
Significant Differences in the Reversal of Cellular Stress Induced by Hydrogen Peroxide and Corticosterone by the Application of Mirtazapine or L-Tryptophan


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Treatments
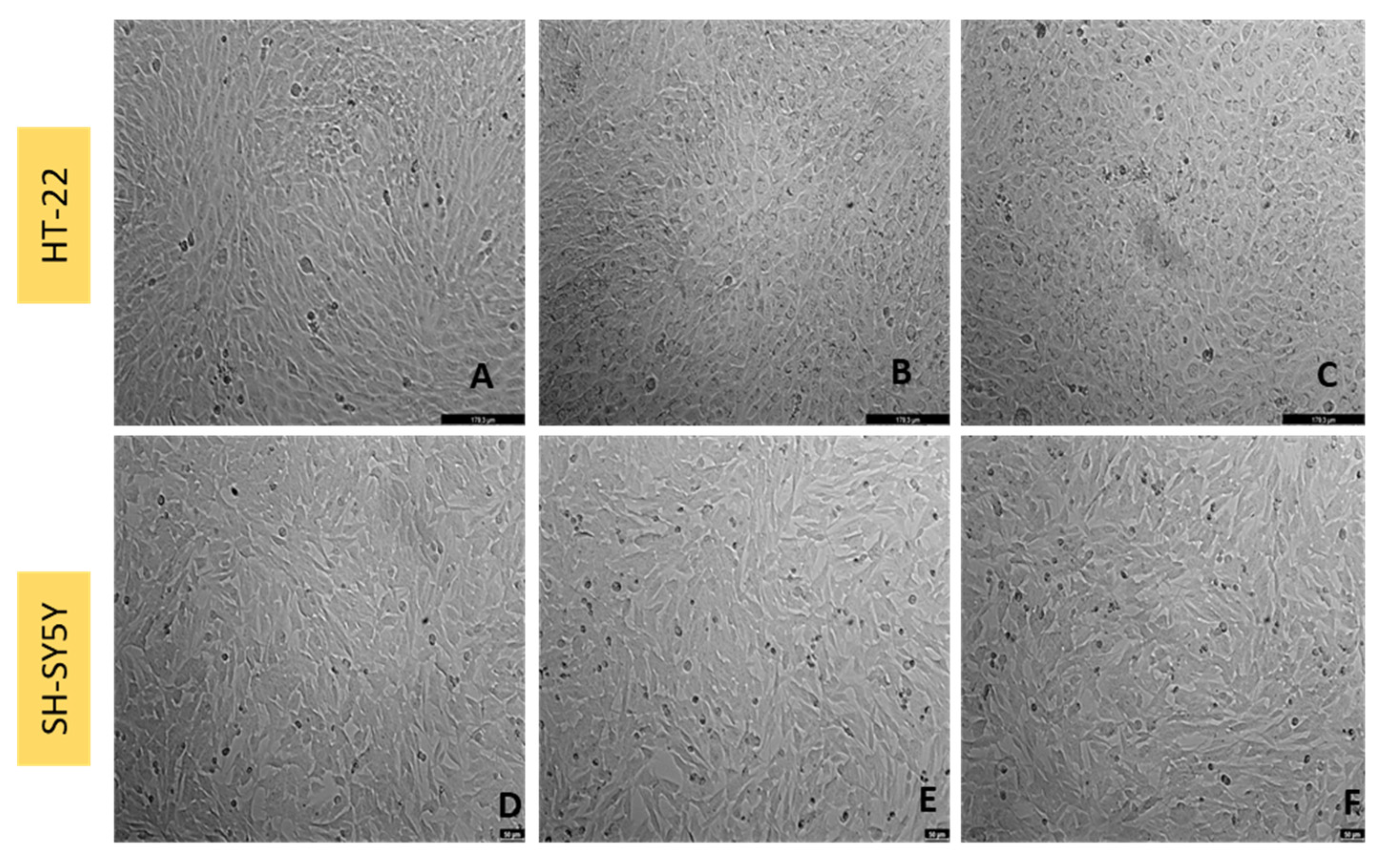
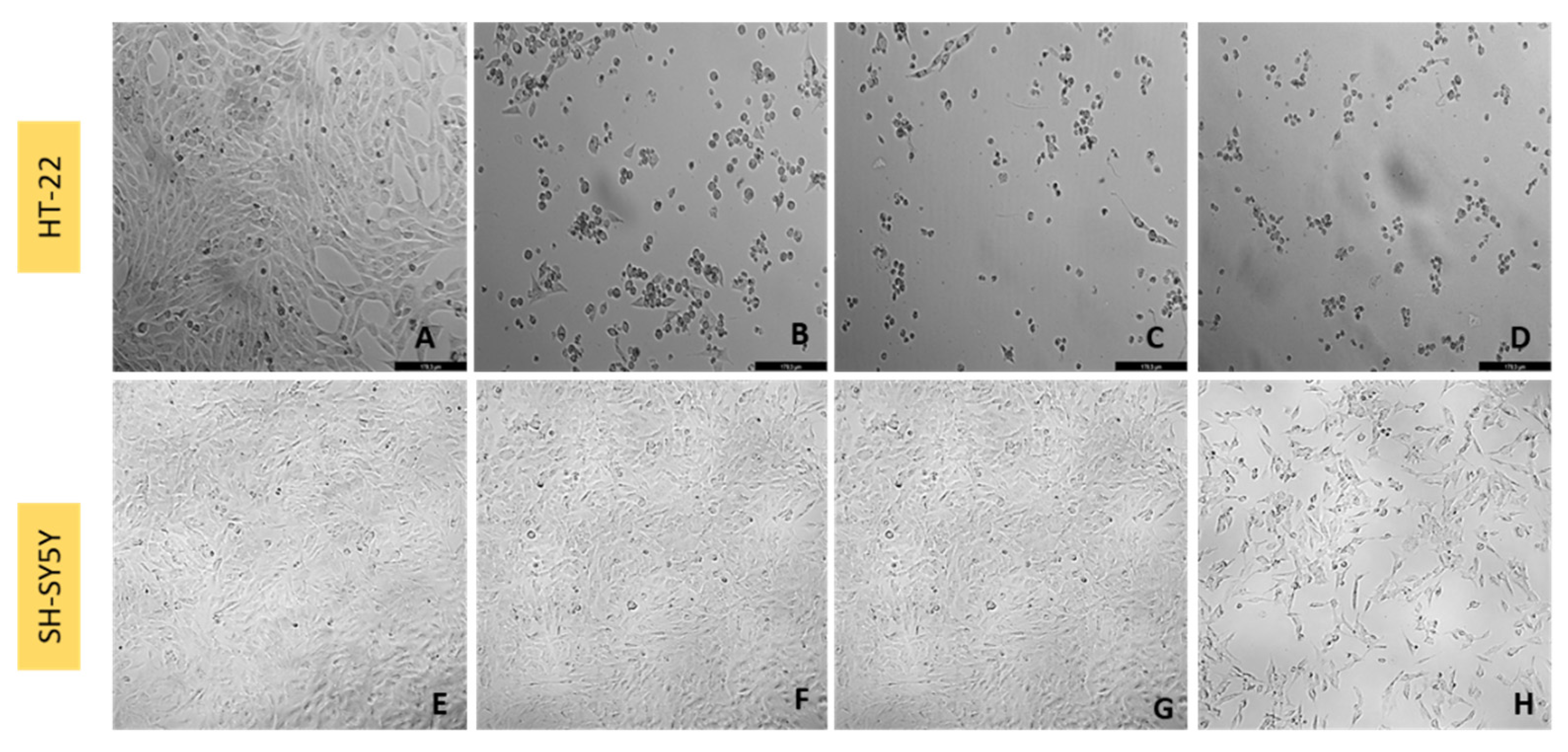
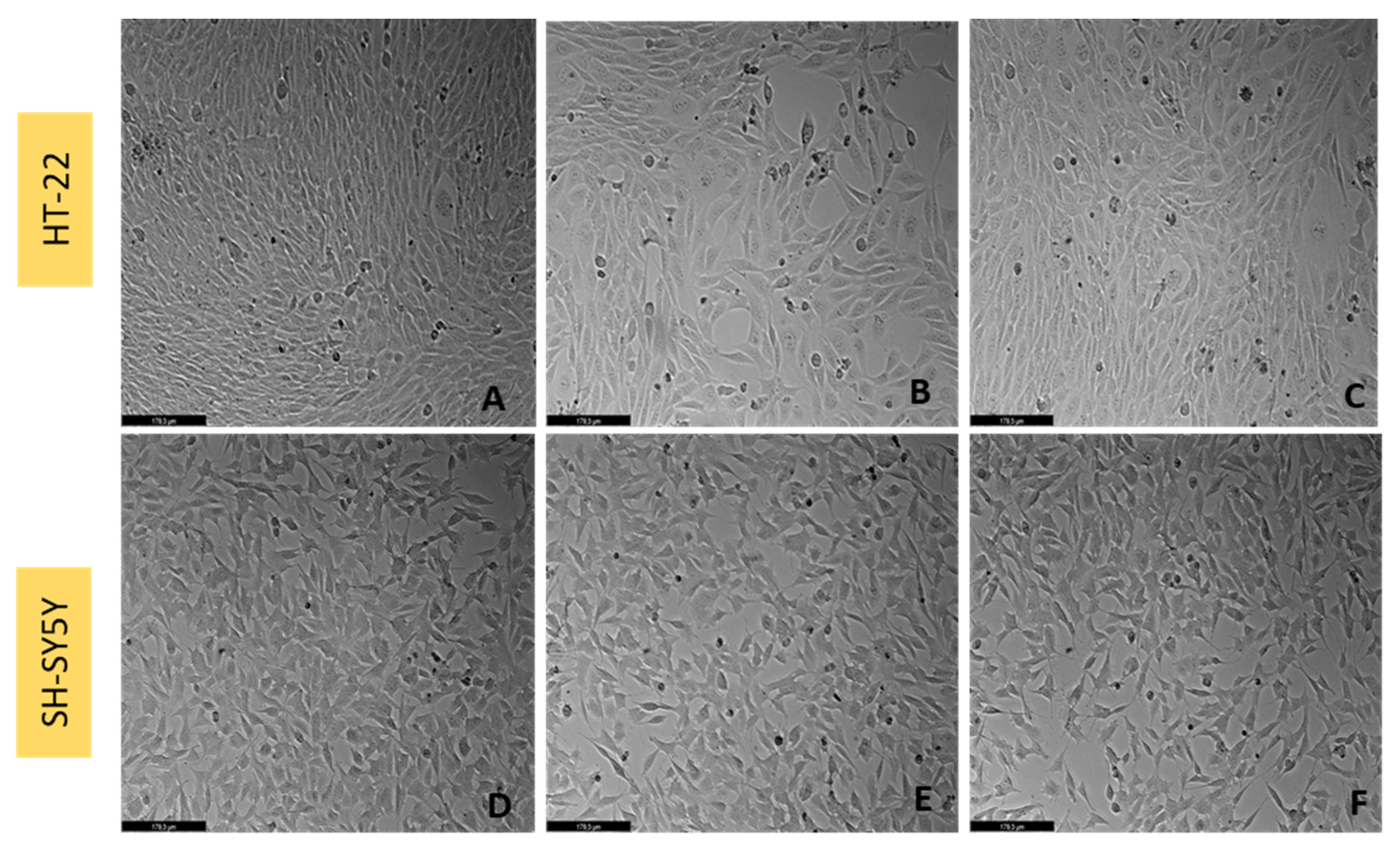
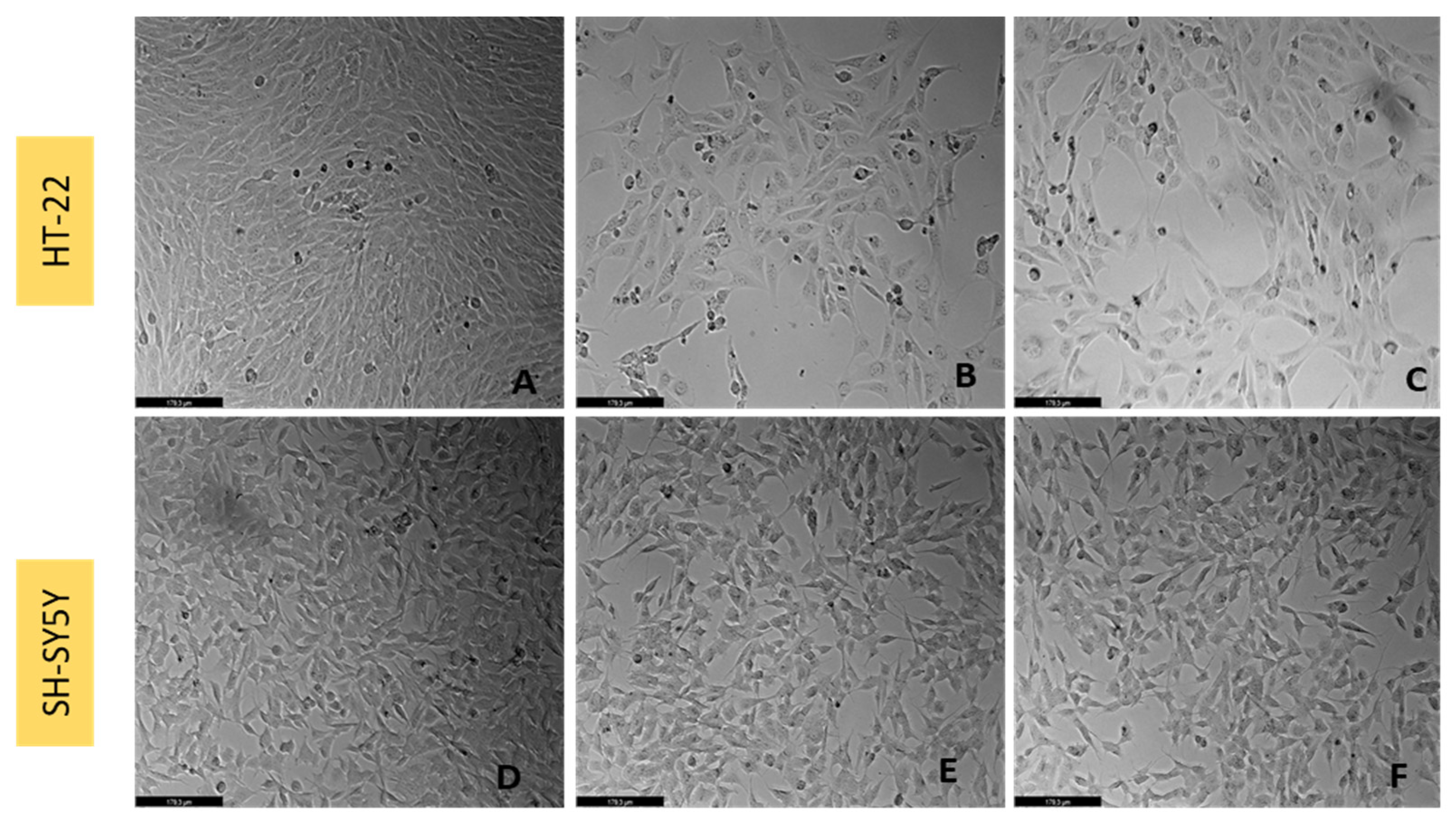
2.4. Cell Morphology Visualization
2.5. MTT and Neutral Red Assays
2.6. DCFDA Assay
2.7. Statistical and Data Analyses
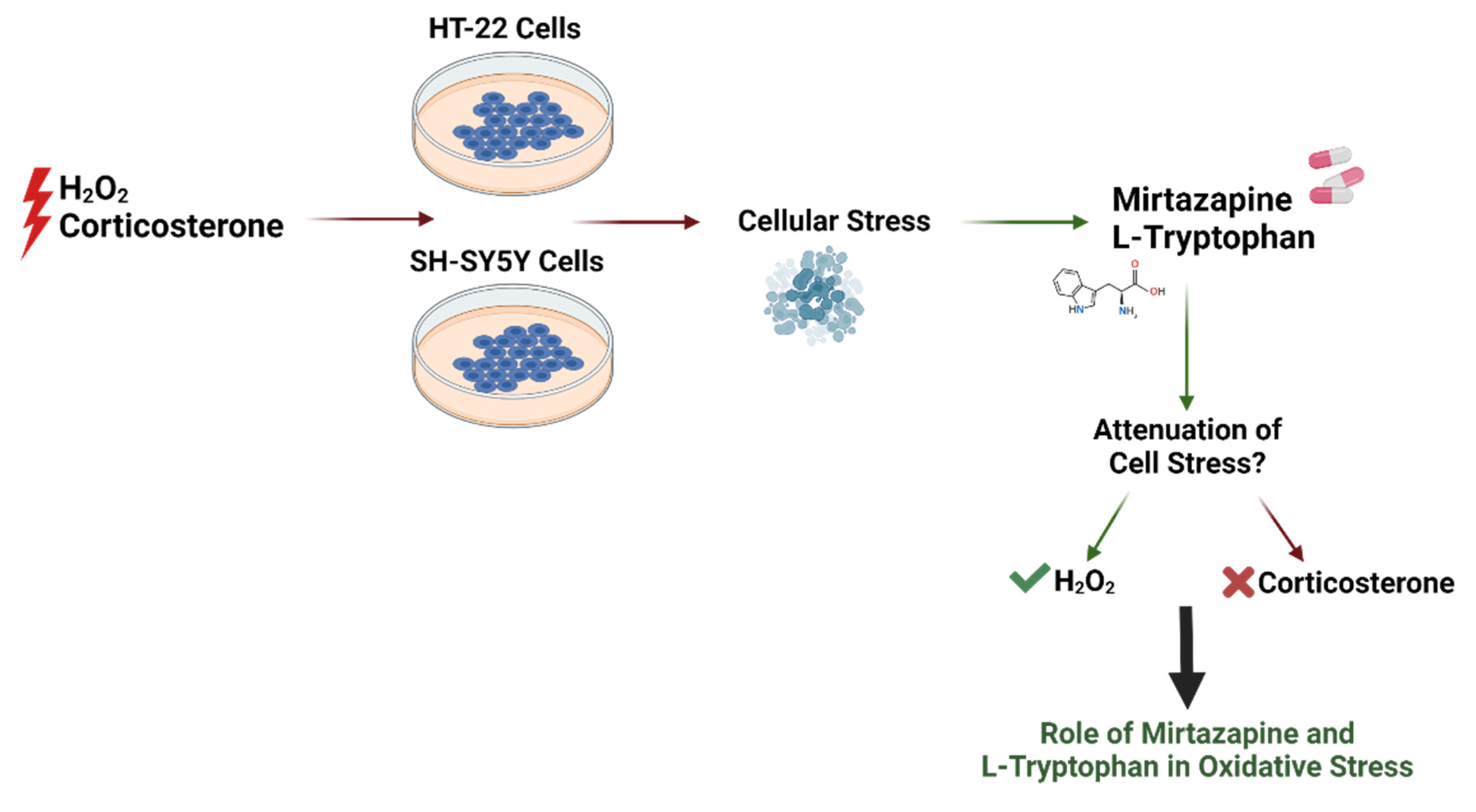
3. Results
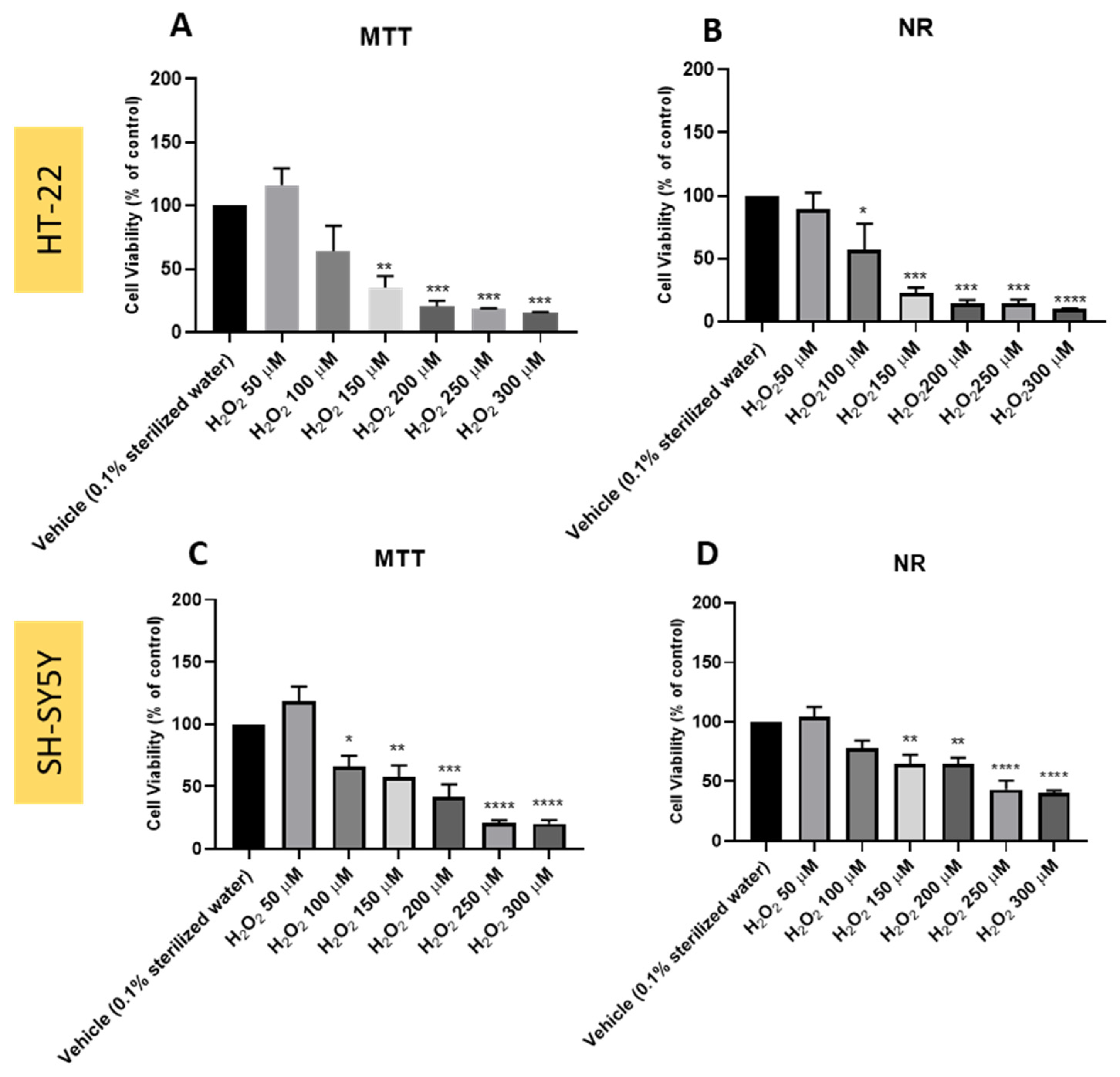
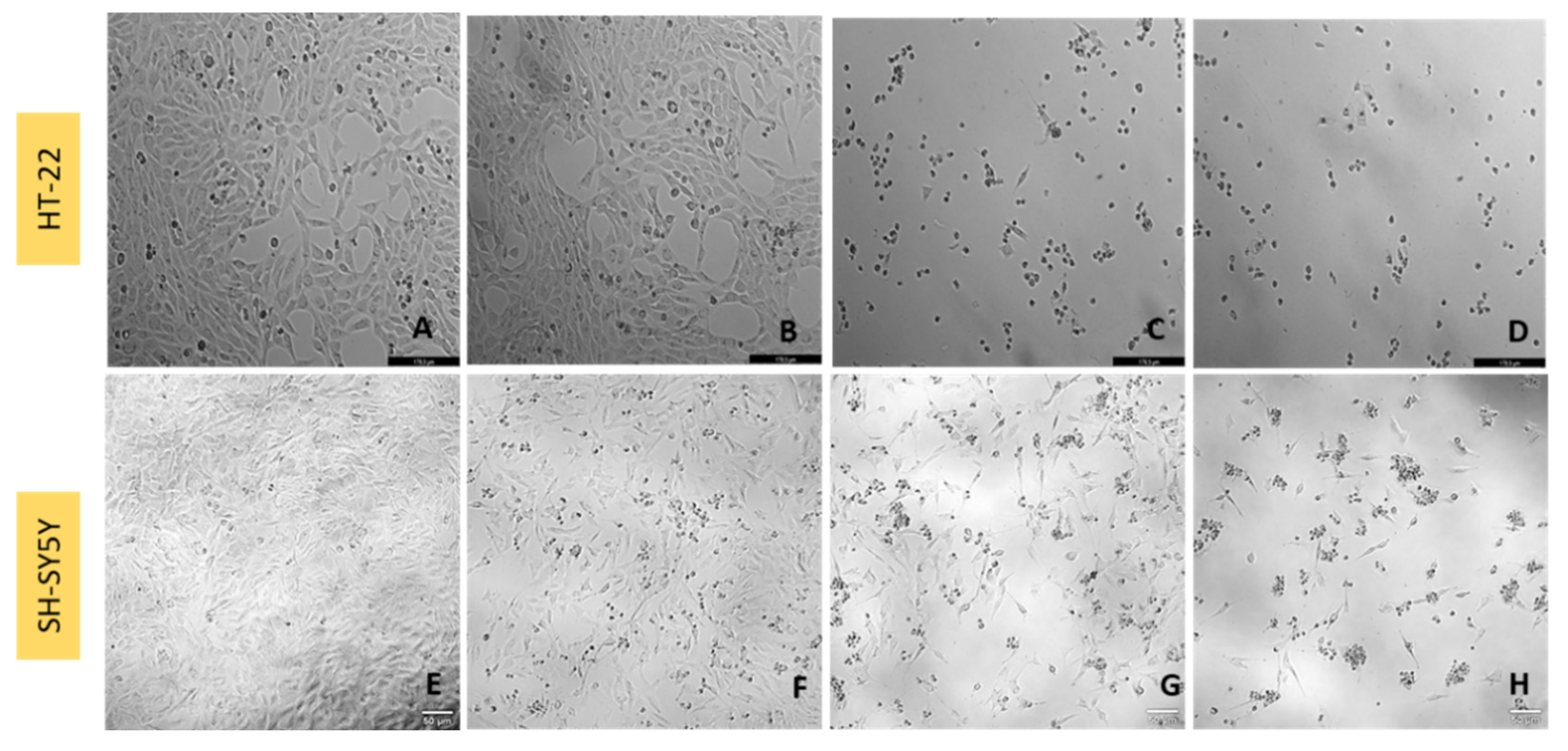
3.1. Effect of Hydrogen Peroxide on SH-SY5Y and HT-22 Cellular Viability
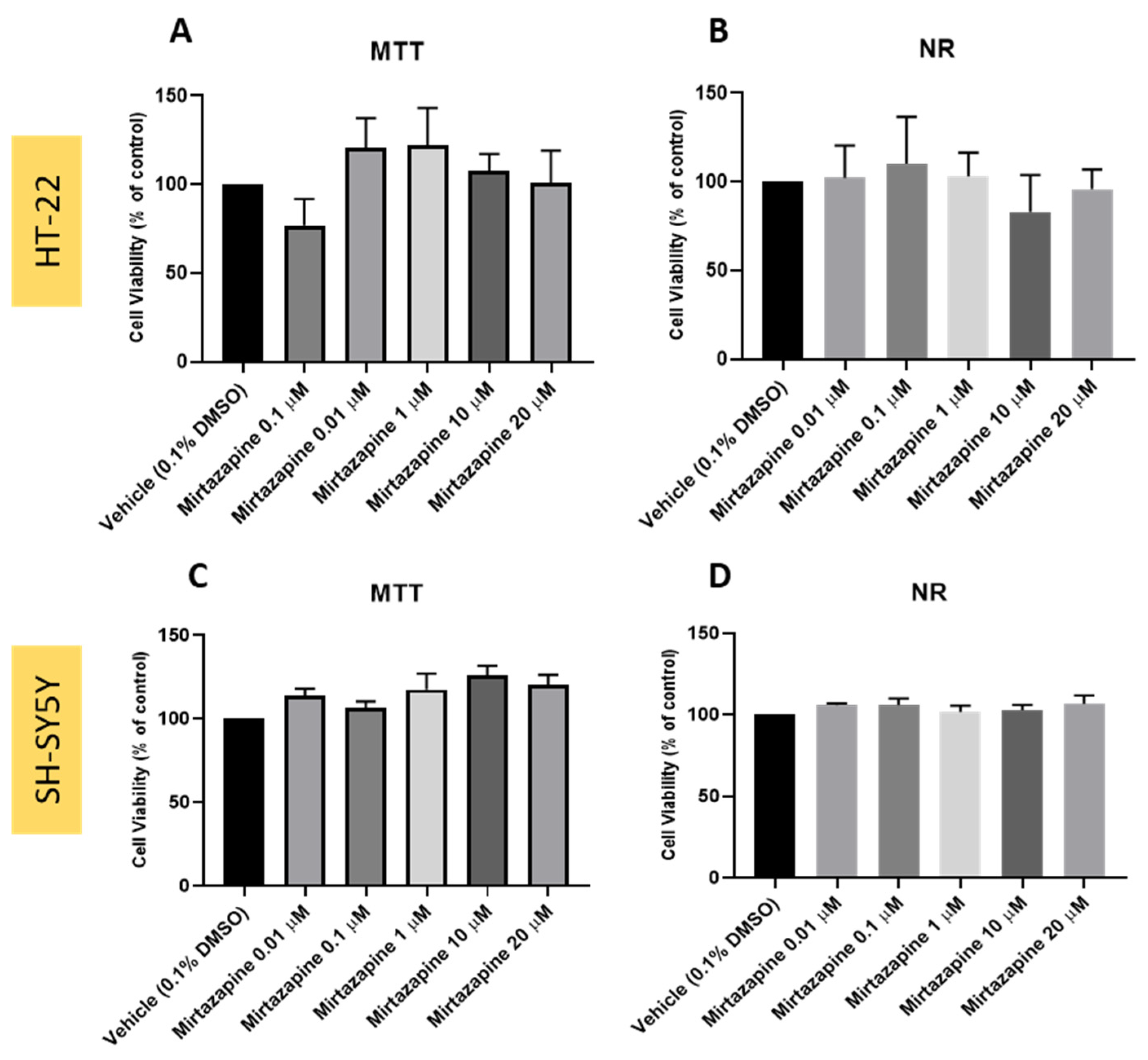
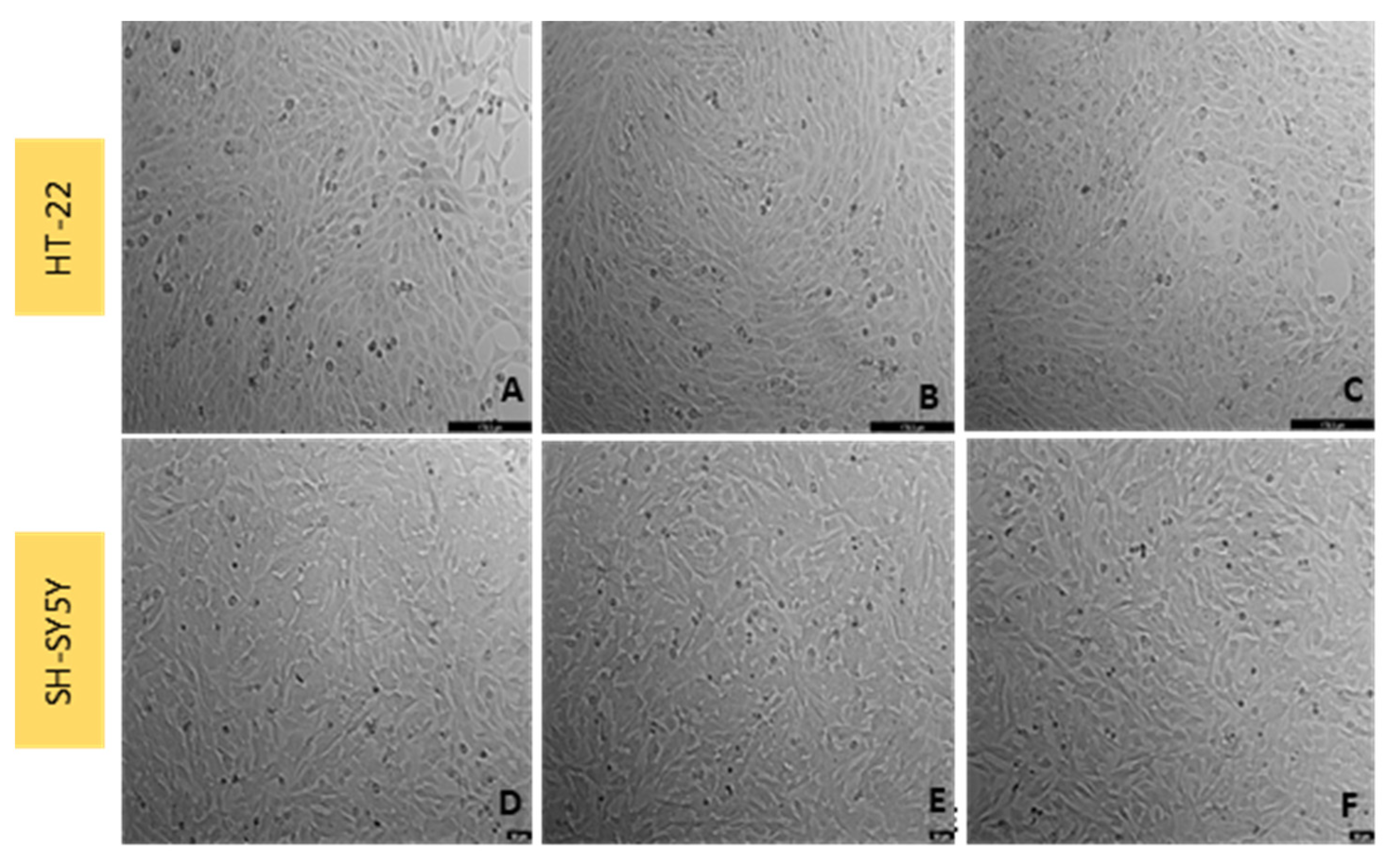
3.2. Effect of Mirtazapine on SH-SY5Y and HT-22 Cellular Viability
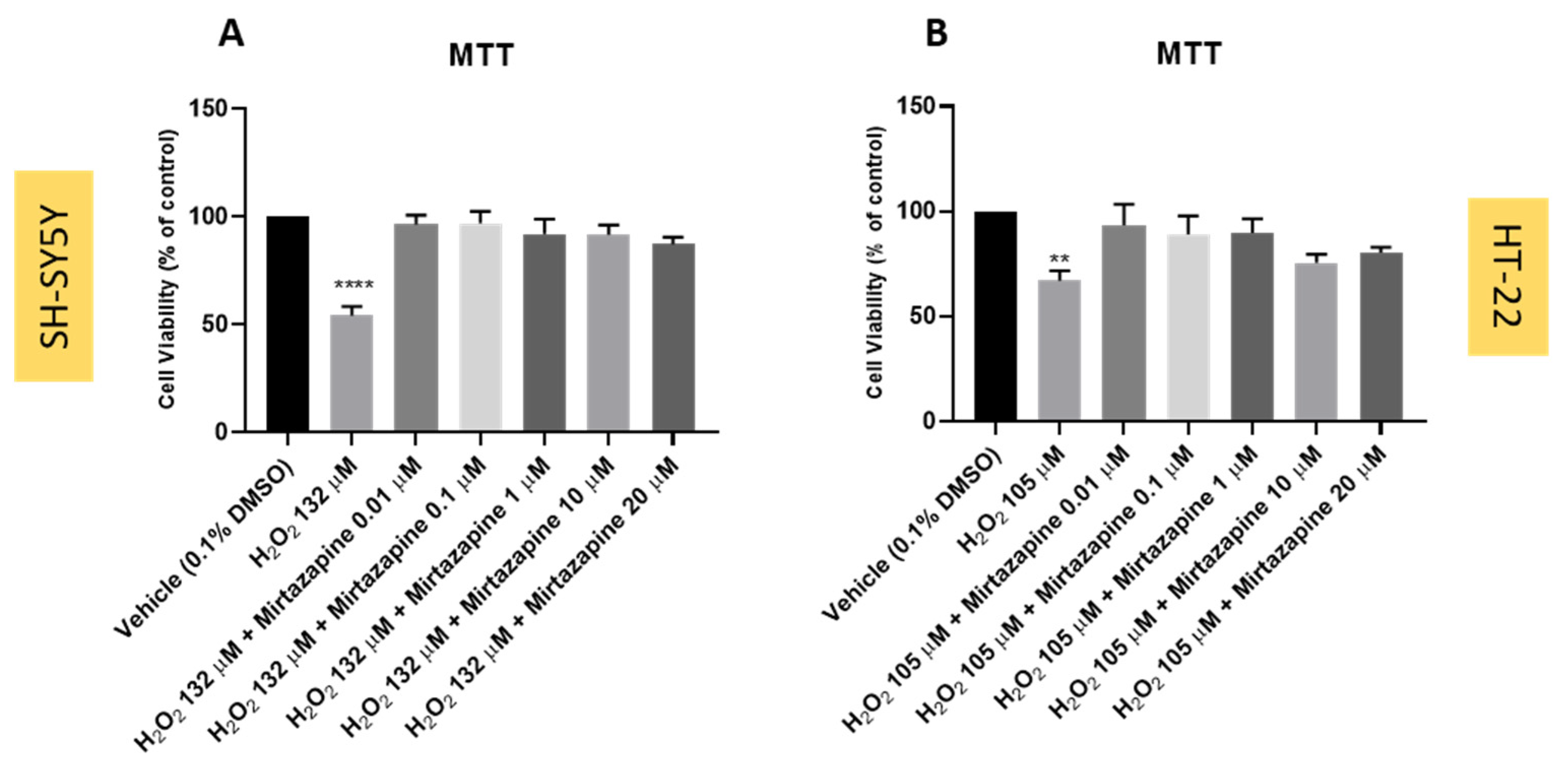
3.3. Effect of Mirtazapine Combined with Hydrogen Peroxide on SH-SY5Y and HT-22 Cellular Viability
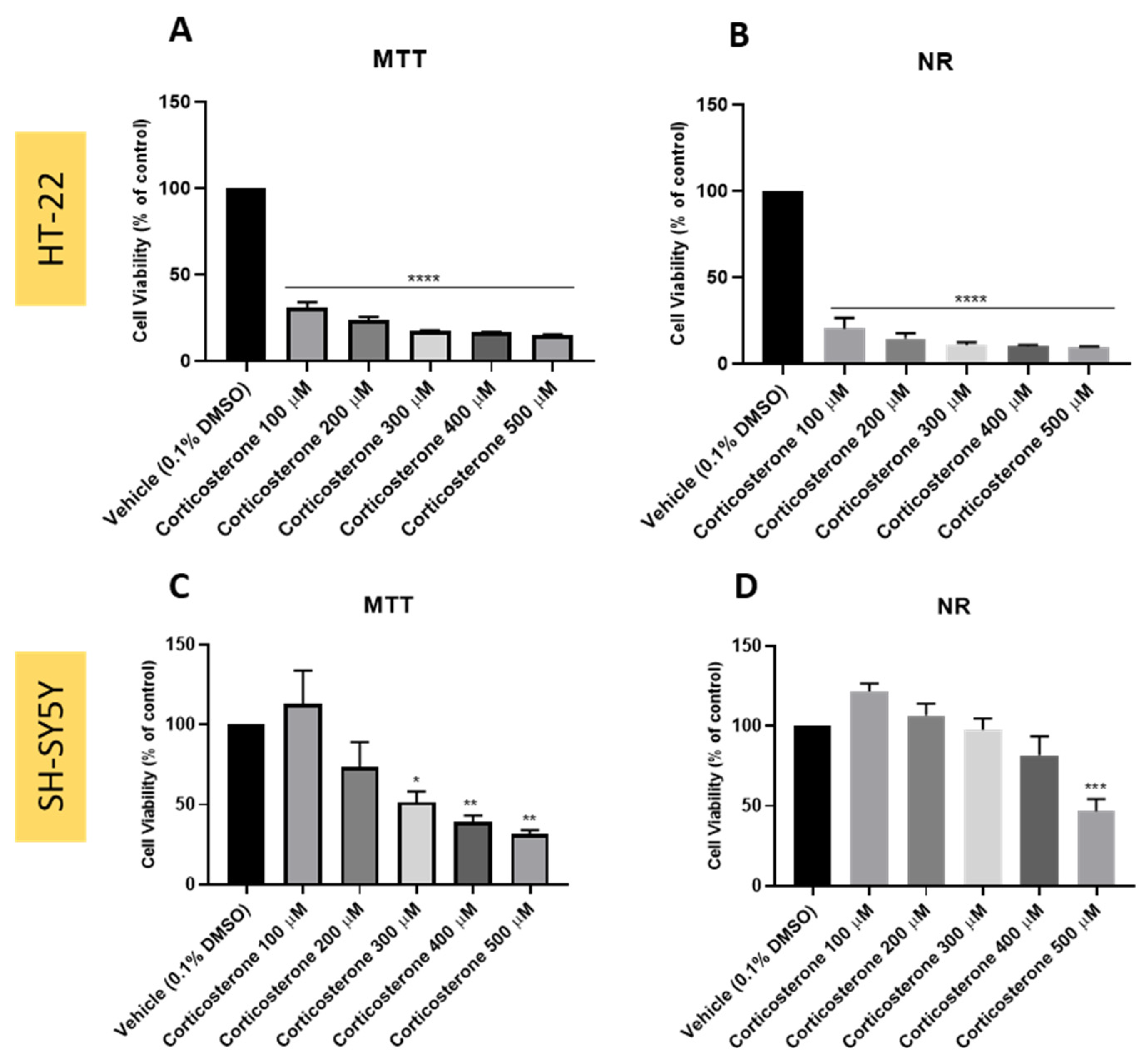
3.4. Effect of Corticosterone on SH-SY5Y and HT-22 Cellular Viability
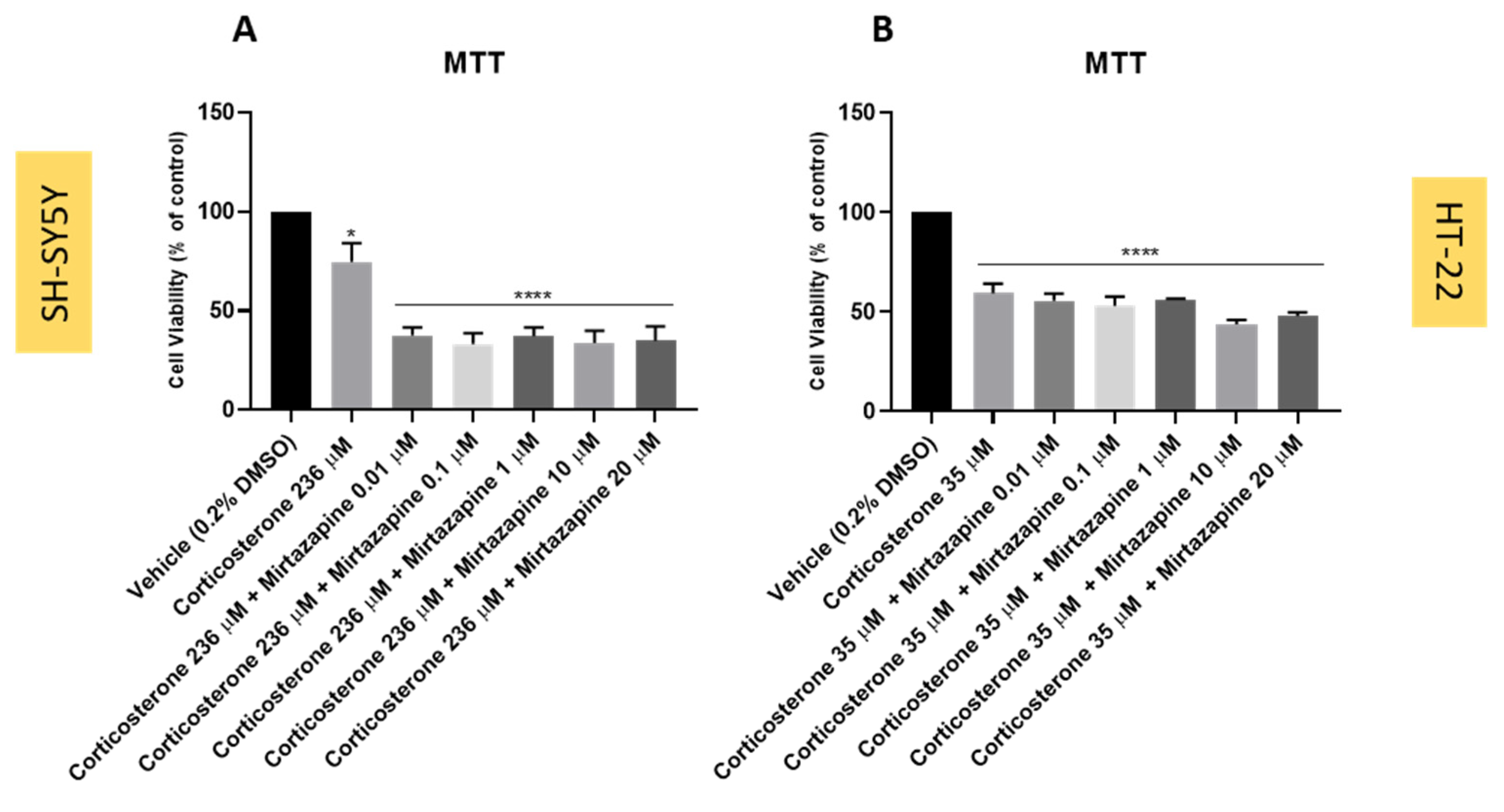
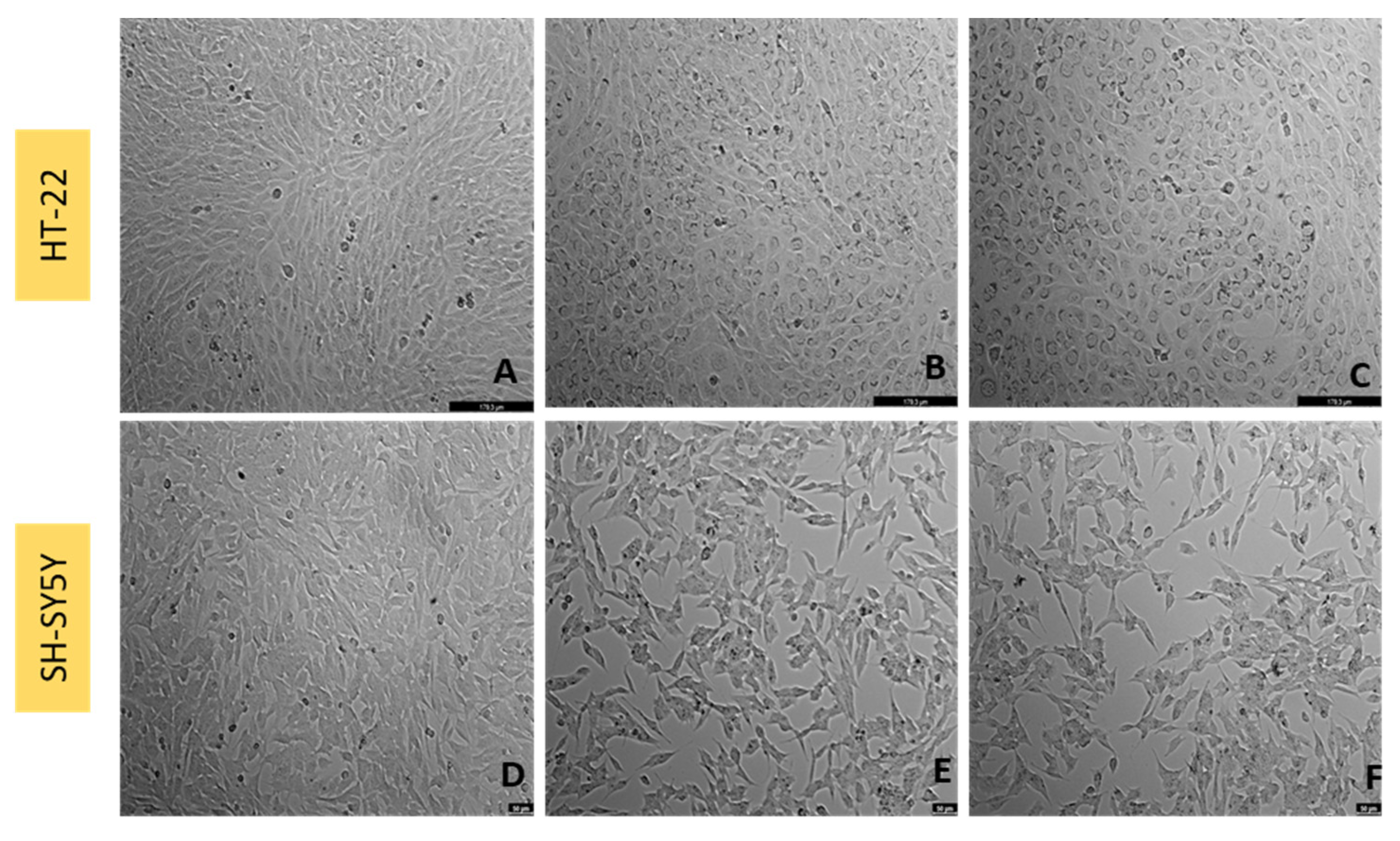
3.5. Effect of Mirtazapine Combined with Corticosterone on SH-SY5Y and HT-22 Cellular Viability
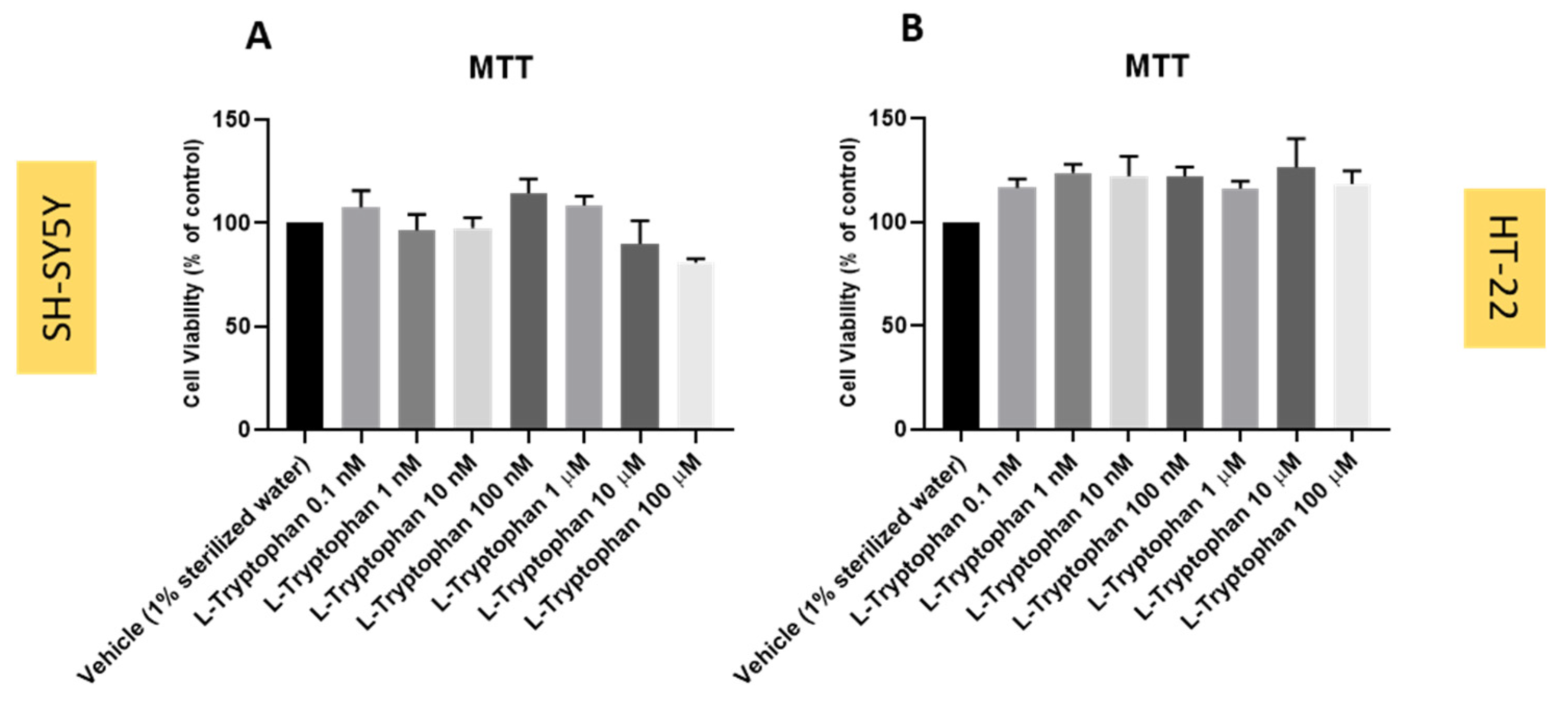
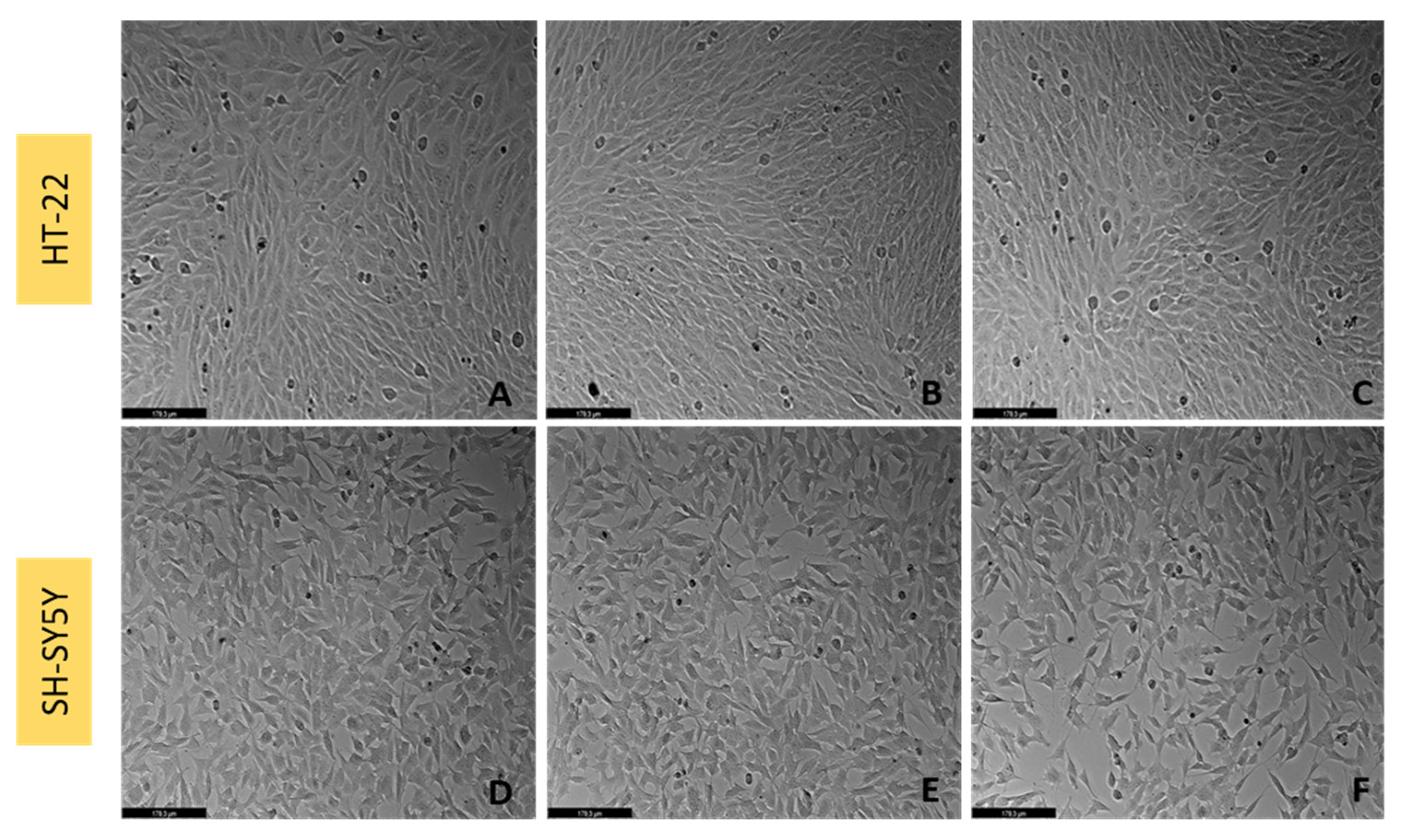
3.6. Effect of L-Tryptophan on SH-SY5Y and HT-22 Cellular Viability
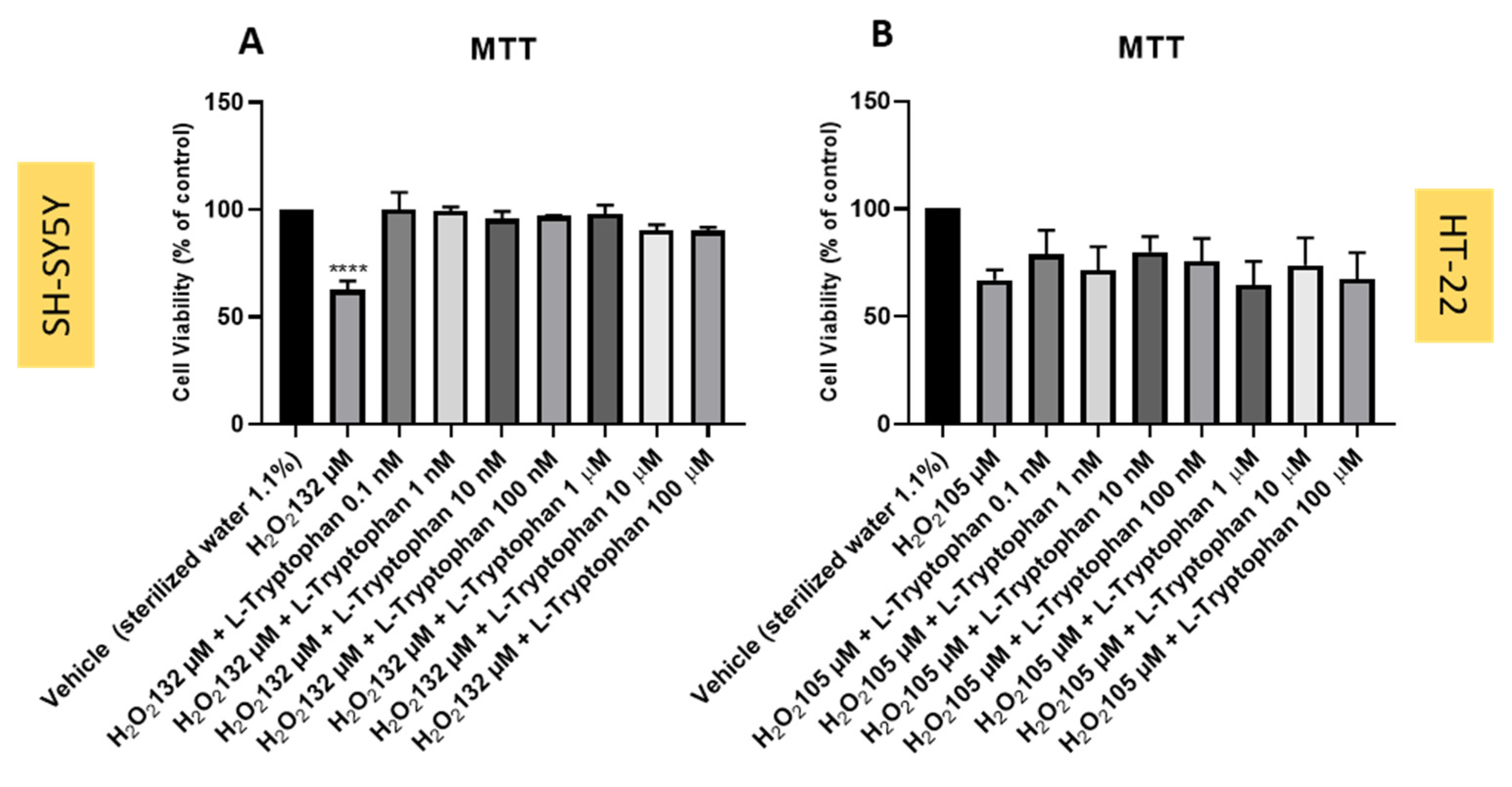
3.7. Effect of L-Tryptophan Combined with Hydrogen Peroxide on SH-SY5Y and HT-22 Cellular Viability
