

Sublethal Exposure to Common Benzalkonium Chloride Leads to Antimicrobial Tolerance and Antibiotic Cross-Resistance in Commensal and Opportunistic Bacterial Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Species and Growth Conditions
2.2. BAC12–14 and Purification and Identification of BAC16
2.3. Determination of BAC Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC)
2.4. Bacterial Acclimation to BAC Compounds
2.5. Determination of the Contribution of Efflux Pump Activity to Antimicrobial Tolerance
2.6. Determination of Antibiotic Susceptibilities
2.7. Statistical Analyses
3. Results
3.1. BAC Compounds and Purification of BAC16
3.2. Bacterial Sensitivities to BAC-Inhibition
3.3. Repeated Exposure to Sublethal BAC Concentrations Leads to Antimicrobial Tolerance in Opportunistic Pathogens
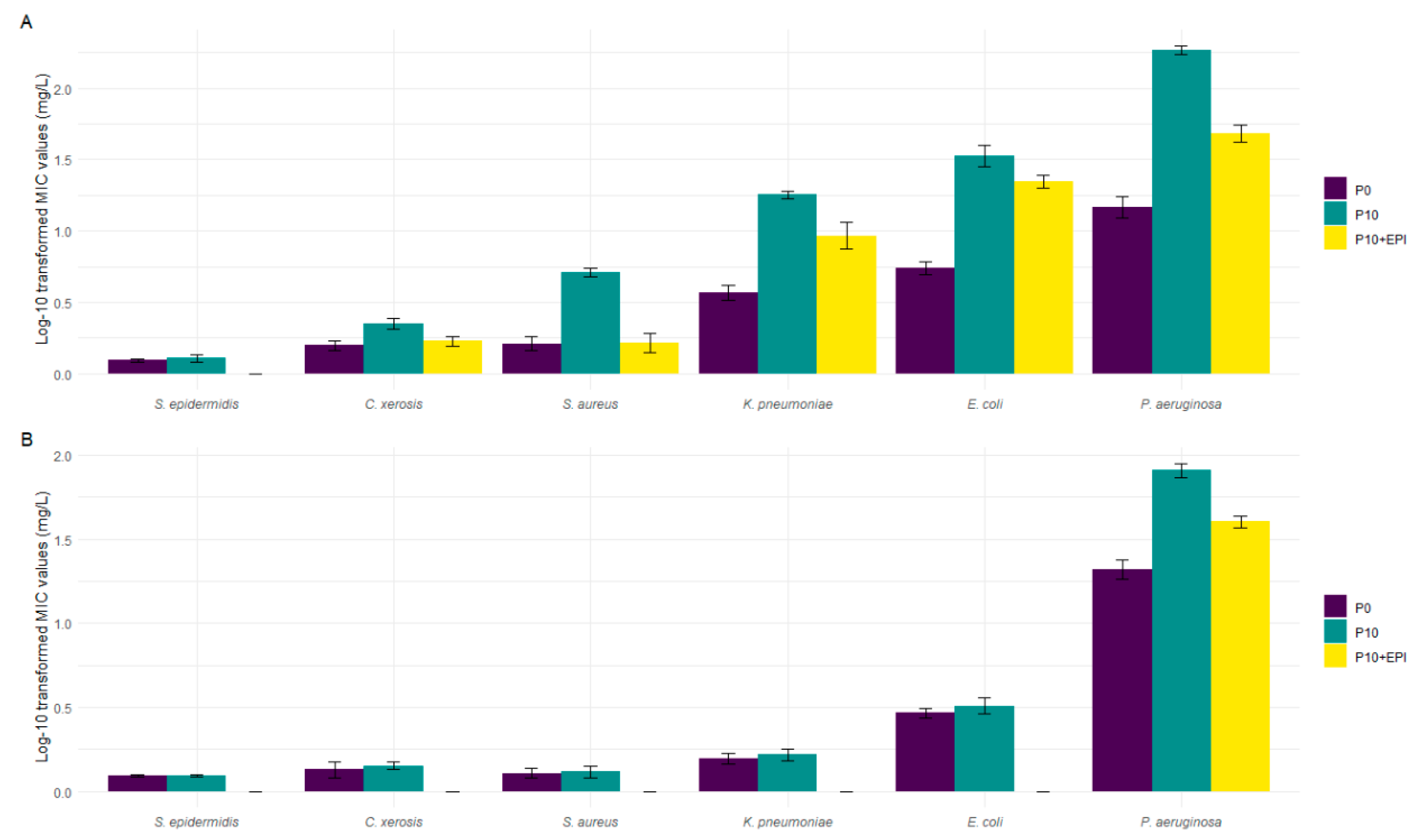
3.4. Treatment with an Efflux Pump Inhibitor (EPI) Reduces Tolerance and Restores Sensitivity to BAC-Inhibition
3.5. BAC Tolerance Is Associated with Antibiotic Cross-Tolerance in BAC-Acclimated Bacteria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kwaśniewska, D.; Chen, Y.; Wieczorek, D. Biological Activity of Quaternary Ammonium Salts and Their Derivatives. Pathogens 2020, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cui, F.; Zeng, G.; Jiang, M.; Yang, Z.; Yu, Z.; Zhu, M.; Shen, L. Quaternary ammonium compounds (QACs): A review on occurrence, fate and toxicity in the environment. Sci. Total Environ. 2015, 518–519, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Luz, A.; DeLeo, P.; Pechacek, N.; Freemantle, M. Human health hazard assessment of quaternary ammonium compounds: Didecyl dimethyl ammonium chloride and alkyl (C12–C16) dimethyl benzyl ammonium chloride. Regul. Toxicol. Pharmacol. 2020, 116, 104717. [Google Scholar] [CrossRef] [PubMed]
