
Detection of Neutralizing Antibodies in COVID-19 Patients from Steve Biko Academic Hospital Complex: A Pilot Study
 , , , , , ,
, , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Specimen Collection
2.2. RNA Extraction
2.3. SARS-CoV-2 Genotyping
2.4. COVID-19 IgG/IgM Rapid Test
2.5. SARS-CoV-2 Neutralizing Antibody Detection ELISA Assay
- [≥30%] = a positive result, indicating the presence of SARS-CoV-2 neutralizing antibodies;
- [<30%] = a negative result, indicating the absence of SARS-CoV-2 neutralizing antibodies, or that the detected level was below the limit of detection.
2.6. Statistical Analysis
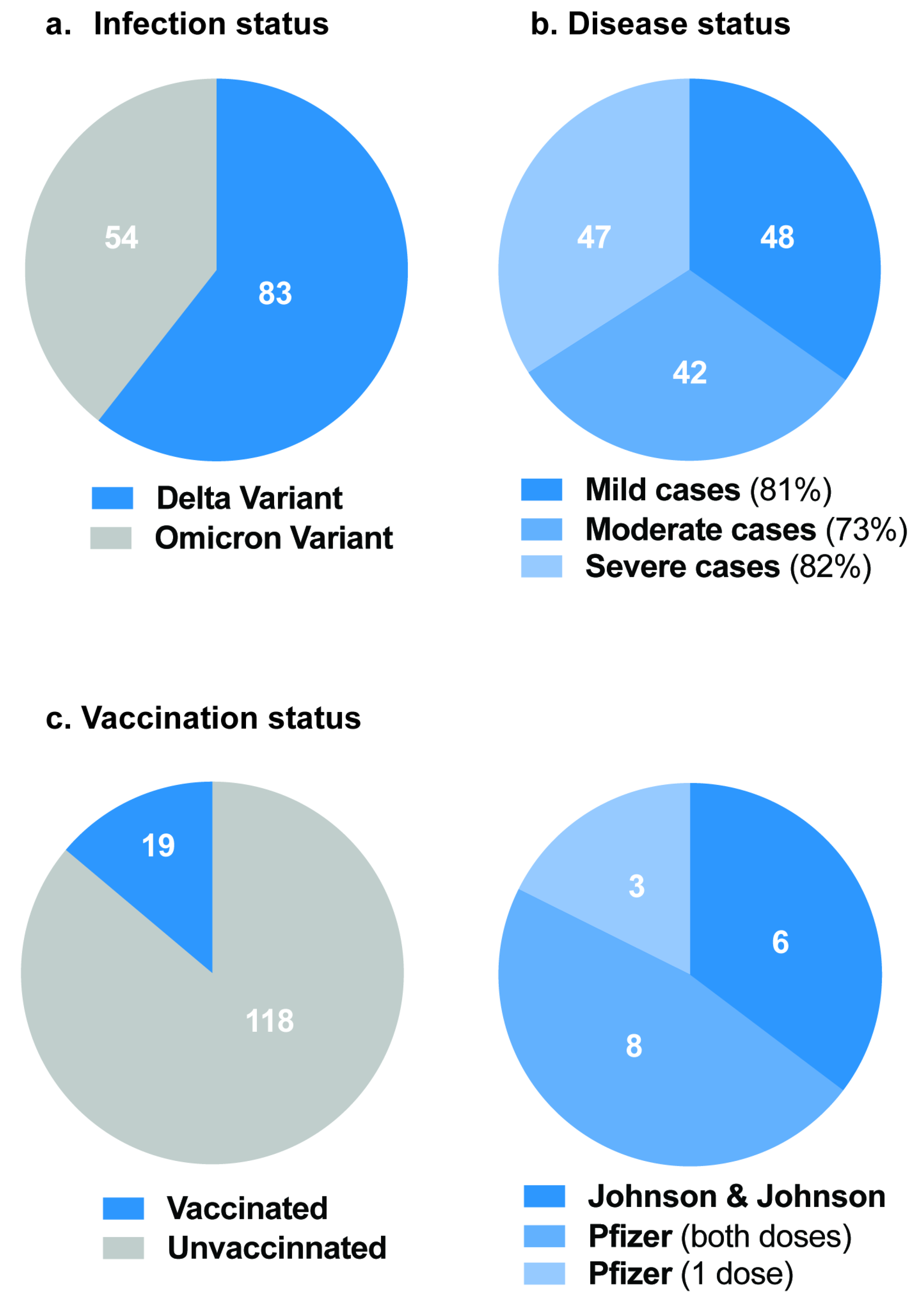
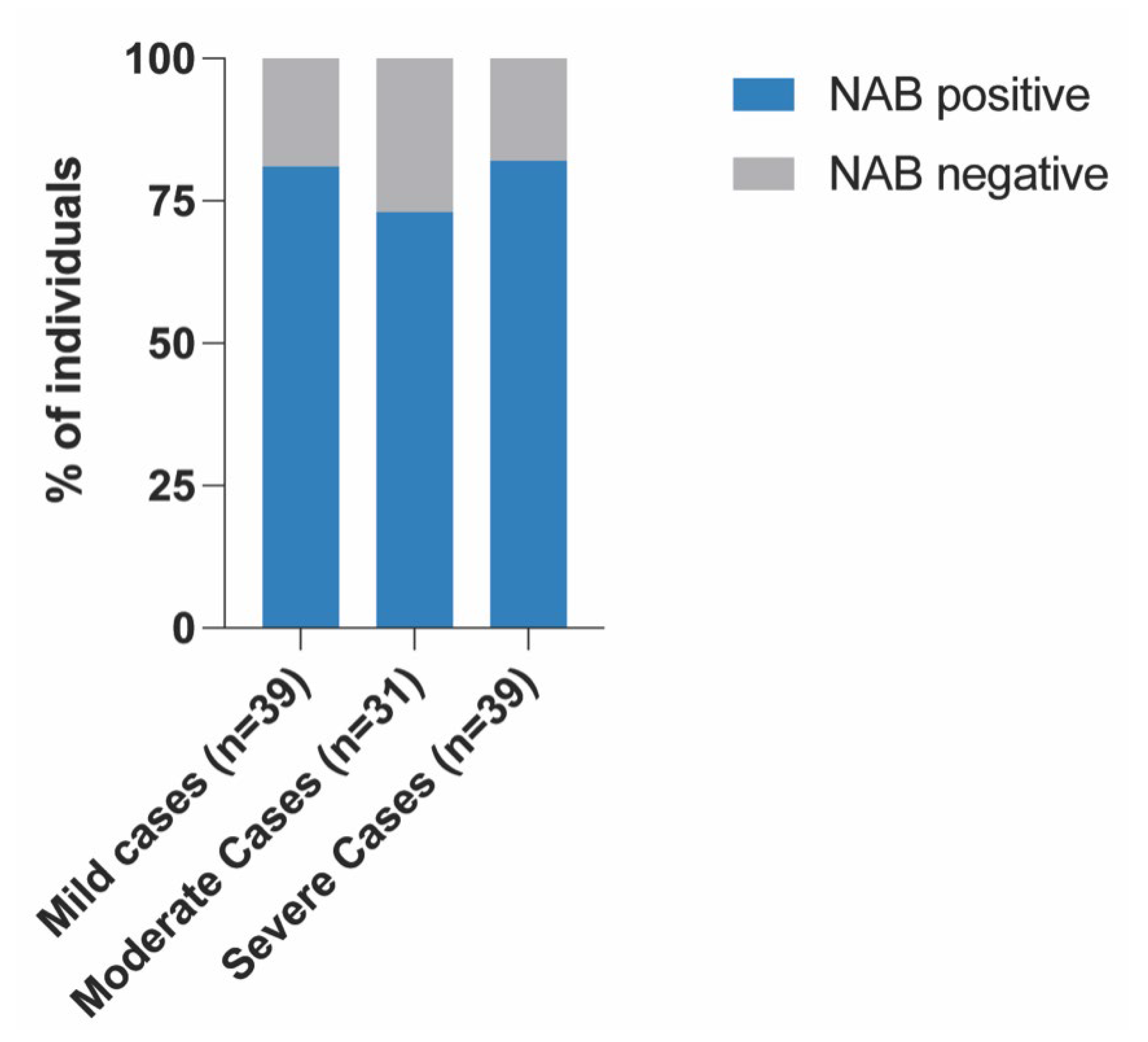
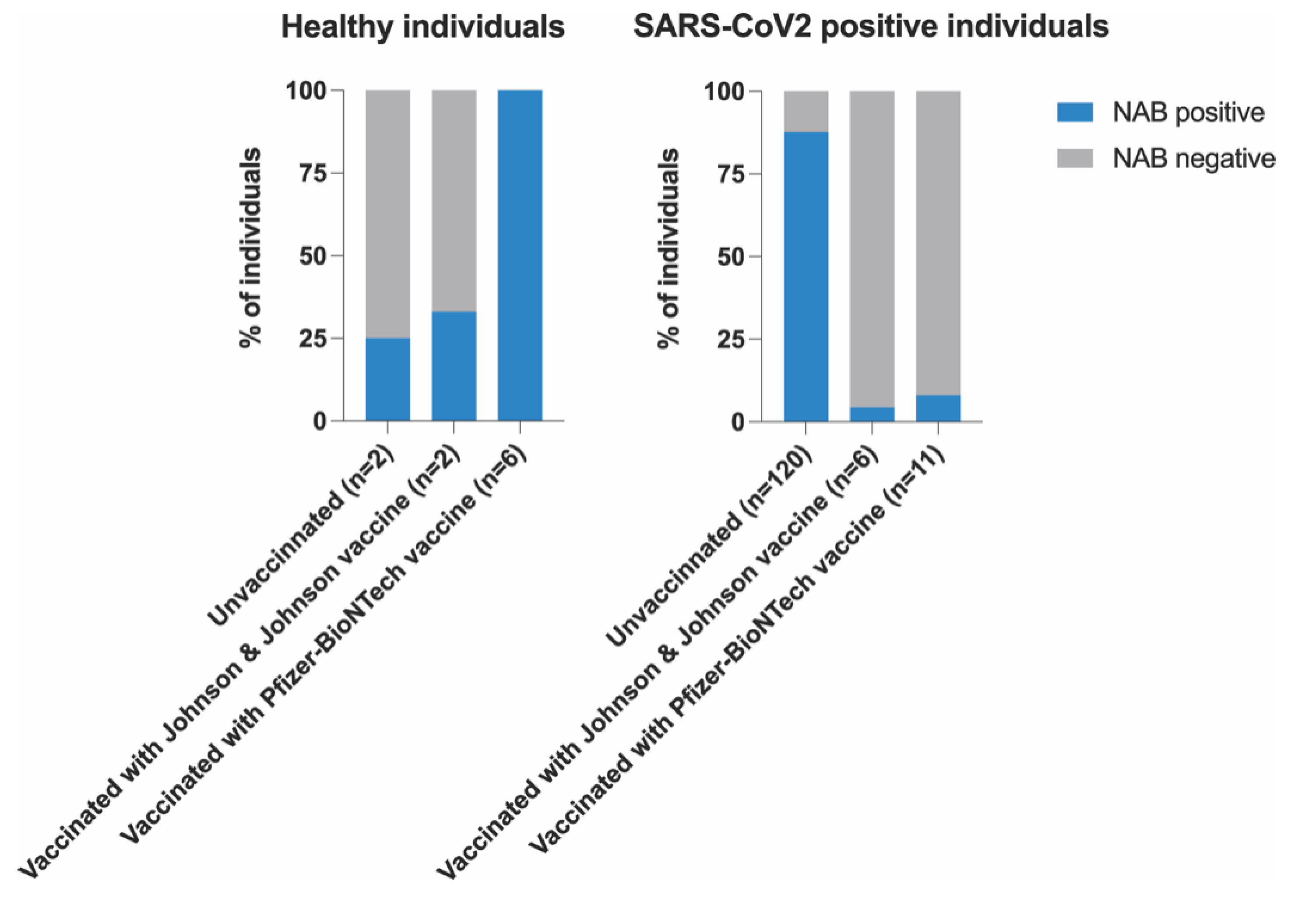
3. Results
Correlation between NABs, COVID-19 Variants and Healthy Participants
4. Discussion
5. Conclusions
6. Recommendations
7. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jebril, N. World Health Organization Declared a Pandemic Public Health Menace: A Systematic Review of the Coronavirus Disease 2019 “COVID-19”. 2020. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3566298 (accessed on 11 July 2022).