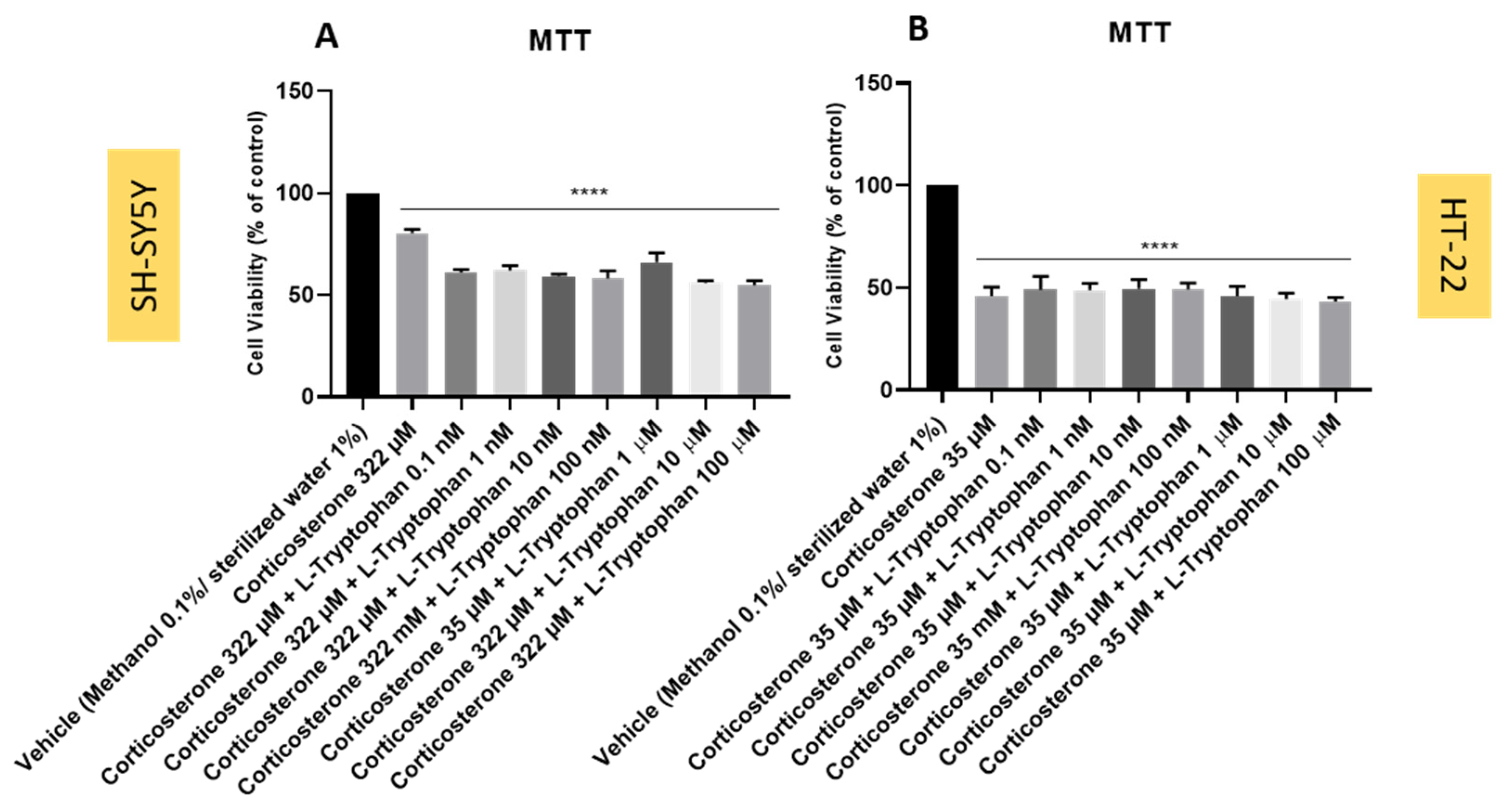
3.8. Effect of L-Tryptophan Combined with Corticosterone on SH-SY5Y and HT-22 Cellular Viability
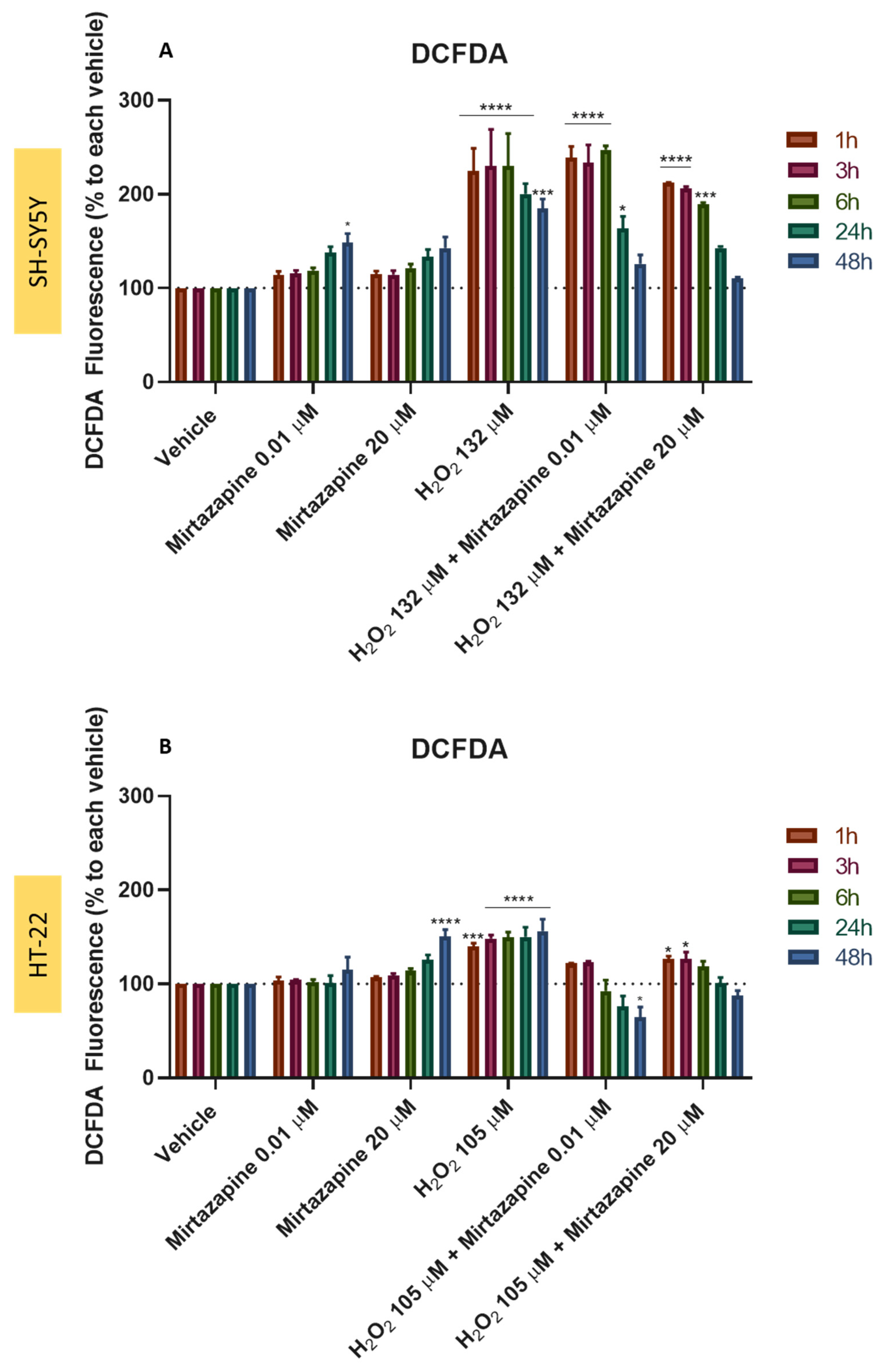
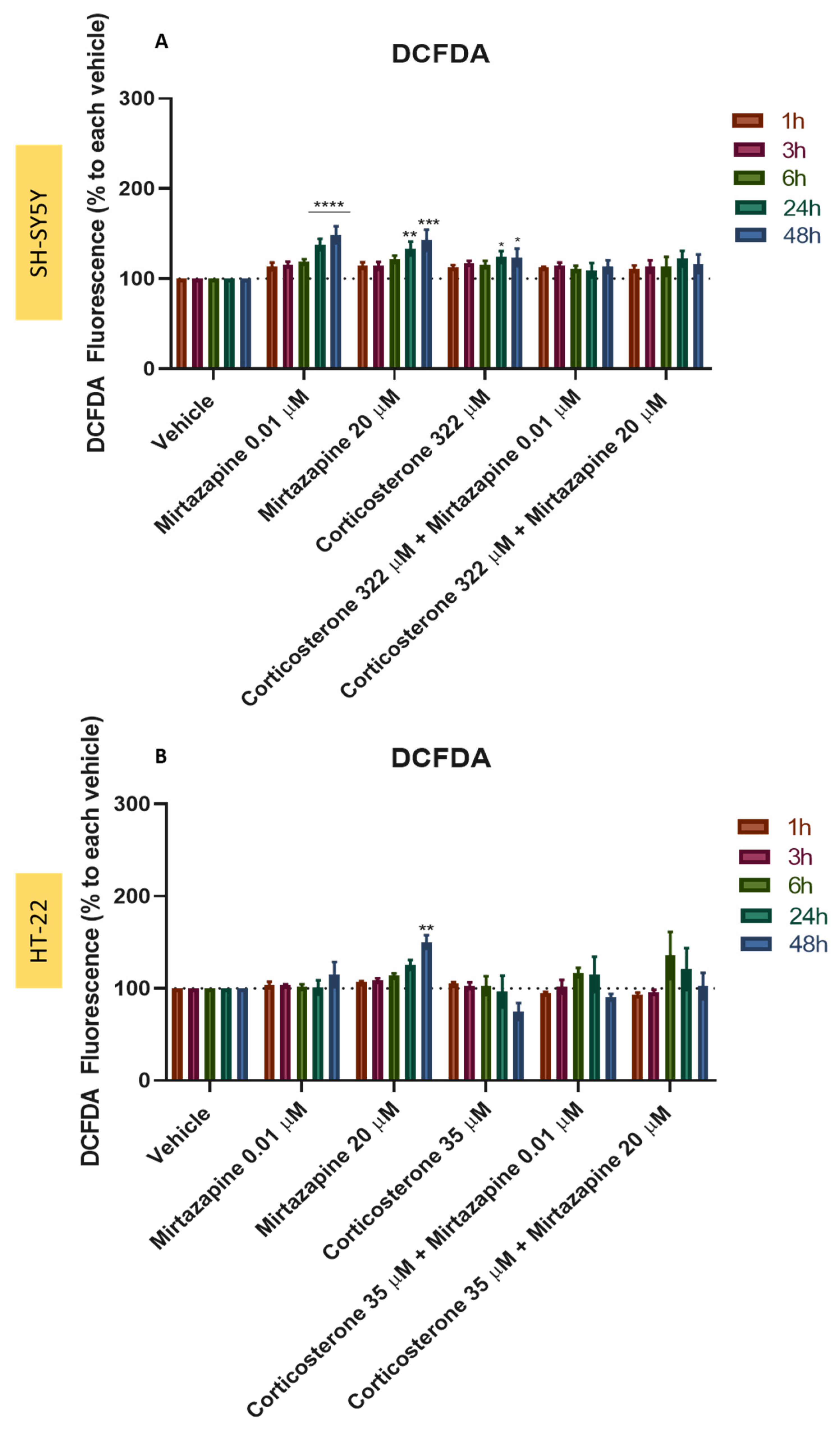
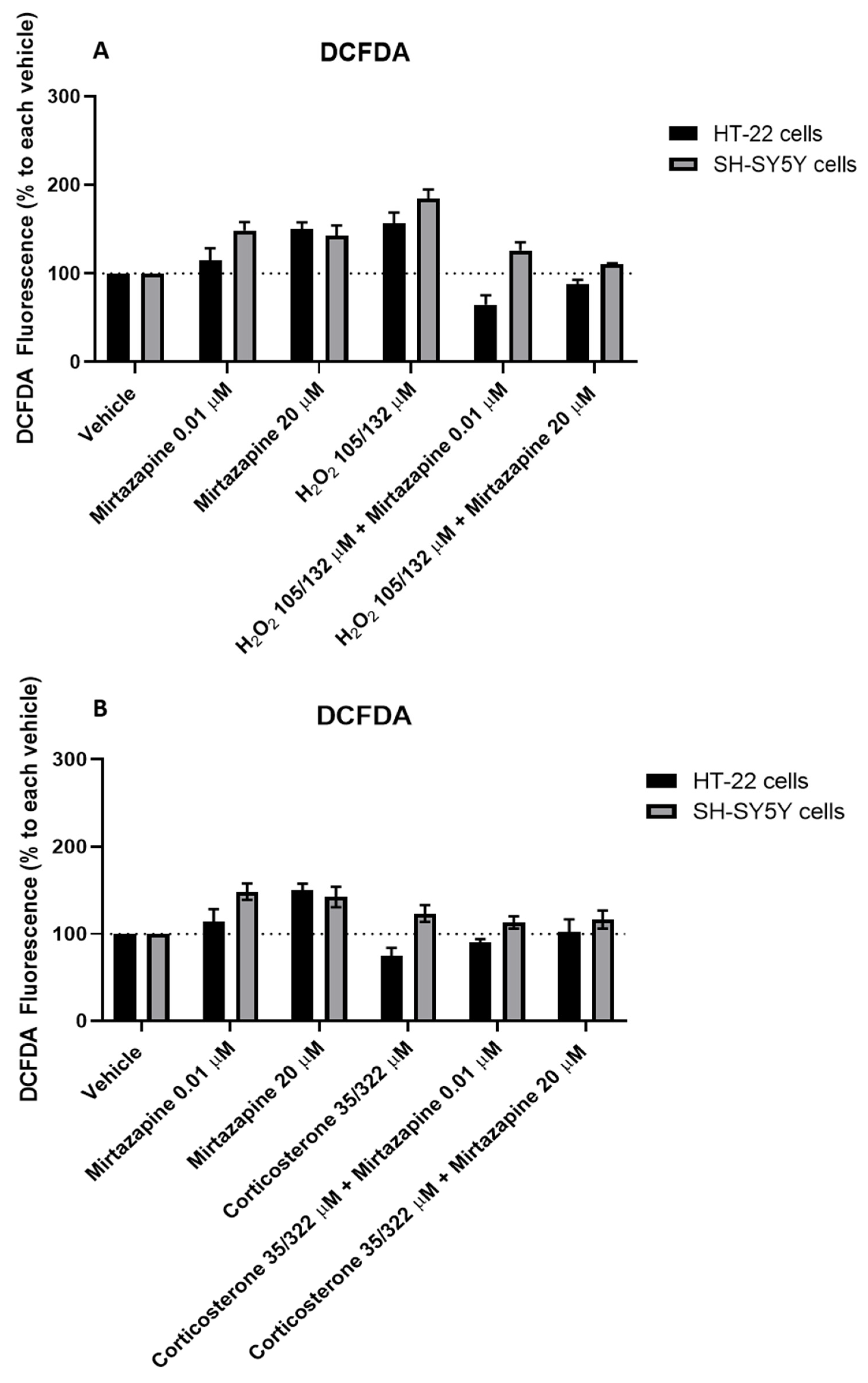
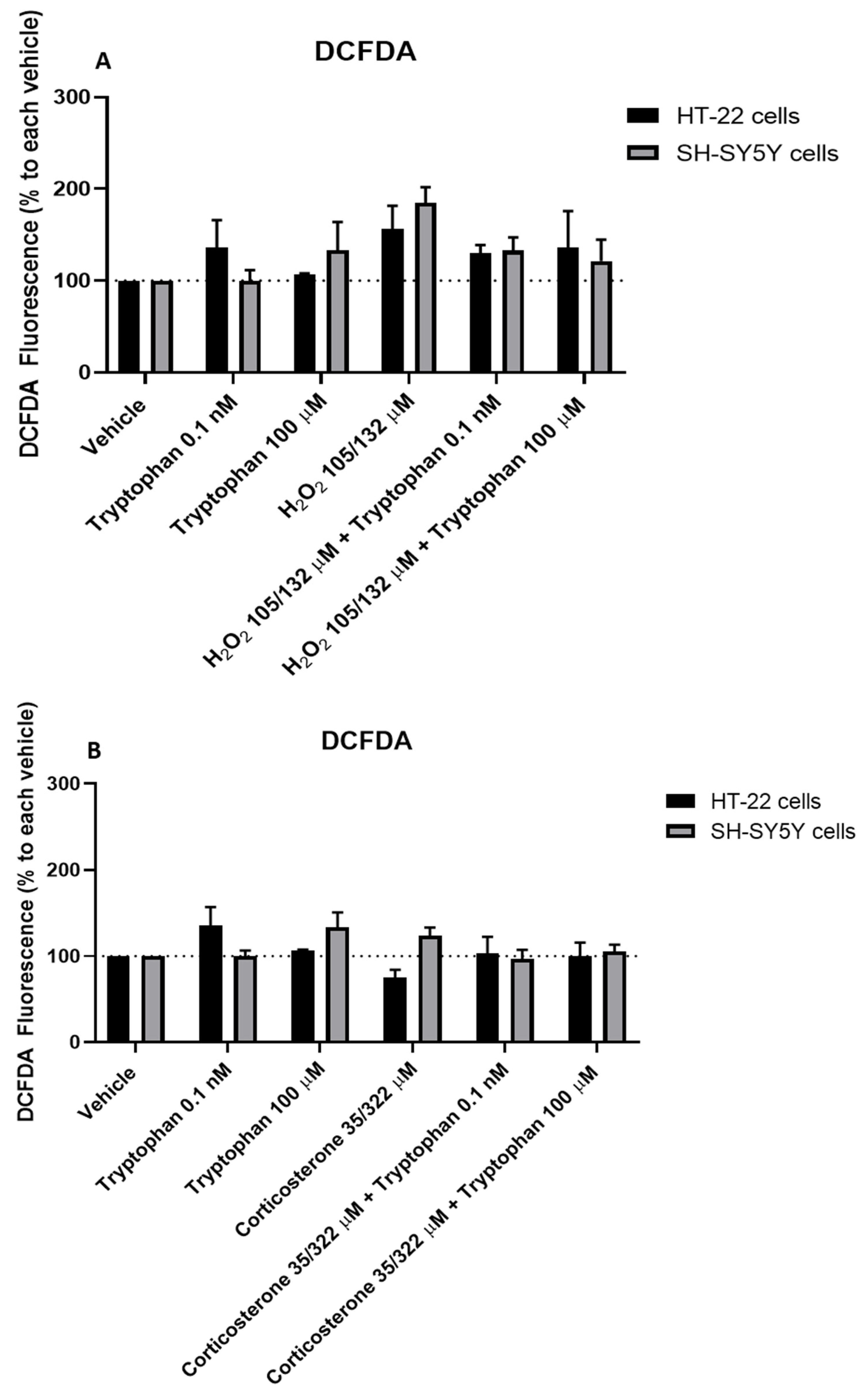
3.9. Effect of Mirtazapine Combined with H2O2 and Corticosterone on SH-SY5Y and HT-22 ROS Production
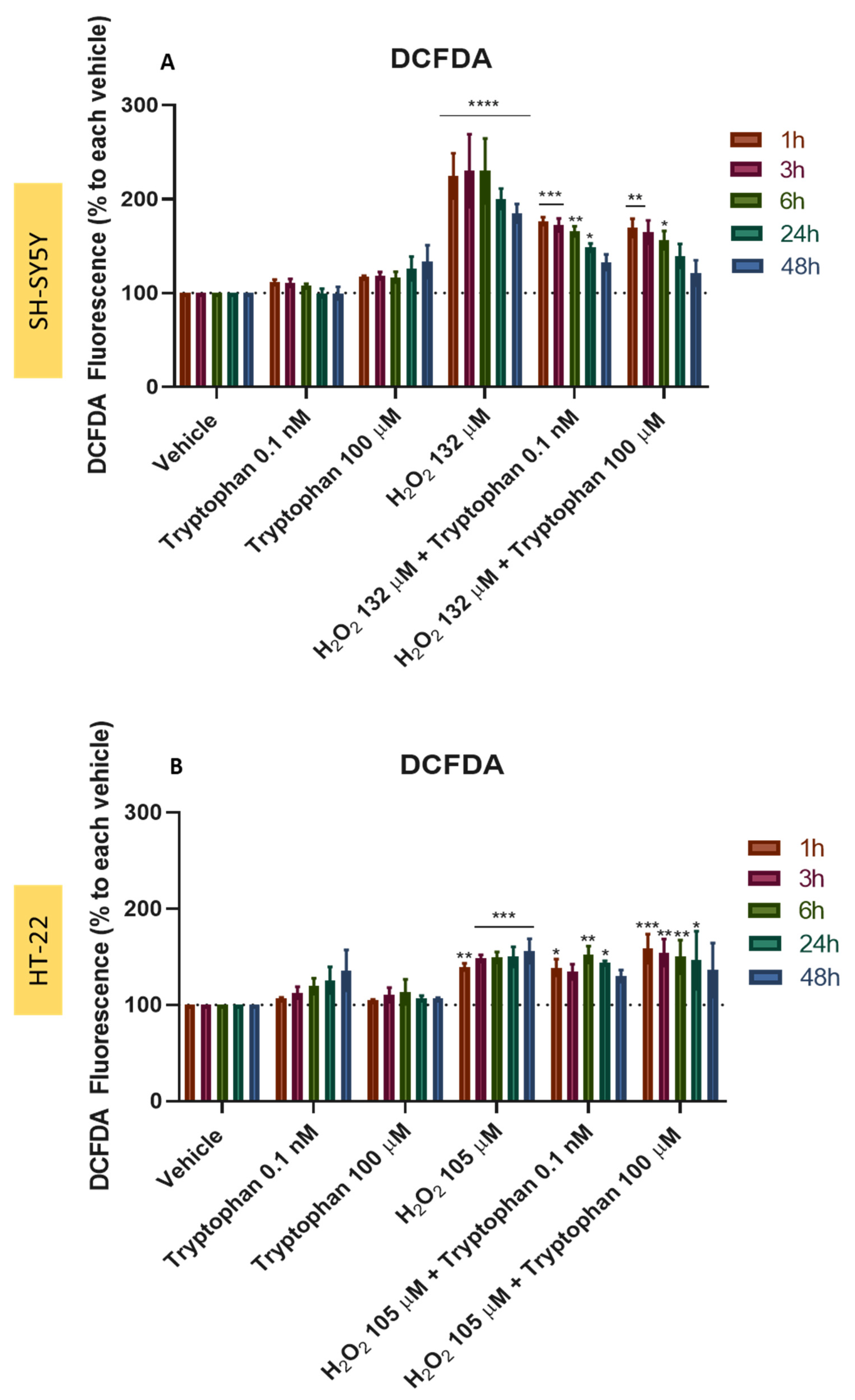
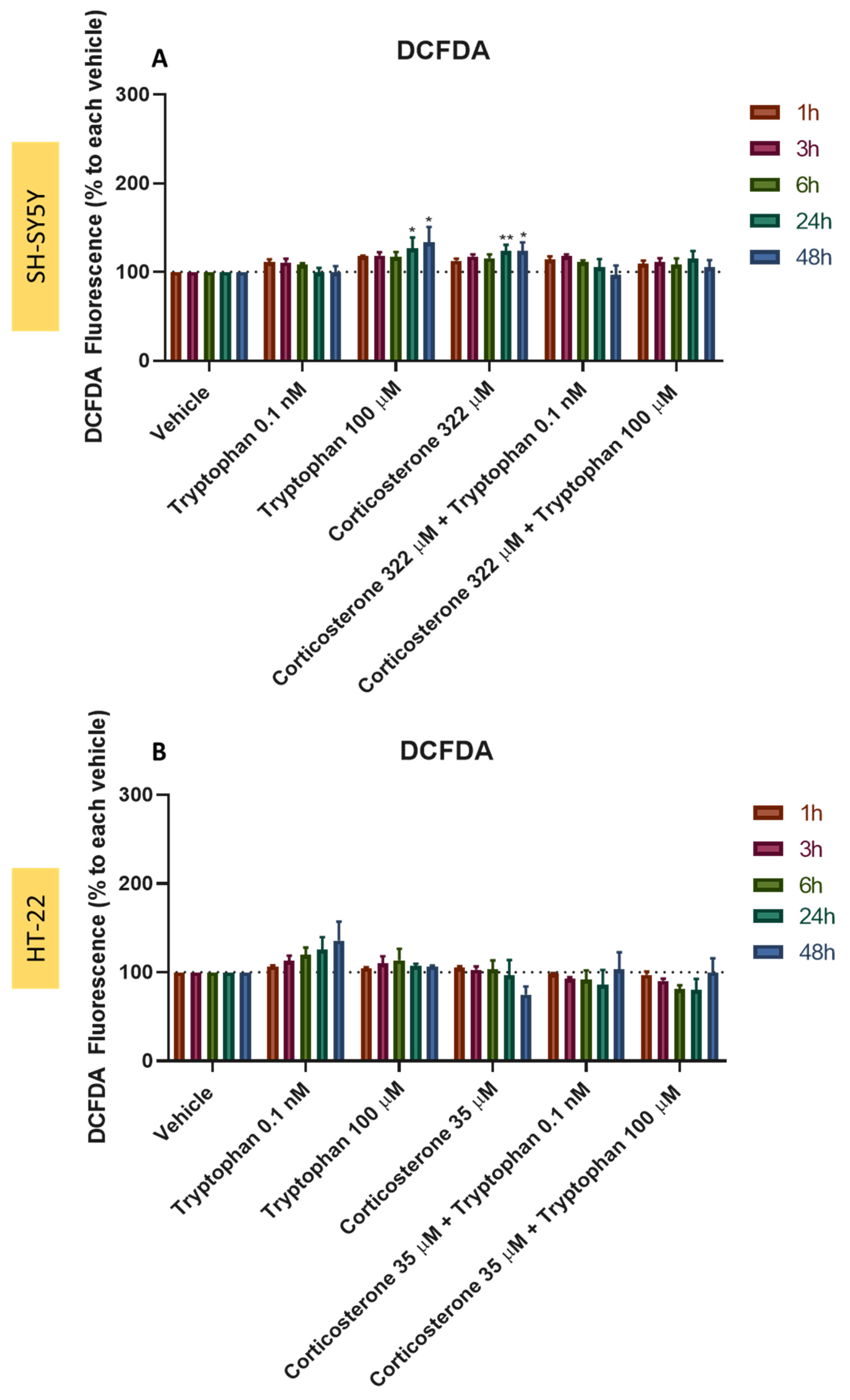
3.10. Effect of L-Tryptophan Combined with H2O2 and Corticosterone on SH-SY5Y and HT-22 ROS Production
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nuggerud-Galeas, S.; Sáez-Benito Suescun, L.; Berenguer Torrijo, N.; Sáez-Benito Suescun, A.; Aguilar-Latorre, A.; Magallón Botaya, R.; Oliván Blázquez, B. Analysis of depressive episodes, their recurrence and pharmacologic treatment in primary care patients: A retrospective descriptive study. PLoS ONE 2020, 15, e0233454. [Google Scholar] [CrossRef] [PubMed]
- Otte, C.; Gold, S.M.; Penninx, B.W.; Pariante, C.M.; Etkin, A.; Fava, M.; Mohr, D.C.; Schatzberg, A.F. Major depressive disorder. Nat. Rev. Dis. Prim. 2016, 2, 16065. [Google Scholar] [CrossRef] [PubMed]
- Jantas, D. Cell-Based Systems of Depression: An Overview. In [3] Medicine in Depression; Springer International Publishing: Cham, Switzerland, 2016; pp. 75–117. ISBN 9783319140216. [Google Scholar]