- Bondurant, S.W.; Duley, C.M.; Harbell, J.W. Demonstrating the persistent antibacterial efficacy of a hand sanitizer containing benzalkonium chloride on human skin at 1, 2, and 4 hours after application. Am. J. Infect. Control 2019, 47, 928–932. [Google Scholar] [CrossRef] [Green Version]
- PRN 88-2: Clustering of Quaternary Ammonium Compounds. Available online: https://www.epa.gov/pesticide-registration/prn-88-2-clustering-quaternary-ammonium-compounds (accessed on 3 August 2020).
- Hora, P.I.; Pati, S.G.; McNamara, P.J.; Arnold, W.A. Increased Use of Quaternary Ammonium Compounds during the SARS-CoV-2 Pandemic and Beyond: Consideration of Environmental Implications. Environ. Sci. Technol. Lett. 2020, 7, 622–631. [Google Scholar] [CrossRef]
- Gerba, C.P. Quaternary ammonium biocides: Efficacy in application. Appl. Environ. Microbiol. 2015, 81, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Hrubec, T.C.; Seguin, R.P.; Xu, L.; Cortopassi, G.A.; Datta, S.; Hanlon, A.L.; Lozano, A.J.; McDonald, V.A.; Healy, C.A.; Anderson, T.C.; et al. Altered toxicological endpoints in humans from common quaternary ammonium compound disinfectant exposure. Toxicol. Rep. 2021, 8, 646–656. [Google Scholar] [CrossRef]
- Melin, V.E.; Potineni, H.; Hunt, P.; Griswold, J.; Siems, B.; Werre, S.R.; Hrubec, T.C. Exposure to common quaternary ammonium disinfectants decreases fertility in mice. Reprod. Toxicol. 2014, 50, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J. Hormesis: Why it is important to toxicology and toxicologists. Environ. Toxicol. Chem. 2008, 27, 1451–1474. [Google Scholar] [CrossRef]
- EPA’s List of Approved SARS-CoV-2 Surface Disinfectant Products Passes 500. Available online: https://www.epa.gov/newsreleases/epas-list-approved-sars-cov-2-surface-disinfectant-products-passes-500 (accessed on 26 April 2023).
- About List N: Disinfectants for Coronavirus (COVID-19). Available online: https://www.epa.gov/coronavirus/about-list-n-disinfectants-coronavirus-covid-19-0 (accessed on 26 April 2023).
