
Structural Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Probiotic Leuconostoc mesenteroides 201607 Isolated from Fermented Food

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Partial Purification of EPS (PP-LMEPS)
2.2. Exopolysaccharide Characterization
2.2.1. Determination of the Monosaccharide Composition
2.2.2. Structural Characterization of (PP-LMEPS)
Fourier Transform Infrared Spectroscopy (FTIR) Analysis
Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
2.3. Functional Properties of PP-LMEPS
2.3.1. Antioxidant and Antimicrobial Assay of PP-LMEPS
2.3.2. Immunomodulatory Activity of PP-LMEPS
Cell Culture
Cells Viability Assay
IL-6 Assay
2.4. Statistical Analysis
3. Results and Discussion
3.1. LMEPS Production and Its Characterization
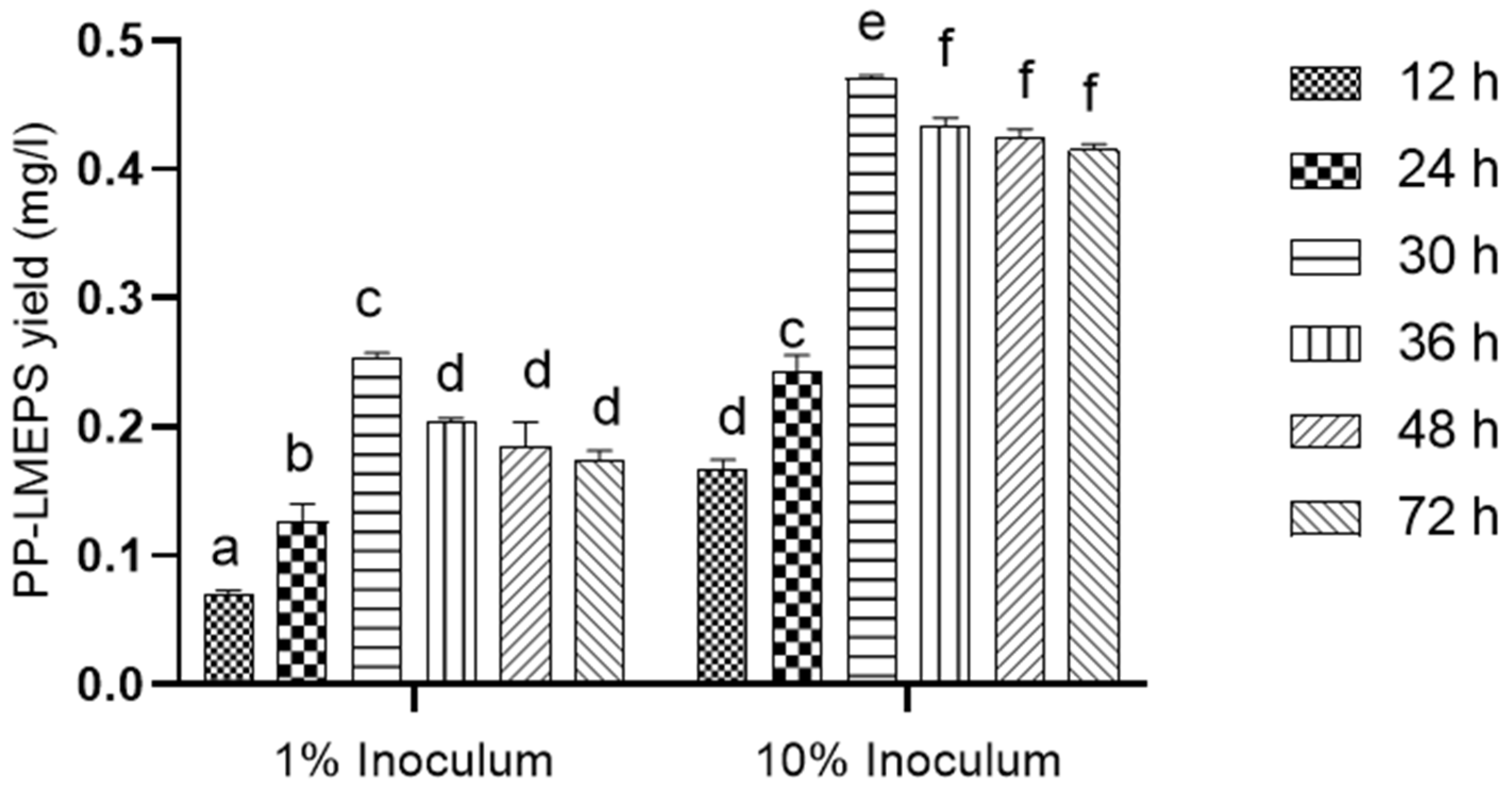
3.1.1. Partial Purification of EPS (PP-LMEPS)
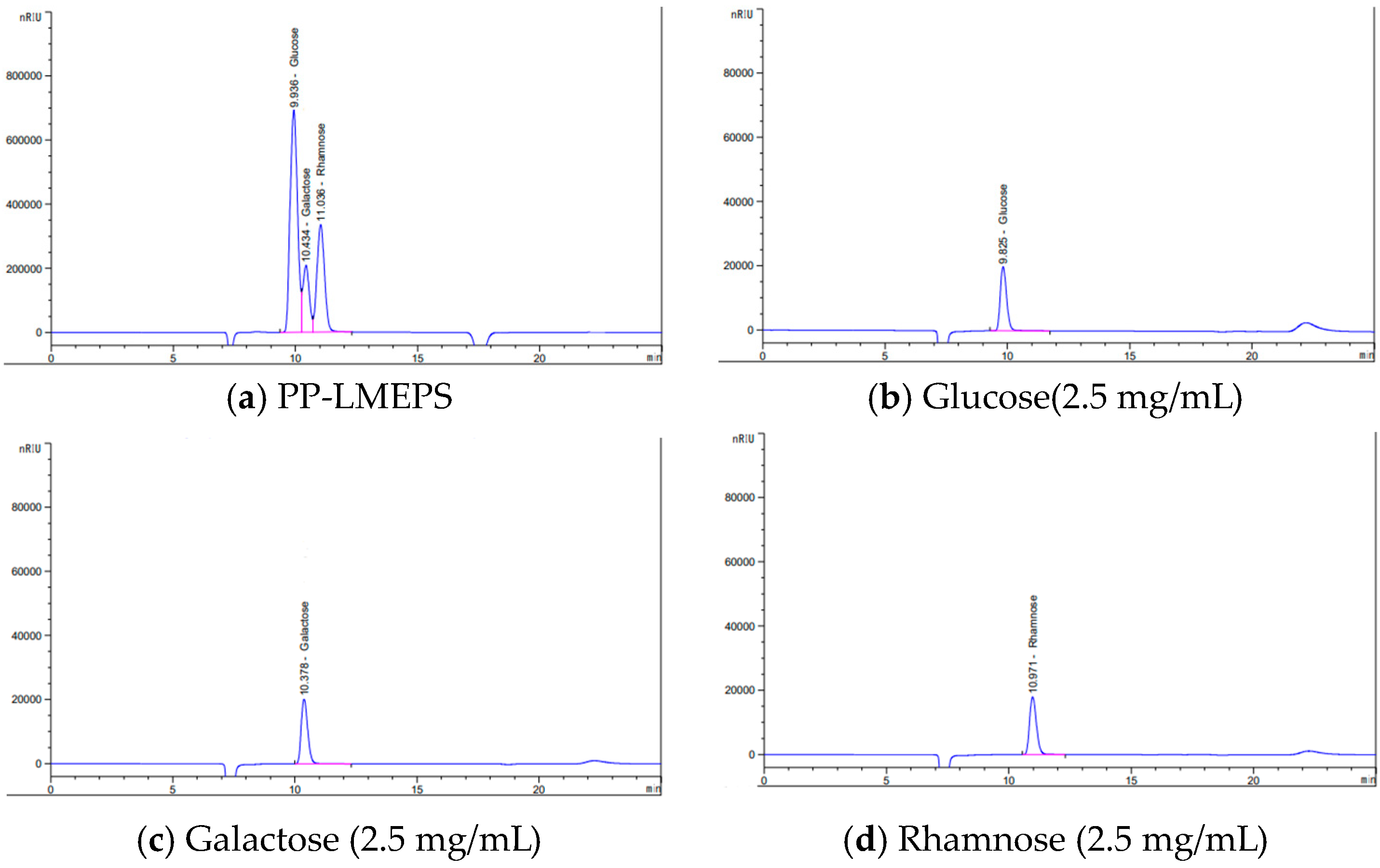
3.1.2. Monosaccharide Composition Analysis
3.1.3. Structural Characterization
FT-IR Spectrum Analysis
NMR Spectral Analysis
3.2. Functional Properties of PP-LMEPS
3.2.1. In vitro Antioxidant Analysis
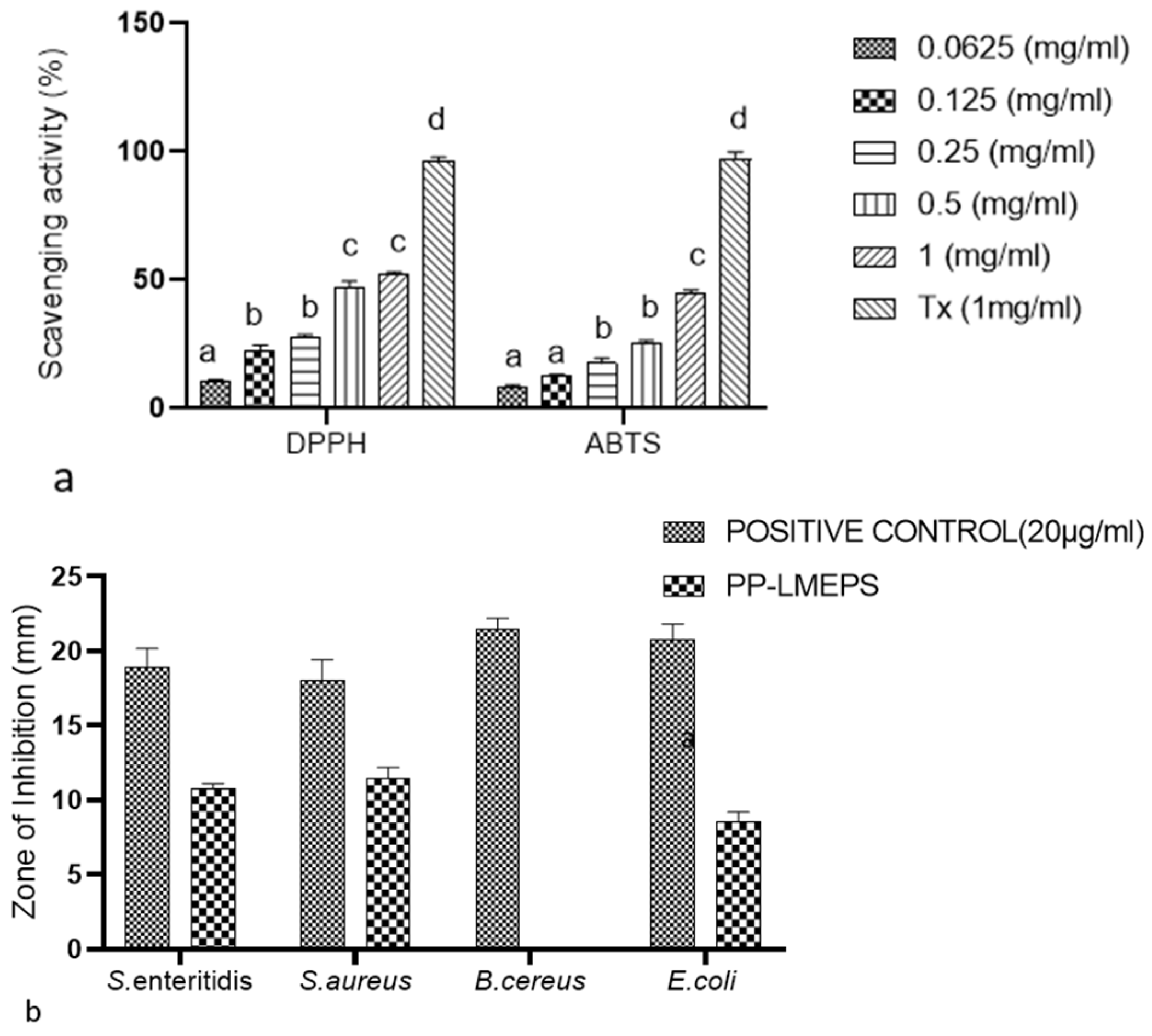
DPPH Radical Scavenging Activity
ABTS Radical Scavenging Activity
3.2.2. Antimicrobial Activity of PP-LMEPS
3.2.3. Effect of PP-LMEPS on Cell Viability against RAW 264.7
3.2.4. Effect of PP-LMEPS on Immunomodulatory Cytokine Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Badel, S.; Bernardi, T.; Michaud, P. New perspectives for Lactobacilli exopolysaccharides. Biotechnol. Adv. 2011, 29, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.; Cao, C.; Zhu, X.; Wang, C.; Wu, R.; Wu, J. Extraction and biological activity of exopolysaccharide produced by Leuconostoc mesenteroides SN-8. Int. J. Biol. Macromol. 2020, 157, 36–44. [Google Scholar] [CrossRef]