- Ogawa, Y.; Kobayashi, T.; Nishioka, A.; Kariya, S.; Ohnishi, T.; Hamasato, S.; Seguchi, H.; Yoshida, S. Reactive oxygen species-producing site in hydrogen peroxide-induced apoptosis of human peripheral T cells: Involvement of lysosomal membrane destabilization. Int. J. Mol. Med. 2004, 13, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Yin, J.; Luo, Y.; Huang, S. Hydrogen peroxide-induced neuronal apoptosis is associated with inhibition of protein phosphatase 2A and 5, leading to activation of MAPK pathway. Int. J. Biochem. Cell Biol. 2009, 41, 1284–1295. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, A.; Verma, A.K.; Srivastava, M.; Srivastava, R. Oxidative Stress and Major Depression. J. Clin. Diagn. Res. 2014, 8, CC04–CC07. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Maker, G.L.; Hood, S.D.; Drummond, P.D. A review of peripheral biomarkers in major depression: The potential of inflammatory and oxidative stress biomarkers. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 102–111. [Google Scholar] [CrossRef]
- Horowitz, M.A.; Zunszain, P.A.; Anacker, C.; Musaelyan, K.; Pariante, C.M. Glucocorticoids and Inflammation: A Double-Headed Sword in Depression? Inflamm. Psychiatry 2013, 28, 127–143. [Google Scholar] [CrossRef]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef]
- Qin, D.; Li, Z.; Li, Z.; Wang, L.; Hu, Z.; Lü, L.; Wang, Z.; Liu, Y.; Yin, Y.; Li, Z.; et al. Chronic Glucocorticoid Exposure Induces Depression-Like Phenotype in Rhesus macaque (Macaca mulatta). Front. Neurosci. 2019, 13, 188. [Google Scholar] [CrossRef]