- Kuehn, B.M. More Than 1 in 3 US Adults Use Disinfectants Unsafely. JAMA 2020, 324, 328. [Google Scholar] [CrossRef]
- Rosenman, K.D.; Reilly, M.J.; Wang, L. Calls to a State Poison Center Concerning Cleaners and Disinfectants from the Onset of the COVID-19 Pandemic Through April 2020. Public Health Rep. 2021, 136, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Soave, P.M.; Grassi, S.; Oliva, A.; Romanò, B.; Di Stasio, E.; Dominici, L.; Pascali, V.; Antonelli, M. Household disinfectant exposure during the COVID-19 pandemic: A retrospective study of the data from an Italian poison control center. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 1738–1742. [Google Scholar] [CrossRef] [PubMed]
- Sousa-Silva, M.; Simões, M.; Melo, L.; Machado, I. Pseudomonas fluorescens tolerance to benzyldimethyldodecyl ammonium chloride: Altered phenotype and cross-resistance. J. Glob. Antimicrob. Resist. 2018, 15, 188–195. [Google Scholar] [CrossRef]
- Knapp, L.; Amézquita, A.; McClure, P.; Stewart, S.; Maillard, J. Development of a protocol for predicting bacterial resistance to microbicides. Appl. Environ. Microbiol. 2015, 81, 2652–2659. [Google Scholar] [CrossRef] [Green Version]
- Tandukar, M.; Oh, S.; Tezel, U.; Konstantinidis, K.T.; Pavlostathis, S.G. Long-term exposure to benzalkonium chloride disinfectants results in change of microbial community structure and increased antimicrobial resistance. Environ. Sci. Technol. 2013, 47, 9730–9738. [Google Scholar] [CrossRef] [PubMed]
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist. 2010, 16, 91–104. [Google Scholar] [CrossRef]
- Rozman, U.; Pušnik, M.; Kmetec, S.; Duh, D.; Šostar Turk, S. Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review. Microorganisms 2021, 9, 2550. [Google Scholar] [CrossRef]
- WHO|Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 17 October 2020).
- EU Action on Antimicrobial Resistance. Available online: https://health.ec.europa.eu/antimicrobial-resistance/eu-action-antimicrobial-resistance_en (accessed on 18 May 2023).
- Bengoechea, J.A.; Bamford, C.G. SARS-CoV-2, bacterial co-infections, and AMR: The deadly trio in COVID-19? EMBO Mol. Med. 2020, 12, e12560. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, A.R.; Safaee, M.M.; Wuest, W.M.; Furst, A.L. The silent pandemic: Emergent antibiotic resistances following the global response to SARS-CoV-2. iScience 2021, 24, 102304. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G. Biocidal Agents Used for Disinfection Can Enhance Antibiotic Resistance in Gram-Negative Species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Hardy, K.; Sunnucks, K.; Gil, H.; Shabir, S.; Trampari, E.; Hawkey, P.; Webber, M. Increased Usage of Antiseptics Is Associated with Reduced Susceptibility in Clinical Isolates of Staphylococcus aureus. mBio 2018, 9, e00894-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.; Gemmell, C.G.; Hunter, I.S. The association between biocide tolerance and the presence or absence of qac genes among hospital-acquired and community-acquired MRSA isolates. J. Antimicrob. Chemother. 2008, 61, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saá Ibusquiza, P.; Herrera, J.J.R.; Vázquez-Sánchez, D.; Parada, A.; Cabo, M.L. A new and efficient method to obtain benzalkonium chloride adapted cells of Listeria monocytogenes. J. Microbiol. Methods 2012, 91, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Aguinaga, U.; Oyinwola, F.J.; Childs, M.J.; Aldarkazanly, S.; Yousufuddin, M. Benzylhexadecyldimethylammonium chloride dihydrate. IUCrData 2023, 8, x230096. [Google Scholar] [CrossRef]
- M100Ed32|Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 19 February 2023).
