
Effect of Ischemic Preconditioning (IPC) on Recovery of Exercise Performance Following a Bout of Exercise to Volitional Exhaustion

Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Experimental Design
2.3. Incremental Exercise
2.4. Experimental Intervention
2.5. Constant Work-Rate Exercise
2.6. Measurements
2.7. Pulmonary Gas Exchange and Oxygen Uptake Kinetics
2.8. Near-Infrared Spectroscopy (NIRS)
2.9. Oxygen Uptake Kinetics
2.10. NIRS Kinetic Analysis
2.11. Blood Lactate Sampling
2.12. Statistical Analysis
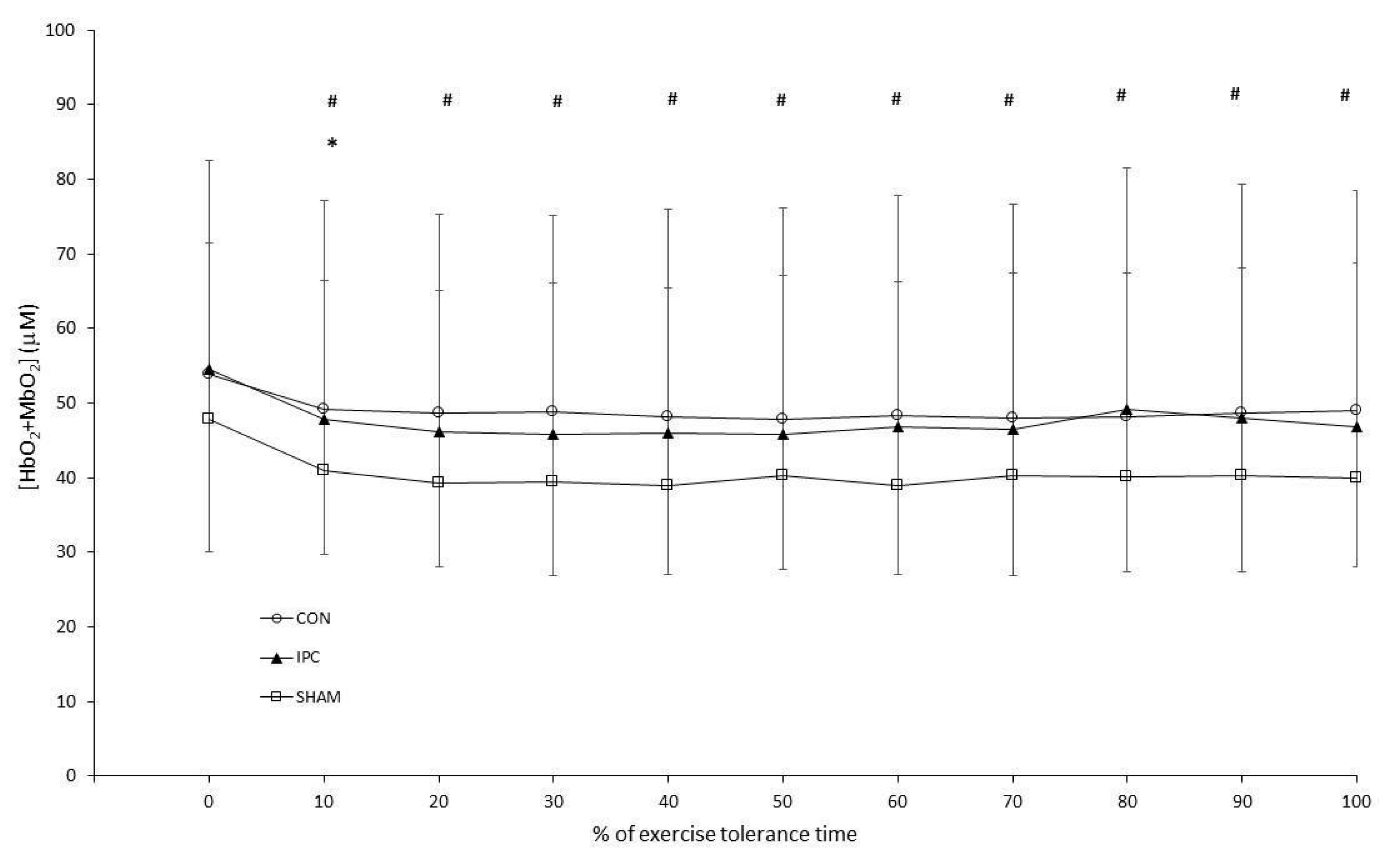
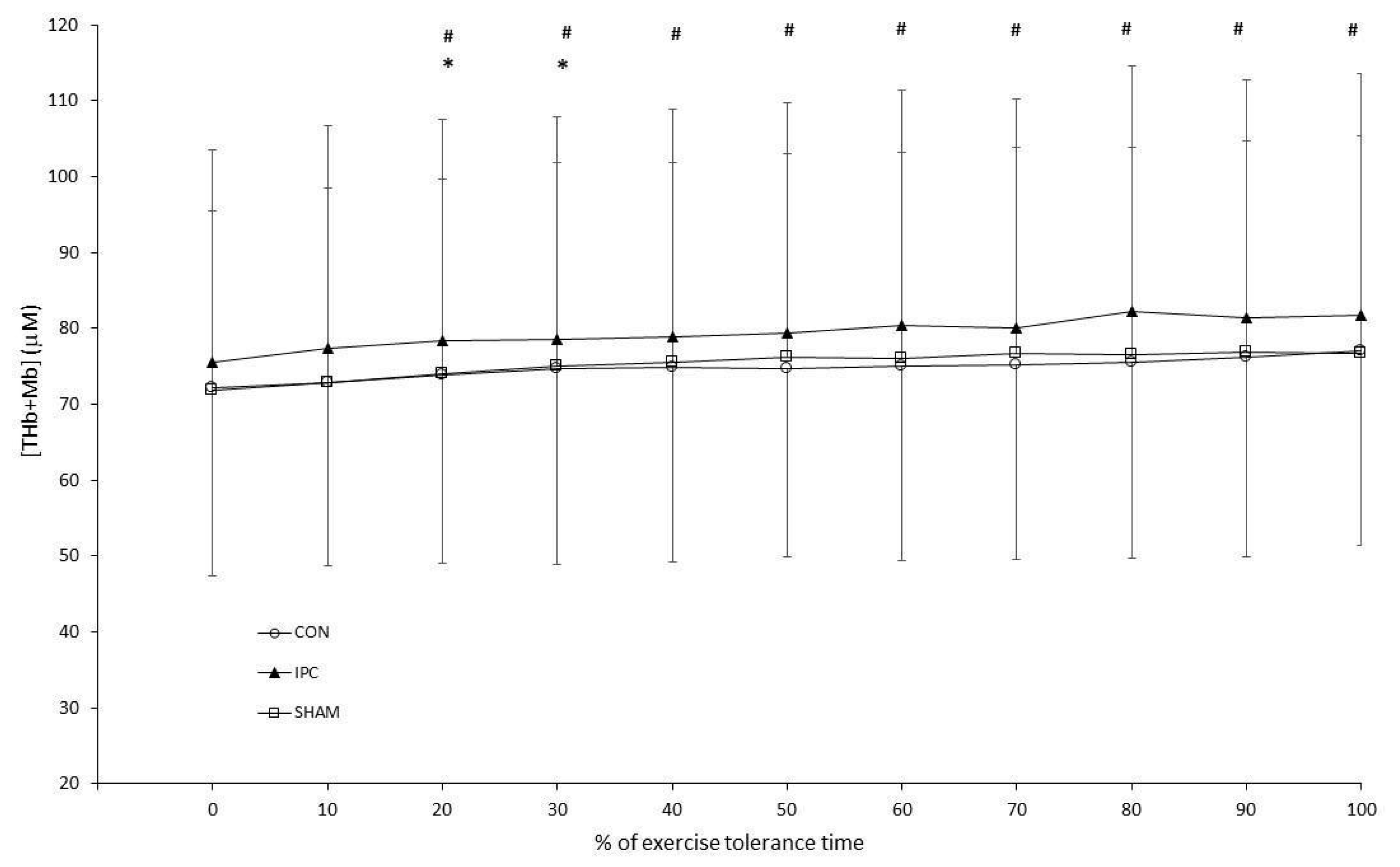
3. Results
3.1. Incremental Exercise
3.2. Constant Work-Rate Exercise
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Groot, P.C.E.; Thijssen, D.H.J.; Sanchez, M.; Ellenkamp, R.; Hopman, M.T.E. Ischemic Preconditioning Improves Maximal Performance in Humans. Eur. J. Appl. Physiol. 2010, 108, 141–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telles, L.G.; Billaut, F.; de Souza Ribeiro, A.; Junqueira, C.G.; Leitão, L.; Barreto, A.C.; Panza, P.; Vianna, J.M.; da Silva Novaes, J. Ischemic Preconditioning with High and Low Pressure Enhances Maximum Strength and Modulates Heart Rate Variability. IJERPH 2022, 19, 7655. [Google Scholar] [CrossRef]
- Da Silva Novaes, J.; da Silva Telles, L.G.; Monteiro, E.R.; da Silva Araujo, G.; Vingren, J.L.; Silva Panza, P.; Reis, V.M.; Laterza, M.C.; Vianna, J.M. Ischemic Preconditioning Improves Resistance Training Session Performance. J. Strength Cond. Res. 2021, 35, 2993–2998. [Google Scholar] [CrossRef]
- Ferreira, T.N.; Sabino-Carvalho, J.L.C.; Lopes, T.R.; Ribeiro, I.C.; Succi, J.E.; Da Silva, A.C.; Silva, B.M. Ischemic Preconditioning and Repeated Sprint Swimming: A Placebo and Nocebo Study. Med. Sci. Sport. Exerc. 2016, 48, 1967–1975. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, J.; Muradov, O.; Li, X.; Lee, J.K.W.; Qiu, J. Effect of Ischemic Preconditioning on Maximum Accumulated Oxygen Deficit in 400-Meter Runners. Eur. J. Sport Sci. 2022, 23, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.; Russell, M.; Cook, C.J.; Kilduff, L.P. Effect of Ischemic Preconditioning on Maximal Swimming Performance. J. Strength Cond. Res. 2021, 35, 221–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoye, A.H.; Mitchinson, C.J.; Townsend, O.R.; Nemmers, C.H.; Serkaian, C.N.; Rider, B.C. Ischemic Preconditioning Does Not Improve Time Trial Performance in Recreational Runners. Int. J. Exerc. Sci. 2020, 13, 1402–1417. [Google Scholar]