- Berger, S.; Gureczny, S.; Reisinger, S.N.; Horvath, O.; Pollak, D.D. Effect of Chronic Corticosterone Treatment on Depression-Like Behavior and Sociability in Female and Male C57BL/6N Mice. Cells 2019, 8, 1018. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Li, J.; Cai, Y.; Yang, Z.; Li, R.; Fu, W. Glycyrrhizic Acid Alleviates 6-Hydroxydopamine and Corticosterone-Induced Neurotoxicity in SH-SY5Y Cells Through Modulating Autophagy. Neurochem. Res. 2018, 43, 1914–1926. [Google Scholar] [CrossRef] [PubMed]
- Duksal, F.; Kilic, I.; Tufan, A.C.; Akdogan, I. Effects of different corticosteroids on the brain weight and hippocampal neuronal loss in rats. Brain Res. 2009, 1250, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, Y.; Deng, N.; Chen, Y.; Huang, J.; Xie, W. Protective effect of resveratrol against corticosterone-induced neurotoxicity in Pc12 cells. Transl. Neurosci. 2019, 10, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Kong, D.; Liang, Y.; Zhao, X.; Du, G. Protective effects of VMY-2-95 on corticosterone-induced injuries in mice and cellular models. Acta Pharm. Sin. B 2021, 11, 1903–1913. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Lee, G.H.; Hur, J.; Lee, H.G.; Kim, E.; Won, J.P.; Cho, Y.; Choi, M.J.; Seo, H.G. Taurine and Ginsenoside Rf Induce BDNF Expression in SH-SY5Y Cells: A Potential Role of BDNF in Corticosterone-Triggered Cellular Damage. Molecules 2020, 25, 2819. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Lee, H.; Park, H.; Jeon, J.W.; Cho, W.-K.; Ma, J.Y. Neuroprotective effects of Liriope platyphylla extract against hydrogen peroxide-induced cytotoxicity in human neuroblastoma SH-SY5Y cells. BMC Complement. Altern. Med. 2015, 15, 171. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fang, J.; Li, S.; Gaur, U.; Xing, X.; Wang, H.; Zheng, W. Artemisinin Attenuated Hydrogen Peroxide (H2O2)-Induced Oxidative Injury in SH-SY5Y and Hippocampal Neurons via the Activation of AMPK Pathway. Int. J. Mol. Sci. 2019, 20, 2680. [Google Scholar] [CrossRef] [PubMed]
- Lieberknecht, V.; Engel, D.; Rodrigues, A.L.S.; Gabilan, N.H. Neuroprotective effects of mirtazapine and imipramine and their effect in pro- and anti-apoptotic gene expression in human neuroblastoma cells. Pharmacol. Rep. 2020, 72, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wang, H.; Li, Q.; Qian, Y.; Yao, W. Salidroside inhibits H2O2-induced apoptosis in PC 12 cells by preventing cytochrome c release and inactivating of caspase cascade. Acta Biochim. Biophys. Sin. 2008, 40, 796–802. [Google Scholar] [CrossRef]
- Wang, C.-M.; Yang, C.-Q.; Cheng, B.-H.; Chen, J.; Bai, B. Orexin-A protects SH-SY5Y cells against H2O2-induced oxidative damage via the PI3K/MEK 1/2 /ERK 1/2 signaling pathway. Int. J. Immunopathol. Pharmacol. 2018, 32, 205873841878573. [Google Scholar] [CrossRef] [Green Version]
- Correia, A.S.; Fraga, S.; Teixeira, J.P.; Vale, N. Cell Model of Depression: Reduction of Cell Stress with Mirtazapine. Int. J. Mol. Sci. 2022, 23, 4942. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.S.; Vale, N. Antidepressants in Alzheimer’s Disease: A Focus on the Role of Mirtazapine. Pharmaceuticals 2021, 14, 930. [Google Scholar] [CrossRef] [PubMed]