- Taylan, O.; Yilmaz, M.T.; Dertli, E. Partial characterization of a levan type exopolysaccharide (EPS) produced by Leuconostoc mesenteroides showing immunostimulatory and antioxidant activities. Int. J. Biol. Macromol. 2019, 136, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Han, Y.; Zhou, Z. In vitro prebiotic activities of exopolysaccharide from Leuconostoc pseudomesenteroides XG5 and its effect on the gut microbiota of mice. J. Funct. Foods 2020, 67, 103853. [Google Scholar] [CrossRef]
- Saha, I.; Datta, S. Bacterial exopolysaccharides in drug delivery applications. J. Drug Deliv. Sci. Technol. 2022, 74, 103557. [Google Scholar] [CrossRef]
- Miyamoto, J.; Shimizu, H.; Hisa, K.; Matsuzaki, C.; Inuki, S.; Ando, Y.; Nishida, A.; Izumi, A.; Yamano, M.; Ushiroda, C.; et al. Host metabolic benefits of prebiotic exopolysaccharides produced by Leuconostoc mesenteroides. Gut Microbes 2023, 15, 2161271. [Google Scholar] [CrossRef]
- Matsuzaki, C.; Hayakawa, A.; Matsumoto, K.; Katoh, T.; Yamamoto, K.; Hisa, K. Exopolysaccharides produced by Leuconostoc mesenteroides strain NTM048 as an immunostimulant to enhance the mucosal barrier and influence the systemic immune response. J. Agric. Food Chem. 2015, 63, 7009–7015. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef]
- Kook, S.-Y.; Lee, Y.; Jeong, E.-C.; Kim, S. Immunomodulatory effects of exopolysaccharides produced by Bacillus licheniformis and Leuconostoc mesenteroides isolated from Korean kimchi. J. Funct. Foods 2019, 54, 211–219. [Google Scholar] [CrossRef]
- Amrutha, T.; Beena, A. Microbial Exopolysaccharides: A Promising Health Booster. J. Phytopharmacol. 2023, 12, 265–271. [Google Scholar] [CrossRef]
- Matsuzaki, C.; Nakashima, Y.; Endo, I.; Tomabechi, Y.; Higashimura, Y.; Itonori, S.; Hosomi, K.; Kunisawa, J.; Yamamoto, K.; Hisa, K. Enzymatically synthesized exopolysaccharide of a probiotic strain Leuconostoc mesenteroides NTM048 shows adjuvant activity to promote IgA antibody responses. Gut Microbes 2021, 13, 1949097. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Jiao, Z.; Wang, T.; Zhang, B.; Zhao, H. Glucans with Different Degrees of Polymerization from Leuconostoc mesenteroides CICC6055: Analysis of Physicochemical Properties and Intestinal Prebiotic Function. Int. J. Mol. Sci. 2023, 25, 258. [Google Scholar] [CrossRef] [PubMed]
