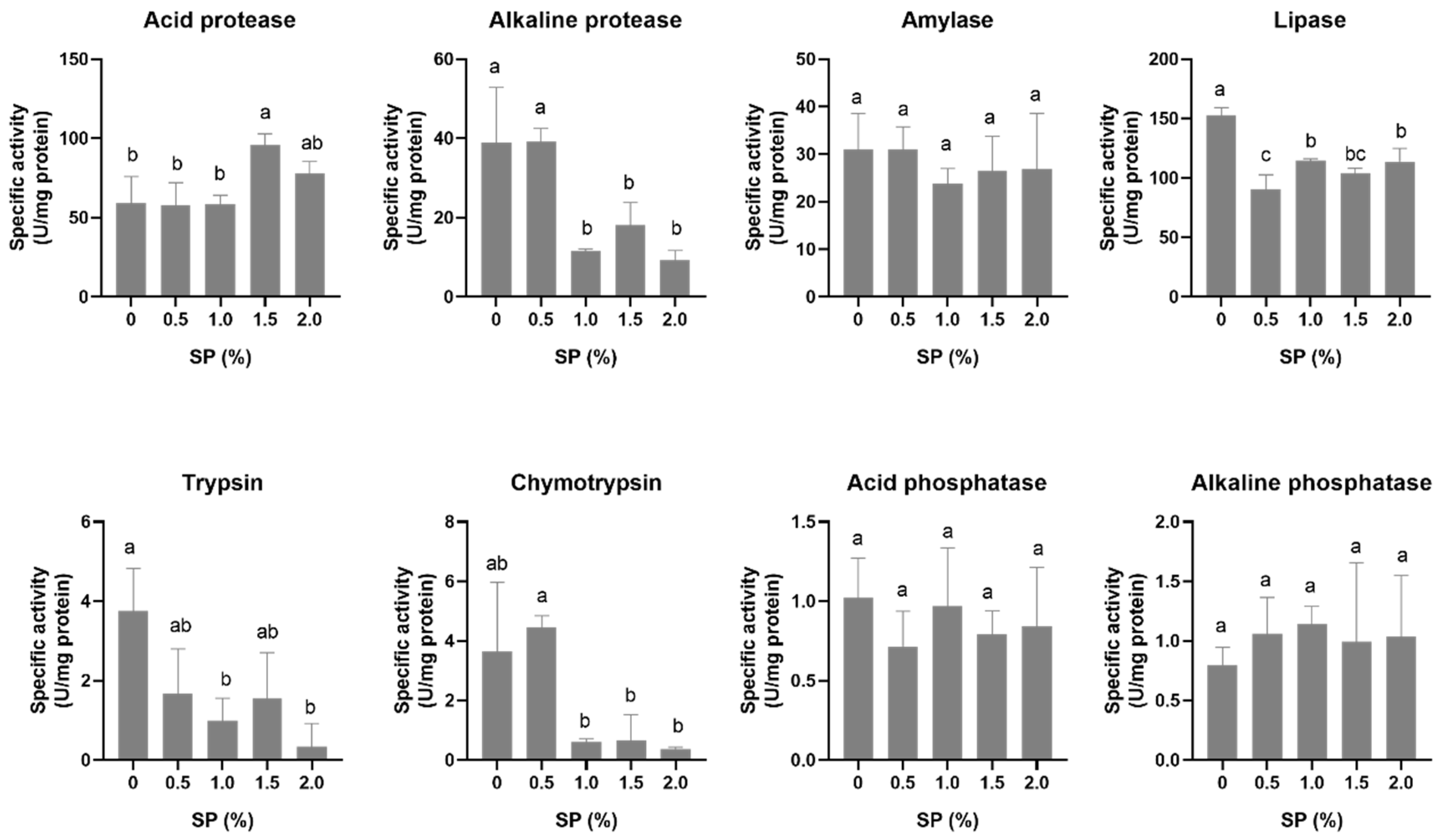
3.2. Digestive Enzyme Activity
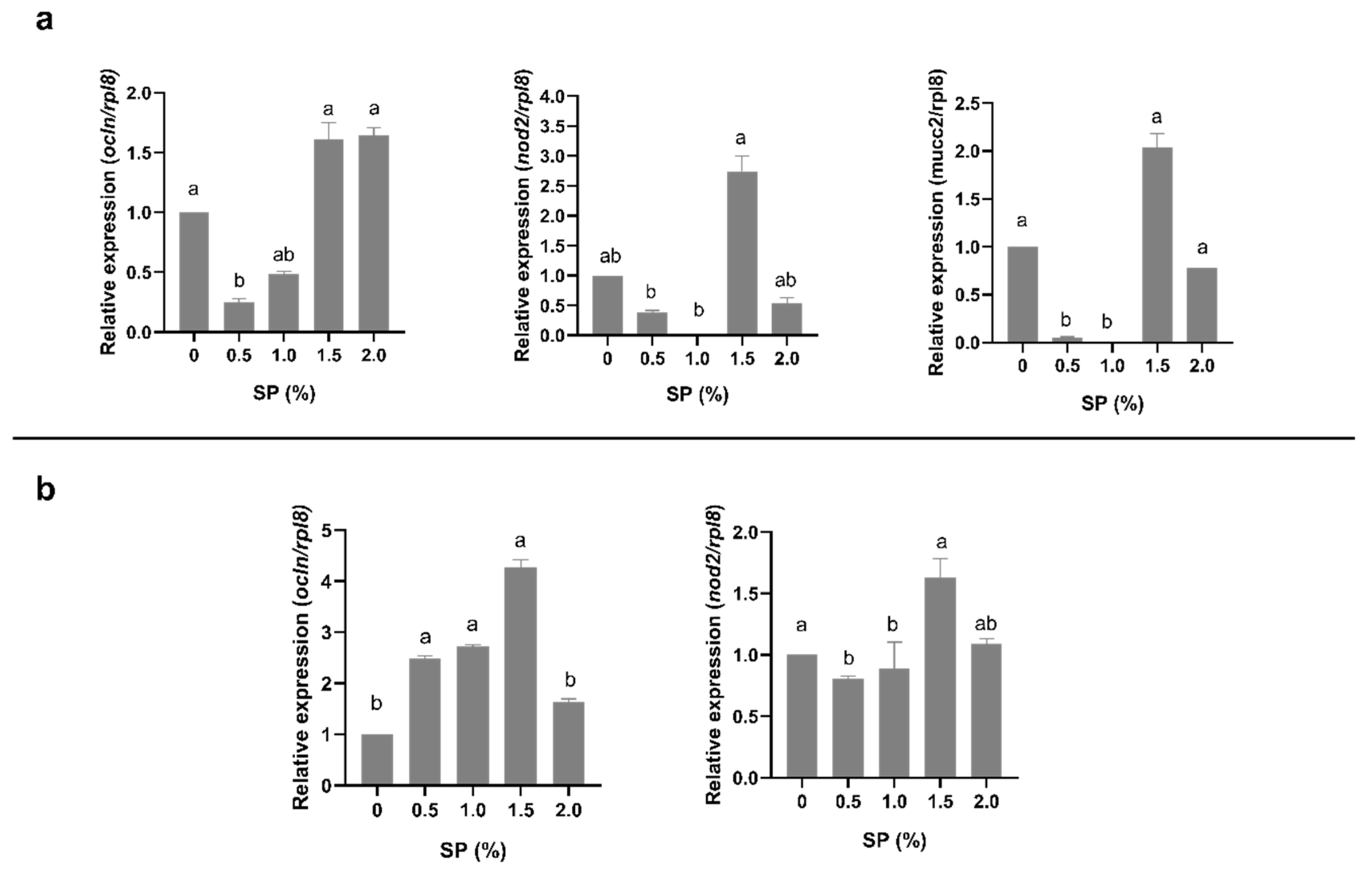
3.3. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Márquez-Couturier, G.; Navarrete-Vázquez, C.; Sánchez-Contreras, W.; Álvarez-González, C.A. Acuicultura Tropical Sustentable, 2nd ed.; Universidad Juárez Autónoma de Tabasco: Villahermosa, Mexico, 2015. [Google Scholar]
- Márquez-Couturier, G.; Vázquez-Navarrete, C. Estado de arte de la biología y cultivo de pejelagarto (Atractosteus tropicus). Agroproductividad 2015, 8, 44–51. [Google Scholar]
- Nájera-Arzola, I.C.; Álvarez-González, C.A.; Frías-Quintana, C.A.; Peña, E.; Martínez-García, R.; Camarillo-Coop, S.; Méndez-Marín, O.; Gisbert, E. Evaluation of Mannan oligosaccharides (MOS) in balanced diets for tropical gar juveniles (Atractosteus tropicus). Hidrobiologica 2018, 28, 239–246. [Google Scholar] [CrossRef]
- Nieves-Rodríguez, K.N.; Álvarez-González, C.A.; Peña-Marín, E.S.; Vega-Villasante, F.; Martínez-García, R.; Camarillo-Coop, S.; Tovar-Ramírez, D.; Guzmán-Villanueva, L.T.; Andree, K.B.; Gisbert, E. Effect of β-Glucans in Diets on Growth, Survival, Digestive Enzyme Activity, and Immune System and Intestinal Barrier Gene Expression for Tropical Gar (Atractosteus tropicus) Juveniles. Fishes 2018, 3, 27. [Google Scholar] [CrossRef]
