
The Growth Medium Affects the Viability of IPEC-J2 Animal Cell Line in the Presence of Probiotic Bacteria


Abstract
:1. Introduction
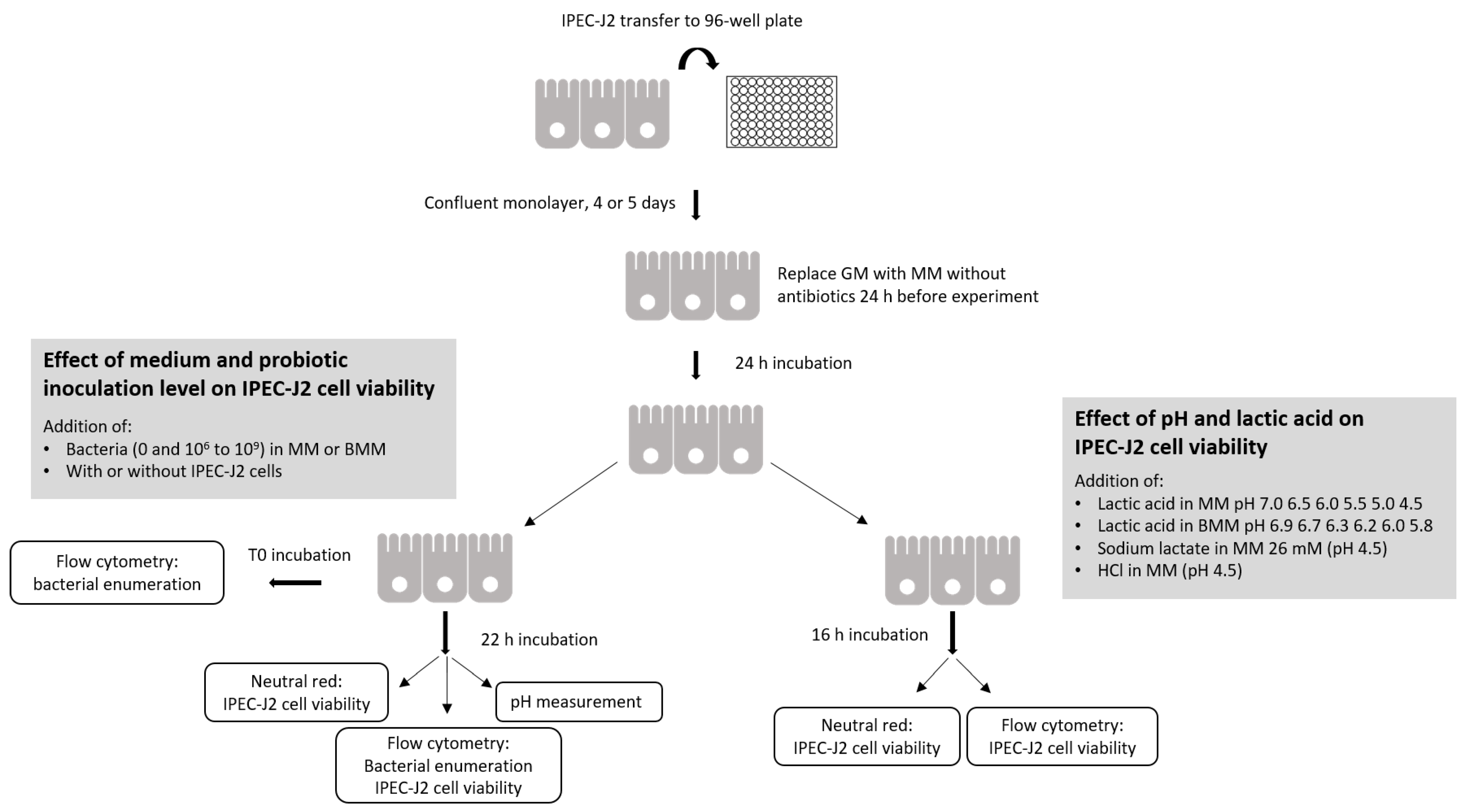
2. Materials and Methods
2.1. IPEC-J2 Cells
2.2. Bacterial Strains
2.3. Addition of Probiotics and Bacteria to Monolayer of IPEC-J2 Cells
2.4. Enumeration of Probiotic Bacteria
2.5. Cytotoxicity Assays with IPEC-J2 Cells
- a = OD derived from the wells incubated with probiotics;
- b = OD derived from blank wells;
- c = OD derived from the positive control.