- Cowen, P.J.; Browning, M. What has serotonin to do with depression? World Psychiatry 2015, 14, 158–160. [Google Scholar] [CrossRef]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan metabolic pathways and brain serotonergic activity: A comparative review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zheng, J.; Huang, X.F.; Zhu, X.; Ding, S.M.; Ke, H.M.; O’Donnell, J.M.; Zhang, H.T.; Song, G.Q.; Xu, Y. The neuroprotective and antidepressant-like effects of Hcyb1, a novel selective PDE2 inhibitor. CNS Neurosci. Ther. 2018, 24, 652–660. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, W.; Feng, Y.; Liu, J.; Chen, H.; Wang, D.; Zhao, R. Melatonin alleviates hippocampal GR inhibition and depression-like behavior induced by constant light exposure in mice. Ecotoxicol. Environ. Saf. 2021, 228, 112979. [Google Scholar] [CrossRef]
- Xu, L.; Su, J.; Guo, L.; Wang, S.; Deng, X.; Ma, S. Modulation of LPA1 receptor-mediated neuronal apoptosis by Saikosaponin-d: A target involved in depression. Neuropharmacology 2019, 155, 150–161. [Google Scholar] [CrossRef]
- Fu, X.; Jiao, J.; Qin, T.; Yu, J.; Fu, Q.; Deng, X.; Ma, S.; Ma, Z. A New Perspective on Ameliorating Depression-Like Behaviors: Suppressing Neuroinflammation by Upregulating PGC-1α. Neurotox. Res. 2021, 39, 872–885. [Google Scholar] [CrossRef]
- Wu, X.; Luo, P.; Rao, W.; Dai, S.; Zhang, L.; Ma, W.; Pu, J.; Yu, Y.; Wang, J.; Fei, Z. Homer1a attenuates hydrogen peroxide-induced oxidative damage in HT-22 cells through AMPK-dependent autophagy. Front. Neurosci. 2018, 12, 51. [Google Scholar] [CrossRef]