- Bisson, G.; Comuzzi, C.; Giordani, E.; Poletti, D.; Boaro, M.; Marino, M. An exopolysaccharide from Leuconostoc mesenteroides showing interesting bioactivities versus foodborne microbial targets. Carbohydr. Polym. 2023, 301, 120363. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gu, Y.; Zheng, X.; Zhang, Y.; Deng, K.; Wu, T.; Cheng, H. Analysis of physicochemical properties of exopolysaccharide from Leuconostoc mesenteroides strain XR1 and its application in fermented milk. LWT 2021, 146, 111449. [Google Scholar] [CrossRef]
- Choi, I.S.; Ko, S.H.; Lee, M.E.; Kim, H.M.; Yang, J.E.; Jeong, S.G.; Lee, K.H.; Chang, J.Y.; Kim, J.-C.; Park, H.W. Production, characterization, and antioxidant activities of an exopolysaccharide extracted from spent media wastewater after Leuconostoc mesenteroides WiKim32 fermentation. ACS Omega 2021, 6, 8171–8178. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Ha, J.-H.; Lee, J.; Jang, H.; Kwon, D.; Cho, M.; Kang, D.; Kim, I.; Kim, M. The Effect of Fermented Momordica charantia with Leuconostoc mesenteroides MKSR on Metabolic Complications Induced by High-Fat High-Cholesterol Diet in C57BL/6 Mice. Fermentation 2023, 9, 718. [Google Scholar] [CrossRef]
- Farinazzo, F.S.; Valente, L.J.; Almeida, M.B.; Simionato, A.S.; Fernandes, M.T.C.; Mauro, C.S.I.; Tomal, A.A.B.; Garcia, S. Characterization and antioxidant activity of an exopolysaccharide produced by Leuconostoc pseudomesenteroides JF17 from juçara fruits (Euterpe edulis Martius). Process. Biochem. 2019, 91, 141–148. [Google Scholar] [CrossRef]
- Wang, L.; Gu, Y.; Lv, Z. Processing properties of yogurt as affected by the EPS produced by Leuconostoc mesenteroides XR1. Int. J. Food Sci. Technol. 2022, 57, 4076–4085. [Google Scholar] [CrossRef]