2.6. Effect of Lactic Acid, Sodium Lactate and HCL on IPEC-J2 Viability
2.7. Statistical Analyses
3. Results and Discussion
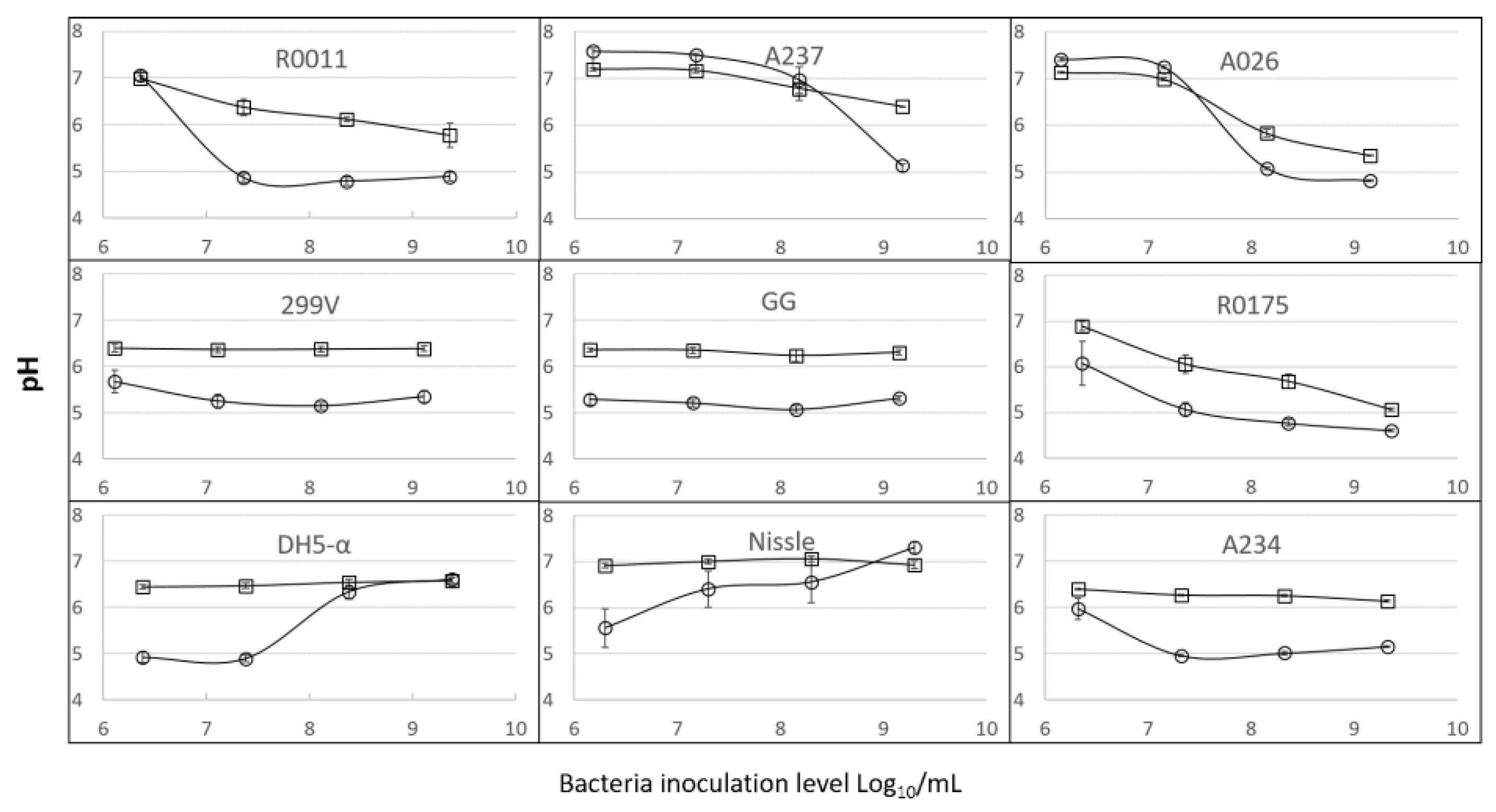
3.1. Growth and Acidification of the Media by the Lactobacilli and Bifidobacteria (LB) Cultures
3.2. Growth and Acidification of the Media by E. coli Strains
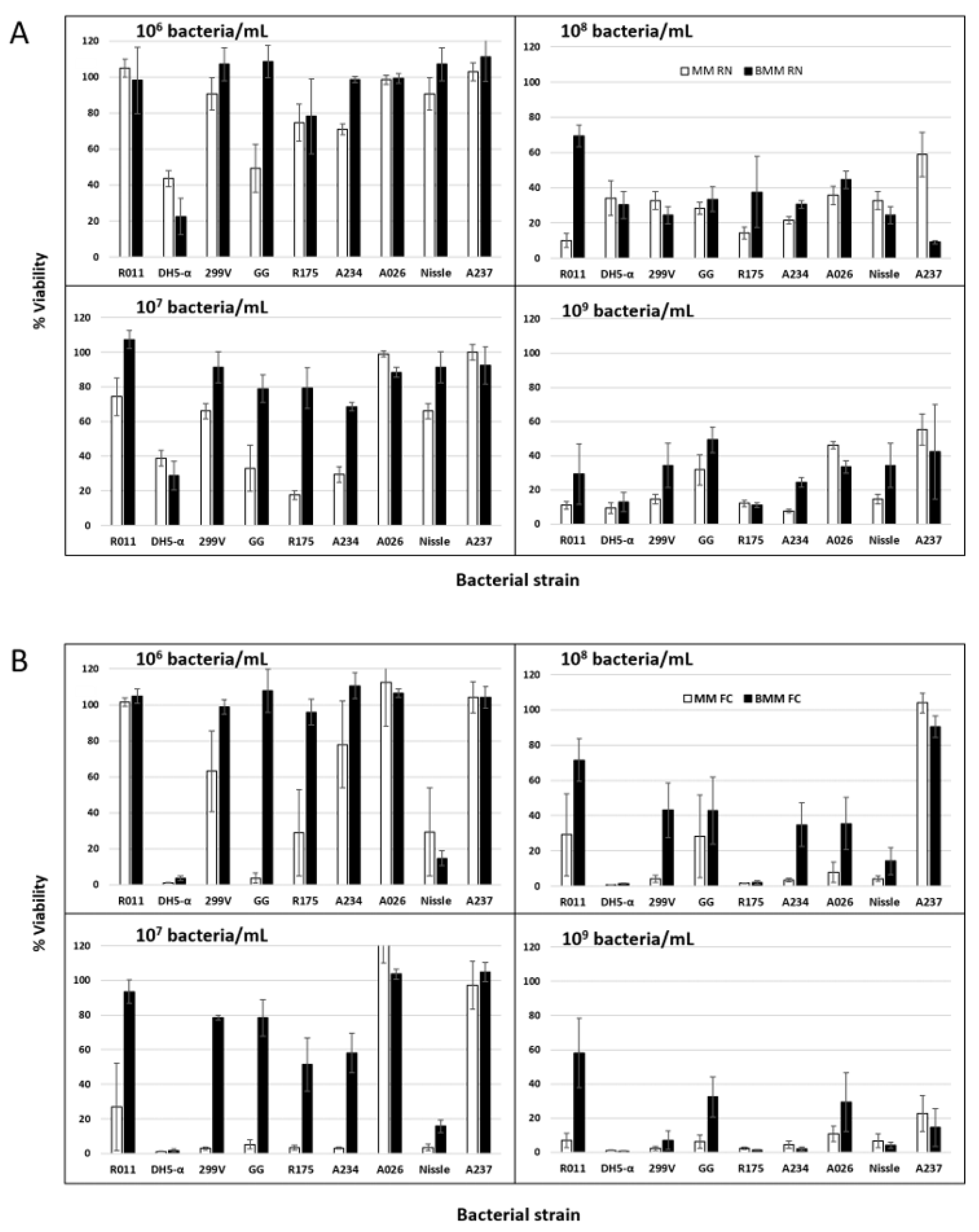
3.3. Effect of Probiotic Inoculation Level on IPEC-J2 Cell Viability as Ascertained by Neutral Red
3.4. Effect of Inoculation Level of E. coli Cultures on IPEC-J2 Cell Viability
3.5. Determination of IPEC-J2 Cell Viability by Flow Cytometry
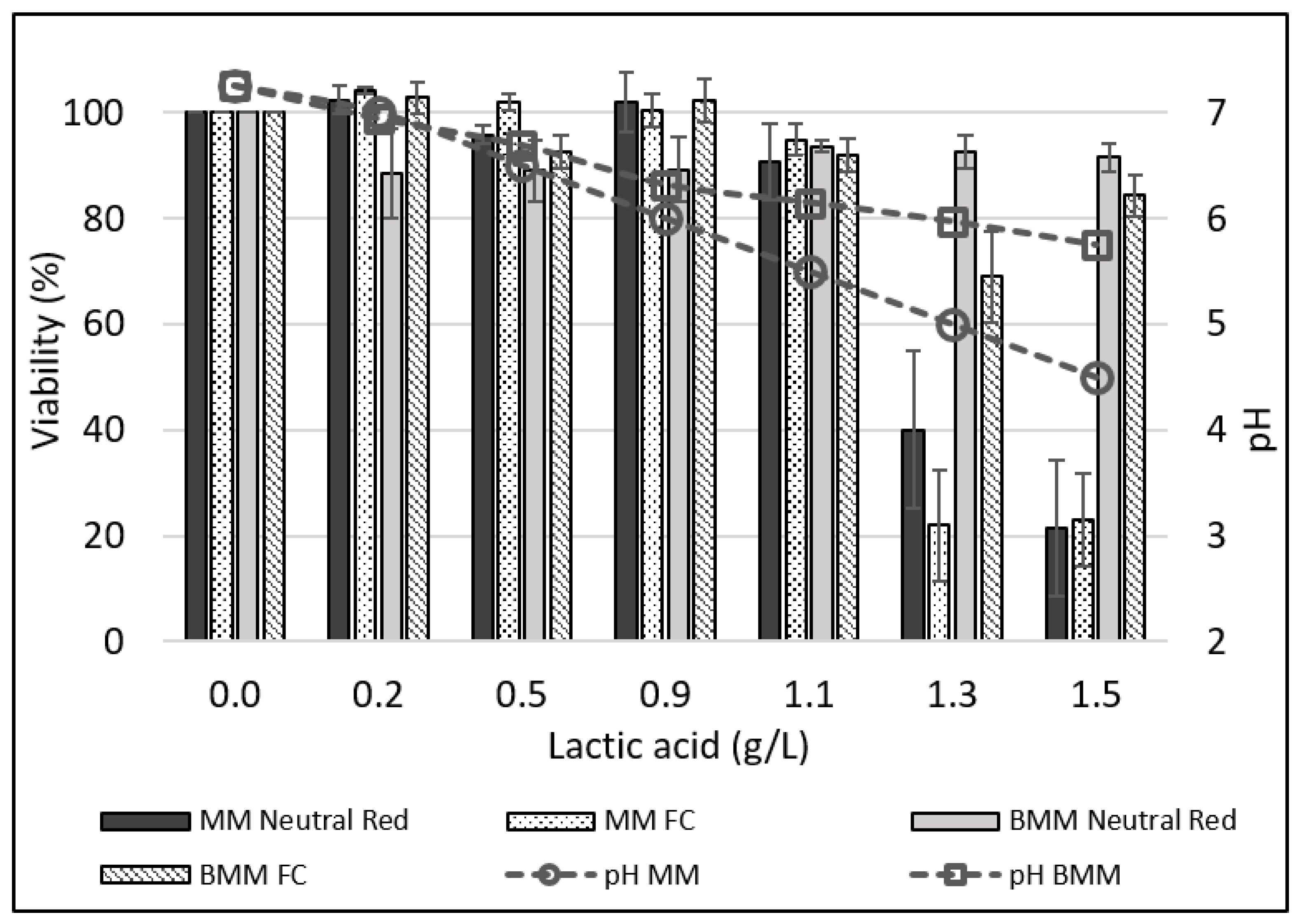
3.6. Effect of pH and Lactic Acid on IPEC-J2 Cell Viability
4. Conclusions
- (1)
- Preliminary trials must be carried out to identify the experimental conditions allowing the highest inoculation level at which a given probiotic strain does not negatively affect the viability of the animal cell cultures; this needs to be determined for each bacterial strain.
- (2)
- The pH of the medium as well as the bacterial counts should be registered, not just at the beginning of the experiment, but also at the end of the incubation.
- (3)
- New media are required; increasing the buffering capacity of the traditional media for IPEC-J2 cultures reduces the drop in pH, which affects the cell viability. This allows for the use of higher inoculation levels of probiotics, which can provide health benefits for the animal cell cultures. Nevertheless, more research should be carried out on how to improve buffering of the medium for animal cell lines when probiotic cultures are inoculated.