- Gulec, M.; Oral, E.; Dursun, O.B.; Yucel, A.; Hacimuftuoglu, A.; Akcay, F.; Suleyman, H. Mirtazapine protects against cisplatin-induced oxidative stress and DNA damage in the rat brain. Psychiatry Clin. Neurosci. 2013, 67, 50–58. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, M.; Yu, H.; Yu, H.; Zhang, S.; Li, R. Mirtazapine prevents cell activation, inflammation, and oxidative stress against isoflurane exposure in microglia. Bioengineered 2021, 13, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Lv, M.; Yu, B.; He, J.; Zheng, P.; Yu, J.; Wang, Q.; Chen, D. The effect of dietary tryptophan levels on oxidative stress of liver induced by diquat in weaned piglets. J. Anim. Sci. Biotechnol. 2014, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.I.; Alves, A.G.; Carraro, L.R., Jr.; da Silva Teixeira Rech, T.; dos Santos Neto, J.S.; Alves, D.; Pereira Soares, M.S.; Spohr, L.; Spanevello, R.M.; Brüning, C.A.; et al. Insights into serotonergic and antioxidant mechanisms involved in antidepressant-like action of 2-phenyl-3-(phenylselanyl)benzofuran in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 102, 109956. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Hryb, A.B.; Platt, N.; Freitas, A.E.; Heinrich, I.A.; López, M.G.; Leal, R.B.; Kaster, M.P.; Rodrigues, A.L.S. Protective Effects of Ursolic Acid Against Cytotoxicity Induced by Corticosterone: Role of Protein Kinases. Neurochem. Res. 2019, 44, 2843–2855. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Wang, X.-Q.; Liu, M.-F.; Ye, B.-J. Meta-analysis of association between TPH2 single nucleotide poiymorphism and depression. Neurosci. Biobehav. Rev. 2022, 134, 104517. [Google Scholar] [CrossRef]