- Wu, J.; Yan, D.; Liu, Y.; Luo, X.; Li, Y.; Cao, C.; Li, M.; Han, Q.; Wang, C.; Wu, R.; et al. Purification, structural characteristics, and biological activities of exopolysaccharide isolated from Leuconostoc mesenteroides SN-8. Front. Microbiol. 2021, 12, 644226. [Google Scholar] [CrossRef]
- Jiang, J.; Guo, S.; Ping, W.; Zhao, D.; Ge, J. Optimization production of exopolysaccharide from Leuconostoc lactis L2 and its partial characterization. Int. J. Biol. Macromol. 2020, 159, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, C.; Takagaki, C.; Tomabechi, Y.; Forsberg, L.S.; Heiss, C.; Azadi, P.; Matsumoto, K.; Katoh, T.; Hosomi, K.; Kunisawa, J.; et al. Structural characterization of the immunostimulatory exopolysaccharide produced by Leuconostoc mesenteroides strain NTM048. Carbohydr. Res. 2017, 448, 95–102. [Google Scholar] [CrossRef]
- Ayyash, M.; Abu-Jdayil, B.; Olaimat, A.; Esposito, G.; Itsaranuwat, P.; Osaili, T.; Obaid, R.; Kizhakkayil, J.; Liu, S.-Q. Physicochemical, bioactive and rheological properties of an exopolysaccharide produced by a probiotic Pediococcus pentosaceus M41. Carbohydr. Polym. 2020, 229, 115462. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Zou, S.; Liang, D.; Luan, L. Structural characterization, antioxidant and hepatoprotective activities of polysaccharides from Sophorae tonkinensis Radix. Carbohydr. Polym. 2018, 184, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Z.; Qian, J.; Zhou, D. Analysis on the Changes of Functional Groups after Coal Dust Explosion at Different Concentrations Based on FTIR and XRD. Combust. Sci. Technol. 2020, 193, 2482–2504. [Google Scholar] [CrossRef]
- Vijayalakshmi, S.; Yoo, D.-S.; Kim, D.-G.; Chelliah, R.; Barathikannan, K.; Aloo, S.-O.; Tyagi, A.; Yan, P.; Shan, L.; Gebre, T.S.; et al. Fermented Perilla frutescens leaves and their untargeted metabolomics by UHPLC-QTOF-MS reveal anticancer and immunomodulatory effects. Food Biosci. 2023, 56, 103065. [Google Scholar] [CrossRef]