- Nehme, M.; Braillard, O.; Alcoba, G.; Perone, S.A.; Courvoisier, D.; Chappuis, F.; Guessous, I. COVID-19 Symptoms: Longitudinal Evolution and Persistence in Outpatient Settings. Ann. Intern. Med. 2021, 174, 723–725. [Google Scholar] [CrossRef]
- He, X.; Lau, E.H.Y.; Wu, P.; Deng, X.; Wang, J.; Hao, X.; Lau, Y.C.; Wong, J.Y.; Guan, Y.; Tan, X.; et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat. Med. 2020, 26, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.A. Mechanisms of Viral Neutralization. In Anonymous Viral Immunology and Immunopathology; Elsevier: Amsterdam, The Netherlands, 1975; pp. 79–97. [Google Scholar]
- Pang, N.Y.; Pang, A.S.; Chow, V.T.; Wang, D. Understanding neutralising antibodies against SARS-CoV-2 and their implications in clinical practice. Mil. Med. Res. 2021, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Sejdic, A.; Frische, A.; Jørgensen, C.S.; Rasmussen, L.D.; Trebbien, R.; Dungu, A.; Holler, J.G.; Ostrowski, S.R.; Eriksson, R.; Søborg, C.; et al. High titers of neutralizing SARS-CoV-2 antibodies six months after symptom onset are associated with increased severity in COVID-19 hospitalized patients. Virol. J. 2023, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef]
- Palacios-Pedrero, M.; Osterhaus, A.D.M.E.; Becker, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Aging and Options to Halt Declining Immunity to Virus Infections. Front. Immunol. 2021, 12, 681449. [Google Scholar] [CrossRef]
- Klein, S.L. Sex influences immune responses to viruses, and efficacy of prophylaxis and treatments for viral diseases. Bioessays 2012, 34, 1050–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chvatal-Medina, M.; Mendez-Cortina, Y.; Patiño, P.J.; Velilla, P.A.; Rugeles, M.T. Antibody Responses in COVID-19: A Review. Front. Immunol. 2021, 12, 633184. [Google Scholar] [CrossRef]
- Merad, M.; Blish, C.A.; Sallusto, F.; Iwasaki, A. The immunology and immunopathology of COVID-19. Science 2022, 375, 1122–1127. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Liu, P.; Li, X.; Zhou, S.; Li, H.; Wang, Z.; Shen, F.; Lu, B.; Long, Y.; Xiao, X.; et al. Neutralizing antibodies and cellular immune responses against SARS-CoV-2 sustained one and a half years after natural infection. Front. Microbiol. 2022, 12, 803031. [Google Scholar] [CrossRef]
- Nam, M.; Seo, J.D.; Moon, H. Evaluation of Humoral Immune Response after SARS-CoV-2 Vaccination Using Two Binding Antibody Assays and a Neutralizing Antibody Assay. ASM J. 2021, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Padoan, A.; Cosma, C.; Bonfante, F.; Della Rocca, F.; Barbaro, F.; Santarossa, C.; Dall’Olmo, L.; Pagliari, M.; Bortolami, A.; Cattelan, A.; et al. SARS-CoV-2 neutralizing antibodies after one or two doses of Comirnaty (BNT162b2, BioNTech/Pfizer): Kinetics and comparison with chemiluminescent assays. Clin. Chim. Acta 2021, 523, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Beltran, W.F.; Lam, E.C.; Astudillo, M.G.; Yang, D.; Miller, T.E.; Feldman, J.; Hauser, B.M.; Caradonna, T.M.; Clayton, K.L.; Nitido, A.D.; et al. COVID-19-neutralizing antibodies predict disease severity and survival. Cell 2020, 184, 476–488.e11. [Google Scholar] [CrossRef] [PubMed]