- Lalonde, F.; Curnier, D.Y. Can Anaerobic Performance Be Improved by Remote Ischemic Preconditioning? J. Strength Cond. Res. 2015, 29, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Da Mota, G.R.; Marocolo, M. The Effects of Ischemic Preconditioning on Human Exercise Performance: A Counterpoint. Sports Med. 2016, 46, 1575–1576. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Yellon, D.M. Remote Ischaemic Preconditioning: Underlying Mechanisms and Clinical Application. Cardiovasc. Res. 2008, 79, 377–386. [Google Scholar] [CrossRef]
- Goulding, R.P.; Marwood, S. Interaction of Factors Determining Critical Power. Sports Med. 2023, 53, 595–613. [Google Scholar] [CrossRef]
- Jones, A.M.; Burnley, M. Oxygen Uptake Kinetics: An Underappreciated Determinant of Exercise Performance. Int. J. Sport. Physiol. Perform. 2009, 4, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Pang, C.Y.; Neligan, P.; Xu, H.; He, W.; Zhong, A.; Hopper, R.; Forrest, C.R. Role of ATP-Sensitive K+ Channels in Ischemic Preconditioning of Skeletal Muscle against Infarction. Am. J. Physiol. Heart Circ. Physiol. 1997, 273, H44–H51. [Google Scholar] [CrossRef]
- Lee, H.T.; Thompson, C.I. Preconditioning with Ischemia or Adenosine Protects Skeletal Muscle from Ischemic Tissue Reperfusion Injury. J. Surg. Res. 1996, 63, 6. [Google Scholar] [CrossRef] [PubMed]
- Hopper, R.A.; Forrest, C.R.; Xu, H.; Zhong, A.; He, W.; Rutka, J.; Neligan, P.; Pang, C.Y. Role and Mechanism of PKC in Ischemic Preconditioning of Pig Skeletal Muscle against Infarction. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2000, 279, R666–R676. [Google Scholar] [CrossRef]
- Goulding, R.P.; Rossiter, H.B.; Marwood, S.; Ferguson, C. Bioenergetic Mechanisms Linking VO2 Kinetics and Exercise Tolerance. Exerc. Sport Sci. Rev. 2021, 49, 274–283. [Google Scholar] [CrossRef]