- Sepúlveda-Quiroz, C.A.; Peña-Marín, E.S.; Pérez-Morales, A.; Martínez-García, R.; Alvarez-Villagomez, C.S.; Maytorena-Verdugo, C.I.; Camarillo-Coop, S.; Vissio, P.G.; Sirkin, D.P.; Tovar-Ramírez, D.; et al. Fructooligosaccharide supplementation in diets for tropical gar (Atractosteus tropicus) juvenile: Effects on morphophysiology and intestinal barrier function. Aquac. Res. 2020, 52, 37–50. [Google Scholar] [CrossRef]
- Cook, S.; Sellin, J.H. Review article: Short chain fatty acids in health and disease. Aliment. Pharmacol. Ther. 1998, 12, 499–507. [Google Scholar] [CrossRef]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lek. Listy 2007, 108, 354–358. [Google Scholar]
- Sotoudeh, E.; Sangari, M.; Bagheri, D.; Morammazi, S.; Mozanzadeh, M.T. Dietary organic acid salts mitigate plant protein induced inflammatory response and improve humoral immunity, antioxidative status and digestive enzyme activities in yellowfin seabream, Acanthopagrus latus. Aquac. Nutr. 2020, 26, 1669–1680. [Google Scholar] [CrossRef]
- da Silva, B.C.; Vieira, F.D.N.; Mouriño, J.L.P.; Bolivar, N.; Seiffert, W.Q. Butyrate and propionate improve the growth performance of Litopenaeus vannamei. Aquac. Res. 2014, 47, 612–623. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Safari, R.; Dadar, M. Dietary sodium propionate affects mucosal immune parameters, growth and appetite related genes expression: Insights from zebrafish model. Gen. Comp. Endocrinol. 2017, 243, 78–83. [Google Scholar] [CrossRef]