- Wang, Y.; Venter, H.; Ma, S. Efflux Pump Inhibitors: A Novel Approach to Combat Efflux-Mediated Drug Resistance in Bacteria. Curr. Drug Targets 2016, 17, 702–719. [Google Scholar] [CrossRef]
- Kaur, B.; Gupta, J.; Sharma, S.; Sharma, D.; Sharma, S. Focused review on dual inhibition of quorum sensing and efflux pumps: A potential way to combat multi drug resistant Staphylococcus aureus infections. Int. J. Biol. Macromol. 2021, 190, 33–43. [Google Scholar] [CrossRef]
- Jiang, X.; Yu, T.; Liang, Y.; Ji, S.; Guo, X.; Ma, J.; Zhou, L. Efflux pump-mediated benzalkonium chloride resistance in Listeria monocytogenes isolated from retail food. Int. J. Food Microbiol. 2016, 217, 141–145. [Google Scholar] [CrossRef]
- Buffet-Bataillon, S.; Tattevin, P.; Maillard, J.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Efflux pump induction by quaternary ammonium compounds and fluoroquinolone resistance in bacteria. Future Microbiol. 2016, 11, 81–92. [Google Scholar] [CrossRef]
- Buffet-Bataillon, S.; Tattevin, P.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Emergence of resistance to antibacterial agents: The role of quaternary ammonium compounds--a critical review. Int. J. Antimicrob. Agents 2012, 39, 381–389. [Google Scholar] [CrossRef]
- Kim, M.; Weigand, M.R.; Oh, S.; Hatt, J.K.; Krishnan, R.; Tezel, U.; Pavlostathis, S.G.; Konstantinidis, K.T. Widely Used Benzalkonium Chloride Disinfectants Can Promote Antibiotic Resistance. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Lai, Y.S.; Zheng, C.; Ilhan, Z.E.; Ontiveros-Valencia, A.; Long, X.; Krajmalnik-Brown, R.; Rittmann, B.E. Increased expression of antibiotic-resistance genes in biofilm communities upon exposure to cetyltrimethylammonium bromide (CTAB) and other stress conditions. Sci. Total Environ. 2021, 765, 144264. [Google Scholar] [CrossRef]
- Dashtbani-Roozbehani, A.; Brown, M.H. Efflux Pump Mediated Antimicrobial Resistance by Staphylococci in Health-Related Environments: Challenges and the Quest for Inhibition. Antibiotics 2021, 10, 1502. [Google Scholar] [CrossRef]
- Lamut, A.; Peterlin Mašič, L.; Kikelj, D.; Tomašič, T. Efflux pump inhibitors of clinically relevant multidrug resistant bacteria. Med. Res. Rev. 2019, 39, 2460–2504. [Google Scholar] [CrossRef]
- Monteiro, K.L.C.; de Aquino, T.M.; Mendonça Junior, F.J.B. An Update on Staphylococcus aureus NorA Efflux Pump Inhibitors. Curr. Top. Med. Chem. 2020, 20, 2168–2185. [Google Scholar] [CrossRef] [PubMed]
- Garrison, M.A.; Mahoney, A.R.; Wuest, W.M. Tricepyridinium-inspired QACs yield potent antimicrobials and provide insight into QAC resistance. ChemMedChem 2021, 16, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Meade, E.; Slattery, M.A.; Garvey, M. Biocidal Resistance in Clinically Relevant Microbial Species: A Major Public Health Risk. Pathogens 2021, 10, 598. [Google Scholar] [CrossRef]
- Richards, R.M.; Mizrahi, L.M. Differences in antibacterial activity of benzalkonium chloride. J. Pharm. Sci. 1978, 67, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Nasr, A.M.; Mostafa, M.S.; Arnaout, H.H.; Elshimy, A.A.A. The effect of exposure to sub-inhibitory concentrations of hypochlorite and quaternary ammonium compounds on antimicrobial susceptibility of Pseudomonas aeruginosa. Am. J. Infect. Control 2018, 46, e57–e63. [Google Scholar] [CrossRef] [Green Version]