- Crisafulli, A.; Tangianu, F.; Tocco, F.; Concu, A.; Mameli, O.; Mulliri, G.; Caria, M.A. Ischemic Preconditioning of the Muscle Improves Maximal Exercise Performance but Not Maximal Oxygen Uptake in Humans. J. Appl. Physiol. 2011, 111, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Kido, K.; Suga, T.; Tanaka, D.; Honjo, T.; Homma, T.; Fujita, S.; Hamaoka, T.; Isaka, T. Ischemic Preconditioning Accelerates Muscle Deoxygenation Dynamics and Enhances Exercise Endurance during the Work-to-Work Test. Physiol. Rep. 2015, 3, e12395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ruiter, C.J.; Jones, D.A.; Sargeant, A.J.; De Haan, A. The Measurement of Force/Velocity Relationships of Fresh and Fatigued Human Adductor Pollicis Muscle. Eur. J. Appl. Physiol. 1999, 80, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Bonde-Petersen, F.; Henriksson, J.; Knuttgen, H.G. Effects of Previous Exercise with Arms or Legs on Metabolism and Performance in Exhaustive Exercise. J. Appl. Physiol. 1975, 38, 763–767. [Google Scholar] [CrossRef]
- Enko, K.; Nakamura, K.; Yunoki, K.; Miyoshi, T.; Akagi, S.; Yoshida, M.; Toh, N.; Sangawa, M.; Nishii, N.; Nagase, S.; et al. Intermittent Arm Ischemia Induces Vasodilatation of the Contralateral Upper Limb. J. Physiol. Sci. 2011, 61, 507–513. [Google Scholar] [CrossRef]
- Paradis-Deschênes, P.; Joanisse, D.R.; Billaut, F. Ischemic Preconditioning Increases Muscle Perfusion, Oxygen Uptake, and Force in Strength-Trained Athletes. Appl. Physiol. Nutr. Metab. 2016, 41, 938–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis-Deschênes, P.; Lapointe, J.; Joanisse, D.R.; Billaut, F. Similar Recovery of Maximal Cycling Performance after Ischemic Preconditioning, Neuromuscular Electrical Stimulation or Active Recovery in Endurance Athletes. J. Sports Sci. Med. 2020, 19, 761–771. [Google Scholar] [PubMed]
- Bailey, T.G.; Birk, G.K.; Cable, N.T.; Atkinson, G.; Green, D.J.; Jones, H.; Thijssen, D.H.J. Remote Ischemic Preconditioning Prevents Reduction in Brachial Artery Flow-Mediated Dilation after Strenuous Exercise. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H533–H538. [Google Scholar] [CrossRef]