- De la Cruz-Alvarado, F.J.; Álvarez-González, C.A.; Llera-Herrera, R.; Monroig, Ó.; Kabeya, N.; Rodríguez-Morales, S.; Concha-Frias, B.; Guerrero-Zárate, R.; Jiménez-Martínez, L.D.; Peña-Marín, E.S. Expression of long-chain polyunsaturated fatty acids biosynthesis genes during the early life-cycle stages of the tropical gar Atractosteus tropicus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110628. [Google Scholar] [CrossRef]
- Frías-Quintana, C.A.; Álvarez-González, C.A.; Tovar-Ramírez, D.; Martínez-García, R.; Camarillo-Coop, S.; Peña, E.; Galaviz, M.A. Use of Potato Starch in Diets of Tropical Gar (Atractosteus tropicus, Gill 1863) Larvae. Fishes 2017, 2, 3. [Google Scholar] [CrossRef]
- Alvarez-González, C.; Civera-Cerecedo, R.; Ortiz-Galindo, J.; Dumas, S.; Moreno-Legorreta, M.; Alamo, T.G.-D. Effect of dietary protein level on growth and body composition of juvenile spotted sand bass, Paralabrax maculatofasciatus, fed practical diets. Aquaculture 2001, 194, 151–159. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemist, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Available online: https://cdnsciencepub.com/doi/abs/10.1139/o59-099 (accessed on 21 September 2022).
- Brett, J.R.; Groves, T.D.D. “Physiological energetics” in Bioenergetics and growth. Fish. Physiol. 1979, 8, 1–786. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999; Especificaciones Técnicas para la Producción, Cuidado y uso de los Animales de Laboratorio. SAGARPA: Mexico City, Mexico, 2001; pp. 1–58.