- Vijayalakshmi, S.; Adeyemi, D.E.; Choi, I.Y.; Sultan, G.; Madar, I.H.; Park, M.-K. Comprehensive in silico analysis of lactic acid bacteria for the selection of desirable probiotics. LWT 2020, 130, 109617. [Google Scholar] [CrossRef]
- Khan, R.; Shah, M.D.; Shah, L.; Lee, P.-C.; Khan, I. Bacterial polysaccharides—A big source for prebiotics and therapeutics. Front. Nutr. 2022, 9, 1031935. [Google Scholar] [CrossRef]
- Madhuri, K.V.; Prabhakar, K.V. Microbial Exopolysaccharides: Biosynthesis and Potential Applications. Orient. J. Chem. 2014, 30, 1401–1410. [Google Scholar] [CrossRef]
- De Vuyst, L.; Degeest, B. Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev. 1999, 23, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Dilna, S.V.; Surya, H.; Aswathy, R.G.; Varsha, K.K.; Sakthikumar, D.N.; Pandey, A.; Nampoothiri, K.M. Characterization of an exopolysaccharide with potential health-benefit properties from a probiotic Lactobacillus plantarum RJF4. LWT 2015, 64, 1179–1186. [Google Scholar] [CrossRef]
- You, X.; Li, Z.; Ma, K.; Zhang, C.; Chen, X.; Wang, G.; Yang, L.; Dong, M.; Rui, X.; Zhang, Q.; et al. Structural characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus helveticus LZ-R-5. Carbohydr. Polym. 2020, 235, 115977. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Saravanakumar, K.; Sathiyaseelan, A.; Park, S.; Hu, X.; Wang, M.-H. Cellular antioxidant properties of nontoxic exopolysaccharide extracted from Lactobacillales (Weissella cibaria) isolated from Korean kimchi. LWT 2022, 154, 112727. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Babinastarlin, S.; Shankar, T.; Sivakumar, T.; Anandapandian, K.T.K. Quantification and characterization of exopolysaccharides from Bacillus subtilis (MTCC 121). Adv. Biol. Res. 2011, 5, 71–76. [Google Scholar]
- Ai, L.; Guo, Q.; Ding, H.; Guo, B.; Chen, W.; Cui, S.W. Structure characterization of exopolysaccharides from Lactobacillus casei LC2W from skim milk. Food Hydrocoll. 2016, 56, 134–143. [Google Scholar] [CrossRef]