- Loukogeorgakis, S.P.; Panagiotidou, A.T.; Broadhead, M.W.; Donald, A.; Deanfield, J.E.; MacAllister, R.J. Remote Ischemic Preconditioning Provides Early and Late Protection Against Endothelial Ischemia-Reperfusion Injury in Humans. J. Am. Coll. Cardiol. 2005, 46, 450–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaver, W.L.; Wasserman, K.; Whipp, B.J. A New Method for Detecting Anaerobic Threshold by Gas Exchange. J. Appl. Physiol. 1986, 60, 2020–2027. [Google Scholar] [CrossRef] [PubMed]
- Marwood, S.; Roche, D.; Rowland, T.; Garrard, M.; Unnithan, V.B. Faster Pulmonary Oxygen Uptake Kinetics in Trained versus Untrained Male Adolescents. Med. Sci. Sport. Exerc. 2010, 42, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Poole, D.C. Oxygen Uptake Dynamics: From Muscle to Mouth—An Introduction to the Symposium. Med. Sci. Sport. Exerc. 2005, 37, 1542–1550. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.J.; Vanhatalo, A.; DiMenna, F.J.; Wilkerson, D.P.; Jones, A.M. Fast-Start Strategy Improves VO2 Kinetics and High-Intensity Exercise Performance. Med. Sci. Sports Exerc. 2011, 43, 457–467. [Google Scholar] [CrossRef]
- Cocking, S.; Landman, T.; Benson, M.; Lord, R.; Jones, H.; Gaze, D.; Thijssen, D.H.J.; George, K. The Impact of Remote Ischemic Preconditioning on Cardiac Biomarker and Functional Response to Endurance Exercise. Scand. J. Med. Sci. Sports 2017, 27, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Sabino-Carvalho, J.L.; Lopes, T.R.; Obeid-Freitas, T.; Ferreira, T.N.; Succi, J.E.; Silva, A.C.; Silva, B.M. Effect of Ischemic Preconditioning on Endurance Performance Does Not Surpass Placebo. Med. Sci. Sport. Exerc. 2017, 49, 124–132. [Google Scholar] [CrossRef]
- Bellini, D.; Chapman, C.; Peden, D.; Hoekstra, S.P.; Ferguson, R.A.; Leicht, C.A. Ischaemic Preconditioning Improves Upper-Body Endurance Performance without Altering VO2 Kinetics. Eur. J. Sport Sci. 2022, 1–9. [Google Scholar] [CrossRef]