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Walter, H.E. Proteinases: Methods with hemoglobin, casein and azocoll as substrates. In Methods of Enzymatic Analysis; Bergmeyer, H.J., Ed.; Verlag Chemie: Weinheim, Germany, 1984; pp. 270–277. [Google Scholar]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys. 1961, 95, 271–278. [Google Scholar] [CrossRef]
- DelMar, E.; Largman, C.; Brodrick, J.; Geokas, M. A sensitive new substrate for chymotrypsin. Anal. Biochem. 1979, 99, 316–320. [Google Scholar] [CrossRef]
- Robyt, J.F.; Whelan, W. Amylases. In Starch and Its Derivates; Radley, J.A., Ed.; Chapman and Hall: London, UK, 1968. [Google Scholar]
- Versaw, W.K.; Cuppett, S.L.; Winters, D.D.; Williams, L.E. An Improved Colorimetric Assay for Bacterial Lipase in Nonfat Dry Milk. J. Food Sci. 1989, 54, 1557–1558. [Google Scholar] [CrossRef]
- Bergmeyer, H.U. Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Aranda-Morales, S.A.; Peña-Marin, E.S.; Jiménez-Martínez, L.D.; Martínez-Burguete, T.; Martínez-Bautista, G.; Alavrez-Villagómez, C.S.; Rosa García, S.R.; Camarillo-Coop, S.; Martínez-García, R.; Guzmán-Villanueva, L.T.; et al. Expression of ion transport proteins and routine metabolism in juveniles of tropical gar (Atractosteus tropicus) exposed to ammonia. Comp. Biochem. Physiol. Part C 2021, 250, 109166. [Google Scholar] [CrossRef]
- Wassef, E.A.; Saleh, N.E.; Abdel-Meguid, N.E.; Barakat, K.M.; Abdel-Mohsen, H.H.; El-Bermawy, N.M. Sodium propionate as a dietary acidifier for European seabass (Dicentrarchus labrax) fry: Immune competence, gut microbiome, and intestinal histology benefits. Aquac. Int. 2019, 28, 95–111. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Zoheiri, F.; Caipang, C.M. Dietary sodium propionate improved performance, mucosal and humoral immune responses in Caspian white fish (Rutilus frisii kutum) fry. Fish Shellfish. Immunol. 2016, 55, 523–528. [Google Scholar] [CrossRef] [PubMed]
- da Silva, B.C.; Vieira, F.D.N.; Mouriño, J.L.P.; Ferreira, G.S.; Seiffert, W.Q. Salts of organic acids selection by multiple characteristics for marine shrimp nutrition. Aquaculture 2013, 384–387, 104–110. [Google Scholar] [CrossRef]
- Gatlin, D.M., III. Nutrition and Fish Health. In Fish Nutrition, 3rd ed.; Halver, J., Hardy, R., Eds.; Academic Press: Cambridge, MA, USA, 2002; pp. 672–699. [Google Scholar]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Daeman, N.H.; Chong, C.M.; Karami, A.; Kumar, V.; Hoseinifar, S.H.; Romano, N. Comparing the effects of different dietary organic acids on the growth, intestinal short-chain fatty acids, and liver histopathology of red hybrid tilapia (Oreochromis sp.) and potential use of these as preservatives. Fish Physiol. Biochem. 2017, 43, 1195–1207. [Google Scholar] [CrossRef]
- Maytorena-Verdugo, C.I.; Peña-Marín, E.S.; Alvarez-Villagómez, C.S.; Pérez-Jiménez, G.M.; Sepúlveda-Quiroz, C.A.; Alvarez-González, C.A. Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes. Fishes 2022, 7, 127. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Ahmadi, A.; Khalili, M.; Raeisi, M.; Van Doan, H.; Caipang, C.M. The study of antioxidant enzymes and immune-related genes expression in common carp (Cyprinus carpio) fingerlings fed different prebiotics. Aquac. Res. 2017, 48, 5447–5454. [Google Scholar] [CrossRef]