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Blais, M.; Fortier, M.; Pouliot, Y.; Gauthier, S.F.; Boutin, Y.; Asselin, C.; Lessard, M. Colostrum whey down-regulates the expression of early and late inflammatory response genes induced by Escherichia coli and Salmonella enterica Typhimurium components in intestinal epithelial cells. Br. J. Nutr. 2015, 113, 200–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botić, T.; Klingberg, T.D.; Weingartl, H.; Cencič, A. A novel eukaryotic cell culture model to study antiviral activity of potential probiotic bacteria. Int. J. Food Microbiol. 2007, 115, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Geens, M.M.; Niewold, T.A. Optimizing culture conditions of a porcine epithelial cell line IPEC-J2 through a histological and physiological characterization. Cytotechnology 2011, 63, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergauwen, H. The IPEC-J2 cell-line. In The Impact of Bio-Actives on Gut Health; Verhoeckx, K., Cotter, P., Lopez-Exposito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzeraland, 2015; Volume 12, pp. 125–134. [Google Scholar]
- Karimi, S.; Jonsson, H.; Lundh, T.; Roos, S. Lactobacillus reuteri strains protect epithelial barrier integrity of IPEC-J2 monolayers from the detrimental effect of enterotoxigenic Escherichia coli. Physiol. Rep. 2018, 6, e13514. [Google Scholar] [CrossRef] [Green Version]
- Skjolaas, K.A.; Burkey, T.E.; Dritz, S.S.; Minton, J.E. Effects of Salmonella enterica serovar Typhimurium, or serovar choleraesuis, lactobacillus reuteri and bacillus licheniformis on chemokine and cytokine expression in the swine jejunal epithelial cell line, IPEC-J2. Vet. Immunol. Immunopathol. 2007, 115, 299–308. [Google Scholar] [CrossRef]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Human-derived probiotic Lactobacillus reuteri strains differentially reduce intestinal inflammation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 299, G1087–G1096. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.W.; El-Nezami, H.; Shah, N.P. Effects of supplementation of citrulline and Lactobacillus helveticus ascc 511 on intestinal epithelial cell integrity. J. Funct. Foods 2020, 64, 103571. [Google Scholar] [CrossRef]
- Hu, J.; Ma, L.; Zheng, W.; Nie, Y.; Yan, X. Lactobacillus gasseri la39 activates the oxidative phosphorylation pathway in porcine intestinal epithelial cells. Front. Microbiol. 2018, 9, 3025. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Qiao, L.; Liu, R.; Yao, H.; Gao, C. Potential probiotic properties of lactic acid bacteria isolated from the intestinal mucosa of healthy piglets. Ann. Microbiol. 2017, 67, 239–253. [Google Scholar] [CrossRef]
- Liu, H.Y.; Roos, S.; Jonsson, H.; Ahl, D.; Dicksved, J.; Lindberg, J.E.; Lundh, T. Effects of Lactobacillus johnsonii and Lactobacillus reuteri on gut barrier function and heat shock proteins in intestinal porcine epithelial cells. Physiol. Rep. 2015, 3, e12355. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, C.; Chen, Z.; Chen, Z.; Zhang, W.; Ma, X.; Wang, L.; Yang, X.; Jiang, Z. Protective effects of Lactobacillus plantarum on epithelial barrier disruption caused by enterotoxigenic Escherichia coli in intestinal porcine epithelial cells. Vet. Immunol. Immunopathol. 2016, 172, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Nissen, P.; Willats, W.G.T. The effect of calcium ions on adhesion and competitive exclusion of Lactobacillus ssp. and E. coli o138. Int. J. Food Microbiol. 