- Jones, A.M.; Burnley, M.; Black, M.I.; Poole, D.C.; Vanhatalo, A. The Maximal Metabolic Steady State: Redefining the “Gold Standard”. Physiol. Rep. 2019, 7, e14098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnley, M.; Doust, J.H.; Jones, A.M. Time Required for the Restoration of Normal Heavy Exercise VO2 Kinetics Following Prior Heavy Exercise. J. Appl. Physiol. 2006, 101, 1320–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilding, A.E.; Sequeira, G.M.; Wood, M.R. Effects of Ischemic Preconditioning on Economy, VO2 Kinetics and Cycling Performance in Endurance Athletes. Eur. J. Appl. Physiol. 2018, 119, 961–967. [Google Scholar] [CrossRef]
- Wiggins, C.C.; Constantini, K.; Paris, H.L.; Mickleborough, T.D.; Chapman, R.F. Ischemic Preconditioning, O2 Kinetics, and Performance in Normoxia and Hypoxia. Med. Sci. Sports Exerc. 2019, 51, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Peden, D.L.; Mitchell, E.A.; Bailey, S.J.; Ferguson, R.A. Ischaemic Preconditioning Blunts Exercise-induced Mitochondrial Dysfunction, Speeds Oxygen Uptake Kinetics but Does Not Alter Severe-Intensity Exercise Capacity. Exp. Physiol. 2022, 107, 1241–1254. [Google Scholar] [CrossRef]
- Black, M.I.; Jones, A.M.; Blackwell, J.R.; Bailey, S.J.; Wylie, L.J.; McDonagh, S.T.J.; Thompson, C.; Kelly, J.; Sumners, P.; Mileva, K.N.; et al. Muscle Metabolic and Neuromuscular Determinants of Fatigue during Cycling in Different Exercise Intensity Domains. J. Appl. Physiol. 2017, 122, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.A.; Mills, D.E.; Brown, P.I.; Sharpe, G.R. Prior Upper Body Exercise Reduces Cycling Work Capacity but Not Critical Power. Med. Sci. Sports Exerc. 2014, 46, 802–808. [Google Scholar] [CrossRef] [Green Version]
- Burnley, M.; Davison, G.; Baker, J.R. Effects of Priming Exercise on VO2 Kinetics and the Power-Duration Relationship. Med. Sci. Sports Exerc. 2011, 43, 2171–2179. [Google Scholar] [CrossRef]
- Li, X.-D.; Cheng, Y.-T.; Yang, Y.-J.; Meng, X.-M.; Zhao, J.-L.; Zhang, H.-T.; Wu, Y.-J.; You, S.-J.; Wu, Y.-L. PKA-Mediated ENOS Phosphorylation in the Protection of Ischemic Preconditioning against No-Reflow. Microvasc. Res. 2012, 84, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Tinken, T.M.; Thijssen, D.H.J.; Hopkins, N.; Dawson, E.A.; Cable, N.T.; Green, D.J. Shear Stress Mediates Endothelial Adaptations to Exercise Training in Humans. Hypertension 2010, 55, 312–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.G.; Jones, H.; Gregson, W.; Atkinson, G.; Cable, N.T.; Thijssen, D.H.J. Effect of Ischemic Preconditioning on Lactate Accumulation and Running Performance. Med. Sci. Sport. Exerc. 