- Lim, P.; Lückstädt, C.; Webster, C.; Kesius, C.D. Organic Acids and Their Salts. In Dietary Nutrients, Additives, and Fish Health; Lee, C.D., Lim, C.-S., Gatlin, C., III, Webster, D.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 305–315. [Google Scholar]
- Guerrero-Zárate, R.; Alvarez-González, C.A.; Olvera-Novoa, M.A.; Perales-García, N.; Frías-Quintana, C.A.; Martínez-García, R.; Contreras-Sánchez, W.M. Partial characterization of digestive proteases in tropical gar Atractosteus tropicus juveniles. Fish Physiol. Biochem. 2013, 40, 1021–1029. [Google Scholar] [CrossRef]
- Xiong, D.M.; Xie, C.X.; Zhang, H.J.; Liu, H.P. Digestive enzymes along digestive tract of a carnivorous fish Glyptosternum maculatum (Sisoridae, Siluriformes). J. Anim. Physiol. Anim. Nutr. 2011, 95, 56–64. [Google Scholar] [CrossRef]
- Dietary Nutrients, Additives, and Fish Health; Lee, C.-S.; Lim, C.; Gatlin, D.M.; Webster, C.D. (Eds.) John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; p. 355. [Google Scholar]
- Safari, R.; Hoseinifar, S.H.; Kavandi, M. Modulation of antioxidant defense and immune response in zebra fish (Danio rerio) using dietary sodium propionate. Fish Physiol. Biochem. 2016, 42, 1733–1739. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. The role of MUC2 mucin in intestinal homeostasis and the impact of dietary components on MUC2 expression. Int. J. Biol. Macromol. 2020, 164, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; Van Der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; Van Seuningen, I.; Renes, I.B. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: Implications for epithelial protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, G.M.; Peña-Marín, E.S.; Maytorena-Verdugo, C.I.; Sepúlveda-Quiroz, C.A.; Jiménez-Martínez, L.D.; De la Rosa-García, S.; Asencio-Alcudia, G.G.; Martínez, R.; Tovar-Ramírez, D.; Galaviz, M.A.; et al. Incorporation of Fructooligosaccharides in Diets Influence Growth Performance, Digestive Enzyme Activity, and Expression of Intestinal Barrier Function Genes in Tropical Gar (Atractosteus tropicus) Larvae. Fishes 2022, 7, 137. [Google Scholar] [CrossRef]
- Treyer, A.; Müsch, A. Hepatocyte polarity. Compr. Physiol. 2013, 3, 243–287. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Z.; Dai, J.; Yang, P.; Xu, W.; Ai, Q.; Zhang, W.; Zhang, Y.; Zhang, Y.; Mai, K. Sodium butyrate supplementation in high-soybean meal diets for turbot (Scophthalmus maximus L.): Effects on inflammatory status, mucosal barriers and microbiota in the intestine. Fish Shellfish. Immunol. 2019, 88, 65–75. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, J.; Wang, W.; Shao, R.; Liang, S.; Xu, W.; Li, M.; Ai, Q.; Mai, K.; Wan, M. The Effects of Sodium Propionate Supplementation in the Diet with High Soybean Meal on Growth Performance, Intestinal Health, and Immune Resistance to Bacterial Infection in Turbot (Scophthalmus maximus L.). Aquac. Nutr. 2022, 2022, 8952755. [Google Scholar] [CrossRef]
- Li, Y.; Jin, L.; Xia, P.; Sui, W.; Huang, A.; Bu, G.; Meng, F.; Kong, F.; Cao, X.; Han, X.; et al. Identification and functional analysis of NOD2 and its two splicing variants associated with a novel pattern of signal regulation in teleost fishes. Dev. Comp. Immunol. 2021, 120, 104049. [Google Scholar] [CrossRef]
- Li, J.; Gao, Y.; Xu, T. Comparative genomic and evolution of vertebrate NOD1 and NOD2 genes and their immune response in miiuy croaker. Fish Shellfish. Immunol. 2015, 46, 387–397. [Google Scholar] [CrossRef]