2007, 114, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, Y.H.; Yang, J.C.; Yang, G.Y.; Zhou, D.; Wang, J.F. A selected Lactobacillus rhamnosus strain promotes egfr-independent akt activation in an enterotoxigenic Escherichia coli k88-infected IPEC-J2 cell model. PLoS ONE 2015, 10, 0125717. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Ye, P.; Lei, Q.; Cheng, Y.; Yu, H.; Du, J.; Pan, H.; Cao, Z. In vitro probiotic properties of pediococcus pentosaceus l1 and its effects on enterotoxigenic Escherichia coli-induced inflammatory responses in porcine intestinal epithelial cells. Microb. Pathog. 2020, 144, 104163. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Sun, Z.; Liang, D.; Li, H. Lactobacillus salivarius alleviates inflammation via nf-κb signaling in etec k88-induced IPEC-J2 cells. J. Anim. Sci. Biotechnol. 2020, 11, 76. [Google Scholar] [CrossRef]
- Zhu, C.; Lv, Y.; Yang, J.; Bai, Y.; Ye, J.; Wang, Z.; Chen, Z.; Jiang, Z. Proteomic alteration of porcine intestinal epithelial cells after pretreatment with Lactobacillus plantarum followed by infection with enterotoxigenic Escherichia coli f4. Vet. Immunol. Immunopathol. 2020, 222, 109943. [Google Scholar] [CrossRef]
- Xu, C.; Yan, S.; Guo, Y.; Qiao, L.; Ma, L.; Dou, X.; Zhang, B. Lactobacillus casei atcc 393 alleviates enterotoxigenic Escherichia coli k88-induced intestinal barrier dysfunction via tlrs/mast cells pathway. Life Sci. 2020, 244, 117281. [Google Scholar] [CrossRef]
- Xia, B.; Yu, J.; He, T.; Liu, X.; Su, J.; Wang, M.; Wang, J.; Zhu, Y. Lactobacillus johnsonii l531 ameliorates enteritis via elimination of damaged mitochondria and suppression of sqstm1-dependent mitophagy in a Salmonella infantis model of piglet diarrhea. FASEB J. 2020, 34, 2821–2839. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Kleta, S.; Tedin, K.; Babila, J.T.; Oswald, S.; Oelschlaeger, T.A.; Hiemann, R.; Paetzold, S.; Wieler, L.H.E. E. coli nissle 1917 affects salmonella adhesion to porcine intestinal epithelial cells. PLoS ONE 2011, 6, e14712. [Google Scholar] [CrossRef] [Green Version]
- Loss, H.; Aschenbach, J.R.; Tedin, K.; Ebner, F.; Lodemann, U. The inflammatory response to enterotoxigenic E. coli and probiotic E. faecium in a coculture model of porcine intestinal epithelial and dendritic cells. Mediat. Inflamm. 2018, 2018, 9368295. [Google Scholar] [CrossRef]
- Pahumunto, N.; Dahlen, G.; Teanpaisan, R. Evaluation of potential probiotic properties of Lactobacillus and Bacillus strains derived from various sources for their potential use in swine feeding. Probiotics Antimicrob. Proteins 2021. [Google Scholar] [CrossRef]
- Chen, S.; Li, Y.; Chu, B.; Yuan, L.; Liu, N.; Zhu, Y.; Wang, J. Lactobacillus johnsonii l531 alleviates the damage caused by Salmonella typhimurium via inhibiting tlr4, nf-κb, and nlrp3 inflammasome signaling pathways. Microorganisms 2021, 9, 1983. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, D.; Raymond, Y.; Lemay, M.J.; Champagne, C.P.; Brassard, J. Effect of probiotic bacteria on porcine rotavirus osu infection of porcine intestinal epithelial IPEC-J2 cells. Arch. Virol. 2022, 167, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, G.; Wen, K.; Bui, T.; Cao, D.; Zhang, Y.; Yuan, L. Porcine small intestinal epithelial cell line (IPEC-J2) of rotavirus infection as a new model for the study of innate immune responses to rotaviruses and probiotics. Viral Immunol. 2010, 23, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Salaün, F.; Mietton, B.; Gaucheron, F. Buffering capacity of dairy products. Int. Dairy J. 2005, 15, 95–109. [Google Scholar] [CrossRef]