- Perera, K.Y.; Jaiswal, A.K.; Jaiswal, S. Biopolymer-Based Sustainable Food Packaging Materials: Challenges, Solutions, and Applications. Foods 2023, 12, 2422. [Google Scholar] [CrossRef] [PubMed]
- Rajoka, M.S.R.; Mehwish, H.M.; Fang, H.; Padhiar, A.A.; Zeng, X.; Khurshid, M.; He, Z.; Zhao, L. Characterization and anti-tumor activity of exopolysaccharide produced by Lactobacillus kefiri isolated from Chinese kefir grains. J. Funct. Foods 2019, 63, 103588. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Tian, Z.; Yang, Y.; Yang, Z. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir. Carbohydr. Polym. 2015, 125, 16–25. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef]
- Ye, G.; Chen, Y.; Wang, C.; Yang, R.; Bin, X. Purification and characterization of exopolysaccharide produced by Weissella cibaria YB-1 from pickle Chinese cabbage. Int. J. Biol. Macromol. 2018, 120, 1315–1321. [Google Scholar] [CrossRef]
- Jeong, D.; Kim, D.-H.; Kang, I.-B.; Kim, H.; Song, K.-Y.; Kim, H.-S.; Seo, K.-H. Characterization and antibacterial activity of a novel exopolysaccharide produced by Lactobacillus kefiranofaciens DN1 isolated from kefir. Food Control 2017, 78, 436–442. [Google Scholar] [CrossRef]
- Abdalla, A.K.; Ayyash, M.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Shah, N.P.; Holley, R. Exopolysaccharides as Antimicrobial Agents: Mechanism and Spectrum of Activity. Front. Microbiol. 2021, 12, 664395. [Google Scholar] [CrossRef]
- Amini, E.; Salimi, F.; Imanparast, S.; Mansour, F. Isolation and characterization of exopolysaccharide derived from Lacticaseibacillus paracasei AS20(1) with probiotic potential and evaluation of its antibacterial activity. Lett. Appl. Microbiol. 2022, 75, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Nam, J.H.; Rod-In, W.; Monmai, C.; Jang, A.-Y.; You, S.; Park, W.J. Korean Ginseng Berry Polysaccharide Enhances Immunomodulation Activities of Peritoneal Macrophages in Mice with Cyclophosphamide-Induced Immunosuppression. J. Microbiol. Biotechnol. 