| Sodium Propionate (%) | |||||
|---|---|---|---|---|---|
| Ingredients (g/kg) | 0 | 0.5 | 1.0 | 1.5 | 2.0 |
| Pork meal 1 | 250 | 250 | 250 | 250 | 250 |
| Poultry meal 1 | 218.4 | 218.4 | 218.4 | 218.4 | 218.4 |
| Soybean meal 2 | 161.1 | 161.1 | 161.1 | 161.1 | 161.1 |
| Corn starch 3 | 150 | 150 | 150 | 150 | 150 |
| Fish meal 1 | 100 | 100 | 100 | 100 | 100 |
| Wheat meal 2 | 55.5 | 55.5 | 55.5 | 40.5 | 35.5 |
| Sodium propionate 4 | 0 | 5 | 10 | 15 | 20 |
| Soybean lecithin 5 | 30 | 30 | 30 | 30 | 30 |
| Grenetin 6 | 20 | 20 | 20 | 20 | 20 |
| Vitamins and minerals premix 7 | 10 | 10 | 10 | 10 | 10 |
| Vitamin C 8 | 5 | 5 | 5 | 5 | 5 |
| Proximate composition (g/100 g dry matter) | |||||
| Protein | 44.32 | 44.81 | 43.99 | 44.83 | 44.74 |
| Lipids | 15.32 | 15.74 | 15.43 | 15.53 | 15.31 |
| Ash | 12.77 | 12.74 | 13.23 | 13.71 | 13.53 |
| NFE 9 | 28.49 | 28.43 | 28.43 | 28.48 | 28.46 |
| Treatments (SP%) | |||||
|---|---|---|---|---|---|
| Growth Parameters | 0 | 0.5 | 1 | 1.5 | 2 |
| Final weight (g) | 24.19 ± 1.2 | 25.79 ± 4.5 | 22.17 ± 2.5 | 23.32 ± 1.9 | 21.38 ± 1.2 |
| Final length (cm) | 17.60 ± 0.1 | 17.74 ± 0.6 | 17.03 ± 0.5 | 17.60 ± 0.5 | 16.79 ± 0.3 |
| Survival (%) | 80.5 ± 12.7 a | 86.1 ± 4.8 b | 86.11 ± 9.6 b | 91.6 ± 8.3 c | 91.6 ± 8.3 c |
| AWG (g/fish) | 20.53 ± 1.26 | 21.93 ± 4.39 | 18.52 ± 2.81 | 19.81 ± 2 | 17.69 ± 1.02 |
| SGR (%/day) | 3.1 ± 0.14 | 3.1± 0.26 | 3.0 ± 0.34 | 3.05 ± 0.17 | 2.9 ± 0.14 |
| Condition factor (k) | 0.44 ± 0.02 | 0.46 ± 0.03 | 0.45 ± 0.01 | 0.41 ± 0.04 | 0.45 ± 0.01 |
| FCR | 1.61 ± 0.09 | 1.92 ± 0.38 | 1.52 ± 0.23 | 1.46 ± 0.14 | 1.40 ± 0.08 |
| Treatments (SP%) | |||||
|---|---|---|---|---|---|
| Somatic Indexes | 0 | 0.5 | 1 | 1.5 | 2 |
| HSI | 3.26 ± 1.22 | 3.04 ± 0.67 | 2.80 ± 0.47 | 3.47 ± 0.5 | 2.95 ± 0.61 |
| VSI | 9.45 ± 3.65 | 8.73 ± 1.09 | 8.01 ± 1.01 | 10.53 ± 1.77 | 8.10 ± 1.23 |
| RIL | 35.86 ± 4.99 | 35.18 ± 3.67 | 38.68 ± 4.91 | 40.26 ± 4.20 | 30.92 ± 6.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arellano-Carrasco, J.G.; Martínez-García, R.; Asiain-Hoyos, A.; Reta-Mendiola, J.L.; Díaz-Rivera, P.; Frías-Gómez, S.A.; Martínez-Burguete, T.; Asencio-Alcudia, G.G.; Jiménez-Martínez, L.D.; Guerrero-Zarate, R.; et al. Effects of Dietary Sodium Propionate on Growth, Digestive Enzyme Activity, and Expression of Immune System Genes in Juveniles of Tropical Gar (Atractosteus tropicus). Aquac. J. 2023, 3, 227-237. https://doi.org/10.3390/aquacj3040018
Arellano-Carrasco JG, Martínez-García R, Asiain-Hoyos A, Reta-Mendiola JL, Díaz-Rivera P, Frías-Gómez SA, Martínez-Burguete T, Asencio-Alcudia GG, Jiménez-Martínez LD, Guerrero-Zarate R, et al. Effects of Dietary Sodium Propionate on Growth, Digestive Enzyme Activity, and Expression of Immune System Genes in Juveniles of Tropical Gar (Atractosteus tropicus). Aquaculture Journal. 2023; 3(4):227-237. https://doi.org/10.3390/aquacj3040018
Chicago/Turabian StyleArellano-Carrasco, Jesús G., Rafael Martínez-García, Alberto Asiain-Hoyos, Juan L. Reta-Mendiola, Pablo Díaz-Rivera, Susana A. Frías-Gómez, Talhia Martínez-Burguete, Gloria Gertrudys Asencio-Alcudia, Luis Daniel Jiménez-Martínez, Rocio Guerrero-Zarate, and et al. 2023. "Effects of Dietary Sodium Propionate on Growth, Digestive Enzyme Activity, and Expression of Immune System Genes in Juveniles of Tropical Gar (Atractosteus tropicus)" Aquaculture Journal 3, no. 4: 227-237. https://doi.org/10.3390/aquacj3040018