- Joëls, M.; Karst, H.; Sarabdjitsingh, R.A. The stressed brain of humans and rodents. Acta Physiol. 2018, 223, e13066. [Google Scholar] [CrossRef]
- HT-22 Mouse Hippocampal Neuronal Cell Line|SCC129. Available online: https://www.merckmillipore.com/PT/en/product/HT-22-Mouse-Hippocampal-Neuronal-Cell-Line,MM_NF-SCC129?ReferrerURL=https%3A%2F%2Fwww.google.com%2F (accessed on 31 May 2022).
- SH-SY5Y|ATCC. Available online: https://www.atcc.org/products/crl-2266 (accessed on 31 May 2022).
- BioRender. Available online: https://biorender.com/ (accessed on 1 June 2022).























Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, A.S.; Cardoso, A.; Vale, N. Significant Differences in the Reversal of Cellular Stress Induced by Hydrogen Peroxide and Corticosterone by the Application of Mirtazapine or L-Tryptophan. Int. J. Transl. Med. 2022, 2, 482-505. https://doi.org/10.3390/ijtm2030036
Correia AS, Cardoso A, Vale N. Significant Differences in the Reversal of Cellular Stress Induced by Hydrogen Peroxide and Corticosterone by the Application of Mirtazapine or L-Tryptophan. International Journal of Translational Medicine. 2022; 2(3):482-505. https://doi.org/10.3390/ijtm2030036
Chicago/Turabian StyleCorreia, Ana Salomé, Armando Cardoso, and Nuno Vale. 2022. "Significant Differences in the Reversal of Cellular Stress Induced by Hydrogen Peroxide and Corticosterone by the Application of Mirtazapine or L-Tryptophan" International Journal of Translational Medicine 2, no. 3: 482-505. https://doi.org/10.3390/ijtm2030036