- Alejo, J.L.; Mitchell, J.; Chang, A.; Chiang, T.P.; Massie, A.B.; Segev, D.L.; Makary, M.A. Prevalence and durability of SARS-CoV-2 antibodies among unvaccinated US adults by history of COVID-19. JAMA 2022, 327, 1085–1087. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Choi, Y.J.; Kim, J.-M.; Kim, D.; Kim, I.-H.; Park, A.K.; Kim, H.M.; Rhee, J.E.; Jang, Y.R.; Yoo, C.K.; et al. Neutralizing antibody responses in vaccinated and unvaccinated individuals infected with Omicron BA.1 variant. J. Clin. Virol. 2022, 155, 105253. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Barouch, D.H. COVID-19 vaccines—Immunity, variants, boosters. N. Engl. J. Med. 2022, 387, 1011–1020. [Google Scholar] [CrossRef]
- Pradenas, E.; Trinité, B.; Urrea, V.; Marfil, S.; Ávila-Nieto, C.; de la Concepción, M.L.R.; Tarrés-Freixas, F.; Pérez-Yanes, F.; Rovirosa, C.; Ainsua-Enrich, E.; et al. Stable neutralizing an-tibody levels 6 months after mild and severe COVID-19 episodes. Med 2021, 2, 313–320.e4. [Google Scholar] [CrossRef]
- Tada, T.; Zhou, H.; Samanovic, M.I.; Dcosta, B.M.; Cornelius, A.; Mulligan, M.J.; Landau, N.R. Comparison of neutralizing antibody titers elicited by mRNA and adenoviral vector vaccine against SARS-CoV-2 variants. BioRxiv 2021. [Google Scholar] [CrossRef]
- Zahid, M.N.; Moosa, M.S.; Perna, S.; Buti, E.B. A review on COVID-19 vaccines: Stages of clinical trials, mode of actions and efficacy. Arab J. Basic Appl. Sci. 2021, 28, 225–233. [Google Scholar] [CrossRef]
- Ng, K.W.; Faulkner, N.; Cornish, G.H.; Rosa, A.; Harvey, R.; Hussain, S.; Ulferts, R.; Earl, C.; Wrobel, A.G.; Benton, D.J.; et al. Preexisting and de novo humoral immunity to SARS-CoV-2 in humans. Science 2020, 370, 1339–1343. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.W.; Xie, Y.; Knuth, K.B.; Mack, C.D.; Brinkley, E.; Toovey, S.; Dreyer, N.A. COVID-19 vaccination breakthrough infections in a real-world setting: Using community reporters to evaluate vaccine effectiveness. Infect. Drug Resist. 2022, 15, 5167–5182. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.B.; Aggarwal, R.; Jani, I.; Jawad, J.; Kochhar, S.; MacDonald, N.; Madhi, S.A.; Mohsni, E.; Mulholland, K.; Neuzil, K.M.; et al. COVID-19 vaccine strategies must focus on severe disease and global equity. Lancet 2021, 399, 406–410. [Google Scholar] [CrossRef]