2023, 33, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Wang, S.L. Recent advances in exopolysaccharides from Paenibacillus spp.: Production, isolation, structure, and bioactivities. Mar. Drugs 2015, 13, 1847–1863. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, F. Polysaccharides: Candidates of promising vaccine adjuvants. Drug Discov. Ther. 2015, 9, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-G.; Lee, H.J.; Cho, J.-Y.; Kim, K.; Yang, S.J.; Kim, D. Anti-inflammatory effects of sucrose-derived oligosaccharides produced by a constitutive mutant L. mesenteroides B-512FMCM dextransucrase in high fat diet-fed mice. Biochem. Biophys. Res. Commun. 2016, 477, 350–355. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory Effects Triggered by Lactic Acid Bacteria Exopolysaccharides: New Insights into Molecular Interactions with Host Cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijayalakshmi, S.; Kim, J.-R.; Chelliah, R.; Barathikannan, K.; Hirad, A.H.; Oh, D.-H. Structural Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Probiotic Leuconostoc mesenteroides 201607 Isolated from Fermented Food. Appl. Microbiol. 2024, 4, 329-340. https://doi.org/10.3390/applmicrobiol4010022
Vijayalakshmi S, Kim J-R, Chelliah R, Barathikannan K, Hirad AH, Oh D-H. Structural Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Probiotic Leuconostoc mesenteroides 201607 Isolated from Fermented Food. Applied Microbiology. 2024; 4(1):329-340. https://doi.org/10.3390/applmicrobiol4010022
Chicago/Turabian StyleVijayalakshmi, Selvakumar, Jong-Rai Kim, Ramachandran Chelliah, Kaliyan Barathikannan, Abdurahman Hajinur Hirad, and Deog-Hwan Oh. 2024. "Structural Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Probiotic Leuconostoc mesenteroides 201607 Isolated from Fermented Food" Applied Microbiology 4, no. 1: 329-340. https://doi.org/10.3390/applmicrobiol4010022