2012, 44, 2084–2089. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Ueda, K.; Goto, C.; Jitsuiki, D.; Nishioka, K.; Umemura, T.; Noma, K.; Yoshizumi, M.; Chayama, K.; Higashi, Y. Repetition of Ischemic Preconditioning Augments Endothelium-Dependent Vasodilation in Humans: Role of Endothelium-Derived Nitric Oxide and Endothelial Progenitor Cells. ATVB 2007, 27, 1403–1410. [Google Scholar] [CrossRef]
- Pang, C.Y.; Yang, R.Z.; Zhong, A.; Xu, N.; Boyd, B.; Forrest, C.R. Acute Ischaemic Preconditioning Protects against Skeletal Muscle Infarction in the Pig. Cardiovasc. Res. 1995, 29, 782–788. [Google Scholar] [CrossRef]
- Jean-St-Michel, E.; Manlhiot, C.; Li, J.; Tropak, M.; Michelsen, M.M.; Schmidt, M.R.; McCrindle, B.W.; Wells, G.D.; Redington, A.N. Remote Preconditioning Improves Maximal Performance in Highly Trained Athletes. Med. Sci. Sports Exerc. 2011, 43, 1280–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.S.; Downey, J.M. Preconditioning: State of the Art Myocardial Protection. Cardiovasc. Res. 1993, 27, 542–550. [Google Scholar] [CrossRef]
- Hashimoto, T.; Brooks, G.A. Mitochondrial Lactate Oxidation Complex and an Adaptive Role for Lactate Production. Med. Sci. Sport. Exerc. 2008, 40, 486–494. [Google Scholar] [CrossRef]
- Andreas, M.; Schmid, A.I.; Keilani, M.; Doberer, D.; Bartko, J.; Crevenna, R.; Moser, E.; Wolzt, M. Effect of Ischemic Preconditioning in Skeletal Muscle Measured by Functional Magnetic Resonance Imaging and Spectroscopy: A Randomized Crossover Trial. J. Cardiovasc. Magn. Reson. 2011, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Lochner, A.; Genade, S.; Tromp, E.; Podzuweit, T.; Moolman, J.A. Ischemic Preconditioning and the β-Adrenergic Signal Transduction Pathway. Circulation 1999, 100, 958–966. [Google Scholar] [CrossRef] [Green Version]