- Madhi, S.A.; Kwatra, G.; Myers, J.E.; Jassat, W.; Dhar, N.; Mukendi, C.K.; Nana, A.J.; Blumberg, L.; Welch, R.; Ngorima-Mabhena, N.; et al. Population Immunity and COVID-19 Severity with Omicron Variant in South Africa. N. Engl. J. Med. 2022, 386, 1314–1326. [Google Scholar] [CrossRef]
- Hasan, D.; Shono, A.; van Kalken, C.K.; van der Spek, P.J.; Krenning, E.P.; Kotani, T. A novel definition and treatment of hyperinflammation in COVID-19 based on purinergic signalling. Purinergic Signal. 2021, 18, 13–59. [Google Scholar] [CrossRef]
- Gustine, J.N.; Jones, D. Immunopathology of Hyperinflammation in COVID-19. Am. J. Pathol. 2020, 191, 4–17. [Google Scholar] [CrossRef]
- Bonnet, B.; Cosme, J.; Dupuis, C.; Coupez, E.; Adda, M.; Calvet, L.; Fabre, L.; Saint-Sardos, P.; Bereiziat, M.; Vidal, M.; et al. Severe COVID-19 is characterized by the co-occurrence of moderate cytokine inflammation and severe monocyte dysregulation. Ebiomedicine 2021, 73, 103622. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Dadras, O.; Afsahi, A.M.; Pashaei, Z.; Mojdeganlou, H.; Karimi, A.; Habibi, P.; Barzegary, A.; Fakhfouri, A.; Mirzapour, P.; Janfaza, N.; et al. The relationship between COVID-19 viral load and disease severity: A systematic review. Immun. Inflamm. Dis. 2021, 10, e580. [Google Scholar] [CrossRef] [PubMed]
- Knudtzen, F.C.; Jensen, T.G.; Lindvig, S.O.; Rasmussen, L.D.; Madsen, L.W.; Hoegh, S.V.; Bek-Thomsen, M.; Laursen, C.B.; Nielsen, S.L.; Johansen, I.S. SARS-CoV-2 viral load as a predictor for disease severity in outpatients and hospitalised patients with COVID-19: A prospective cohort study. PLoS ONE 2021, 16, e0258421. [Google Scholar] [CrossRef] [PubMed]
- Morales-Núñez, J.J.; Muñoz-Valle, J.F.; Torres-Hernández, P.C.; Hernández-Bello, J. Overview of Neutralizing Antibodies and Their Potential in COVID-19. Vaccines 2021, 9, 1376. [Google Scholar] [CrossRef] [PubMed]
- Arankalle, V.; Kulkarni-Munje, A.; Kulkarni, R.; Palkar, S.; Patil, R.; Oswal, J.; Lalwani, S.; Mishra, A.C. Immunogenicity of two COVID-19 vaccines used in India: An observational cohort study in health care workers from a tertiary care hospital. Front. Immunol. 2022, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Acharya, C.B.; Schrom, J.; Mitchell, A.M.; Coil, D.A.; Marquez, C.; Rojas, S.; Wang, C.Y.; Liu, J.; Pilarowski, G.; Solis, L.; et al. Viral Load Among Vaccinated and Unvaccinated, Asymptomatic and Symptomatic Persons Infected With the SARS-CoV-2 Delta Variant. Open Forum Infect. Dis. 2022, 9, ofac135. [Google Scholar] [CrossRef]
- Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Jacquérioz, F.; Kaiser, L.; Vetter, P.; Eckerle, I.; et al. Infectious viral load in unvaccinated and vaccinated individuals infected with ancestral, Delta or Omicron SARS-CoV-2. Nat. Med. 2022, 28, 1491–1500. [Google Scholar] [CrossRef]