- Langsrud, S.; Sundheim, G.; Holck, A.L. Cross-resistance to antibiotics of Escherichia coli adapted to benzalkonium chloride or exposed to stress-inducers. J. Appl. Microbiol. 2004, 96, 201–208. [Google Scholar] [CrossRef]
- Loughlin, M.F.; Jones, M.V.; Lambert, P.A. Pseudomonas aeruginosa cells adapted to benzalkonium chloride show resistance to other membrane-active agents but not to clinically relevant antibiotics. J. Antimicrob. Chemother. 2002, 49, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Nordholt, N.; Kanaris, O.; Schmidt, S.B.I.; Schreiber, F. Persistence against benzalkonium chloride promotes rapid evolution of tolerance during periodic disinfection. Nat. Commun. 2021, 12, 6792. [Google Scholar] [CrossRef]
- Mc Cay, P.H.; Ocampo-Sosa, A.A.; Fleming, G.T.A. Effect of subinhibitory concentrations of benzalkonium chloride on the competitiveness of Pseudomonas aeruginosa grown in continuous culture. Microbiology 2010, 156, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Stone, W.; Tolmay, J.; Tucker, K.; Wolfaardt, G.M. Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion. Microorganisms 2020, 8, 1726. [Google Scholar] [CrossRef] [PubMed]
- Chemaly, R.F.; Simmons, S.; Dale, C.; Ghantoji, S.S.; Rodriguez, M.; Gubb, J.; Stachowiak, J.; Stibich, M. The role of the healthcare environment in the spread of multidrug-resistant organisms: Update on current best practices for containment. Ther. Adv. Infect. Dis. 2014, 2, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Cave, R.; Cole, J.; Mkrtchyan, H.V. Surveillance and prevalence of antimicrobial resistant bacteria from public settings within urban built environments: Challenges and opportunities for hygiene and infection control. Environ. Int. 2021, 157, 106836. [Google Scholar] [CrossRef]
- He, G.; Landry, M.; Chen, H.; Thorpe, C.; Walsh, D.; Varela, M.F.; Pan, H. Detection of benzalkonium chloride resistance in community environmental isolates of staphylococci. J. Med. Microbiol. 2014, 63, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Filippelli, G.M.; Salamova, A. Increased Indoor Exposure to Commonly Used Disinfectants during the COVID-19 Pandemic. Environ. Sci. Technol. Lett. 2020, 7, 760–765. [Google Scholar] [CrossRef]
- Boles, C.; Maier, A.; Vincent, M.; Stewart, C.; Attar, S.; Yeomans, D. Multi-route exposure sampling of quaternary ammonium compounds and ethanol surface disinfectants in a K-8 school. Indoor Air 2022, 32, e13036. [Google Scholar] [CrossRef]
- Li, D.; Sangion, A.; Li, L. Evaluating consumer exposure to disinfecting chemicals against coronavirus disease 2019 (COVID-19) and associated health risks. Environ. Int. 2020, 145, 106108. [Google Scholar] [CrossRef]
- Zhang, M.; O’dononghue, M.; Boost, M.V. Characterization of staphylococci contaminating automated teller machines in Hong Kong. Epidemiol. Infect. 2012, 140, 1366–1371. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.J.; Rutala, W.A.; Sickbert-Bennett, E.E. Outbreaks associated with contaminated antiseptics and disinfectants. Antimicrob. Agents Chemother. 2007, 51, 4217–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T. The role of the healthcare environment in the acquisition of infection. Br. J. Nurs. 2020, 29, 8. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Webster, T.F.; Salamova, A. Quaternary Ammonium Compounds: Bioaccumulation Potentials in Humans and Levels in Blood before and during the Covid-19 Pandemic. Environ. Sci. Technol. 2021, 55, 14689–14698. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Schreder, E.; Sathyanarayana, S.; Salamova, A. The first detection of quaternary ammonium compounds in breast milk: Implications for early-life exposure. J. Expo. Sci. Environ. Epidemiol. 2022, 32, 682–688. [Google Scholar] [CrossRef]


{kind=link}
| MIC (mg/L) | BAC12–14 | BAC16 |
|---|---|---|
| S. epidermidis | 0.24 ± 0.03 | 0.24 ± 0.02 |
| C. xerosis | 0.58 ± 0.12 | 0.40 ± 0.13 |
| S. aureus | 0.63 ± 0.18 | 0.28 ± 0.08 |
| K. pneumoniae | 2.69 ± 0.43 | 0.56 ± 0.11 |
| E. coli | 4.50 ± 0.50 | 1.94 ± 0.17 |
| P. aeruginosa | 13.75 ± 2.17 | 20.00 ± 2.50 |
| MIC (mg/L) | BAC12–14 | BAC16 | ||
|---|---|---|---|---|
| P10 | P10 + EPI | P10 | P10 + EPI | |
| S. epidermidis | 0.28 ± 0.10 | NT | 0.24 ± 0.20 | NT |
| C. xerosis | 1.25 ± 0.20 * | 0.69 ± 0.10 | 0.40 ± 0.10 | NT |
| S. aureus | 4.13 ± 0.30 * | 0.66 ± 0.20 ** | 0.31 ± 0.11 | NT |
| K. pneumoniae | 17.00 ± 1.00 * | 8.44 ± 1.70 **# | 0.66 ± 0.12 | NT |
| E. coli | 33.13 ± 5.60 * | 21.25 ± 2.20 **# | 2.25 ± 0.35 | NT |
| P. aeruginosa | 184.38 ± 12.1 * | 47.50 ± 6.60 **# | 80.63 ± 7.26 * | 39.38 ± 3.00 **# |
| Neomycin | Bacitracin | Chloramphenicol | Streptomycin | Penicillin | |
|---|---|---|---|---|---|
| S. epidermidis | |||||
| Baseline (P0) | S | I | S | S | I |
| BAC12–14 (P10) | S | I | S | S | I |
| BAC16 (P10) | S | I | S | S | I |
| C. xerosis | |||||
| Baseline (P0) | S | S | S | S | R |
| BAC12–14 (P10) | I * | I * | S | S | R |
| BAC16 (P10) | S | S | S | S | R |
| S. aureus | |||||
| Baseline (P0) | S | I | S | S | S |
| BAC12–14 (P10) | S | I | S | S | I * |
| BAC16 (P10) | S | I | S | S | S |
| K. pneumoniae | |||||
| Baseline (P0) | S | R | S | S | R |
| BAC12–14 (P10) | S | R | I * | I * | R |
| BAC16 (P10) | S | R | S | S | R |
| E. coli | |||||
| Baseline (P0) | S | R | S | S | R |
| BAC12–14 (P10) | I * | R | S | S | R |
| BAC16 (P10) | S | R | S | S | R |
| P. aeruginosa | |||||
| Baseline (P0) | R | R | R | I | R |
| BAC12–14 (P10) | R | R | R | R * | R |
| BAC16 (P10) | R | R | R | R * | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pena, S.A.; Salas, J.G.; Gautam, N.; Ramos, A.M.; Frantz, A.L. Sublethal Exposure to Common Benzalkonium Chloride Leads to Antimicrobial Tolerance and Antibiotic Cross-Resistance in Commensal and Opportunistic Bacterial Species. Appl. Microbiol. 2023, 3, 580-591. https://doi.org/10.3390/applmicrobiol3020041
Pena SA, Salas JG, Gautam N, Ramos AM, Frantz AL. Sublethal Exposure to Common Benzalkonium Chloride Leads to Antimicrobial Tolerance and Antibiotic Cross-Resistance in Commensal and Opportunistic Bacterial Species. Applied Microbiology. 2023; 3(2):580-591. https://doi.org/10.3390/applmicrobiol3020041
Chicago/Turabian StylePena, Sheareazade A., Juana G. Salas, Nilisha Gautam, Ashley M. Ramos, and Aubrey L. Frantz. 2023. "Sublethal Exposure to Common Benzalkonium Chloride Leads to Antimicrobial Tolerance and Antibiotic Cross-Resistance in Commensal and Opportunistic Bacterial Species" Applied Microbiology 3, no. 2: 580-591. https://doi.org/10.3390/applmicrobiol3020041