- Salvador, A.F.; De Aguiar, R.A.; Lisbôa, F.D.; Pereira, K.L.; Cruz, R.S.D.O.; Caputo, F. Ischemic Preconditioning and Exercise Performance: A Systematic Review and Meta-Analysis. Int. J. Sport. Physiol. Perform. 2016, 11, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, T.C.; Machado, A.C.; Braz, I.D.; Fernandes, I.A.; Vianna, L.C.; Nobrega, A.C.L.; Silva, B.M. Remote Ischemic Preconditioning Delays Fatigue Development during Handgrip Exercise: RIPC Improves Handgrip Performance. Scand. J. Med. Sci. Sports 2015, 25, 356–364. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Cruz, R.S.; de Aguiar, R.A.; Turnes, T.; Salvador, A.F.; Caputo, F. Effects of Ischemic Preconditioning on Short-Duration Cycling Performance. Appl. Physiol. Nutr. Metab. 2016, 41, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Gentil, P.; Oliveira, E.; Bottaro, M. Effects of Exercise Order on Upper-Body Muscle Activation and Exercise Performance. J. Strength Cond. Res. 2007, 21, 1082–1086. [Google Scholar]



{kind=link}
{kind=link}
| CON | SHAM | IPC | |
|---|---|---|---|
| Baseline (L·min−1) | 0.99 ± 0.19 | 1.02 ± 0.18 | 0.91 ± 0.28 |
| Amplitude (L·min−1) | 2.51 ± 0.16 | 2.66 ± 0.14 | 2.68 ± 0.23 |
| τVO2 (s) | 55.4 ± 8.8 | 68 ± 21 | 59.3 ± 8.6 |
| Absolute amplitude (L·min−1) | 3.52 ± 0.30 | 3.68 ± 0.29 | 3.58 ± 0.21 |
| CON | SHAM | IPC | |
|---|---|---|---|
| Baseline (μM) | 16.5 ± 6.5 | 21.3 ± 9.3 | 22 ± 12 |
| TDHHb+Mb (s) | 9.8 ± 3.3 | 8.6 ± 3.8 | 10.8 ± 3.9 |
| Amplitude (μM) | 10.7 ± 4.0 | 12 ± 10 | 13.7 ± 8.3 |
| τHHb+Mb (s) | 11.4 ± 1.7 | 14.0 ± 6.9 | 10.6 ± 3.6 |
| Absolute amplitude (μM) | 27.2 ± 9.8 | 33 ± 19 | 35 ± 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angell, P.J.; Marwood, S. Effect of Ischemic Preconditioning (IPC) on Recovery of Exercise Performance Following a Bout of Exercise to Volitional Exhaustion. Physiologia 2023, 3, 394-405. https://doi.org/10.3390/physiologia3030027
Angell PJ, Marwood S. Effect of Ischemic Preconditioning (IPC) on Recovery of Exercise Performance Following a Bout of Exercise to Volitional Exhaustion. Physiologia. 2023; 3(3):394-405. https://doi.org/10.3390/physiologia3030027
Chicago/Turabian StyleAngell, Peter J., and Simon Marwood. 2023. "Effect of Ischemic Preconditioning (IPC) on Recovery of Exercise Performance Following a Bout of Exercise to Volitional Exhaustion" Physiologia 3, no. 3: 394-405. https://doi.org/10.3390/physiologia3030027