- Mohammed, H.; Pham-Tran, D.D.; Yeoh, Z.Y.M.; Wang, B.; McMillan, M.; Andraweera, P.H.; Marshall, H.S. A Systematic Review and Meta-Analysis on the Real-World Effectiveness of COVID-19 Vaccines against Infection, Symptomatic and Severe COVID-19 Disease Caused by the Omicron Variant (B.1.1.529). Vaccines 2023, 11, 224. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mild | Moderate | Severe | r-Value | p-Value | |
|---|---|---|---|---|---|
| No.(%) | No.(%) | No.(%) | |||
| Age (years) | −0.01694 | 0.84424 | |||
| Total | 48 | 42 | 47 | ||
| <40 | 10 (20.8) | 7(16.7) | 12(25.5) | ||
| 40–69 | 37(77) | 26(61.9) | 30(63.8) | ||
| ≥70 | 1(2) | 9(2.1) | 5(10.6) | ||
| Gender | −0.11057 | 0.19836 | |||
| Male | 28(58.3) | 19(45.2) | 21(44.7) | ||
| Female | 20(41.7) | 23(54.8) | 26(55.3) | ||
| Smoker | 0.17099 | 0.04574 | |||
| Yes | 11(22.9) | 7(16.7) | 44(93.6) | ||
| No | 34(70.8) | 32(76.2) | 3(6.4) | ||
| Ex-smoker | 3(6.3) | 3(7.1) | 0(0) | ||
| Variants | |||||
| Beta | −0.00567 | 0.94761 | |||
| Delta | 27(56.3) | 22(52.4) | 34(72.3) | ||
| Omicron | 21(43.8) | 20(47.6) | 13(27.7) | ||
| Vaccination | 0.08305 | 0.33465 | |||
| Yes | 6(12.5) | 9(21.4) | 4(8.5) | ||
| No | 42(87.5) | 33(78.6) | 43(91.5) | ||
| Co-morbidities | |||||
| HIV | −0.05431 | 0.52849 | |||
| Yes | 9(18.8) | 6(14.3) | 6(12.8) | ||
| No | 39(81.3) | 36(85.7) | 41(87.2) | ||
| Hypertension | 0.11351 | 0.18659 | |||
| Yes | 19(39.6) | 18(42.9) | 26(55.3) | ||
| No | 29(60.4) | 24(57.1) | 21(44.7) | ||
| Diabetes | −0.03568 | 0.67892 | |||
| Yes | 7(14.6) | 9(21.4) | 18(38.3) | ||
| No | 41(85.4) | 33(78.6) | 29(61.7) | ||
| NAB Production | r | p |
|---|---|---|
| Age | −0.01694 | 0.84424 |
| Gender | 0.05021 | 0.56013 |
| VOCs | −0.00567 | 0.94761 |
| Mild | −0.01061 | 0.94292 |
| Moderate | 0.1574 | 0.31948 |
| Severe | −0.31394 | 0.03164 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kgatle, M.; Chalwe, J.M.; van der Westhuizen, D.; Xu, S.; Damane, B.P.; Mathebela, P.; Ueckermann, V.; Mayaphi, S.; Rodrigues, H.G.; Moura-Alves, P.; et al. Detection of Neutralizing Antibodies in COVID-19 Patients from Steve Biko Academic Hospital Complex: A Pilot Study. COVID 2023, 3, 999-1010. https://doi.org/10.3390/covid3070072
Kgatle M, Chalwe JM, van der Westhuizen D, Xu S, Damane BP, Mathebela P, Ueckermann V, Mayaphi S, Rodrigues HG, Moura-Alves P, et al. Detection of Neutralizing Antibodies in COVID-19 Patients from Steve Biko Academic Hospital Complex: A Pilot Study. COVID. 2023; 3(7):999-1010. https://doi.org/10.3390/covid3070072
Chicago/Turabian StyleKgatle, Mankgopo, Joseph Musonda Chalwe, Donald van der Westhuizen, Shuting Xu, Botle Precious Damane, Precious Mathebela, Veronica Ueckermann, Simnikiwe Mayaphi, Hosana Gomes Rodrigues, Pedro Moura-Alves, and et al. 2023. "Detection of Neutralizing Antibodies in COVID-19 Patients from Steve Biko Academic Hospital Complex: A Pilot Study" COVID 3, no. 7: 999-1010. https://doi.org/10.3390/covid3070072