- Champagne, C.P.; Piette, M.; Saint-Gelais, D. Characteristics of lactococci cultures produced in commercial media. J. Ind. Microbiol. 1995, 15, 472–479. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Lee, D.K.; Park, J.E.; Kim, M.J.; Seo, J.G.; Lee, J.H.; Ha, N.J. Probiotic bacteria, B. longum and L. acidophilus inhibit infection by rotavirus in vitro and decrease the duration of diarrhea in pediatric patients. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 237–244. [Google Scholar] [CrossRef]
- Liu, F.; Li, G.; Wen, K.; Wu, S.; Zhang, Y.; Bui, T.; Yang, X.; Kocher, J.; Sun, J.; Jortner, B.; et al. Lactobacillus rhamnosus gg on rotavirus-induced injury of ileal epithelium in gnotobiotic pigs. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Díaz, J.; Monedero, V. Probiotics against digestive tract viral infections. In Bioactive Food as Dietary Interventions for Liver and Gastrointestinal Disease; Elsevier: Amsterdam, The Netherlands, 2013; pp. 271–284. [Google Scholar]
- Raymond, Y.; Champagne, C.P. The use of flow cytometry to accurately ascertain total and viable counts of Lactobacillus rhamnosus in chocolate. Food Microbiol. 2015, 46, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Champagne, C.P.; Ross, R.P.; Saarela, M.; Hansen, K.F.; Charalampopoulos, D. Recommendations for the viability assessment of probiotics as concentrated cultures and in food matrices. Int. J. Food Microbiol. 2011, 149, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/ cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Champagne, C.P.; Gomes da Cruz, A.; Daga, M. Strategies to improve the functionality of probiotics in supplements and foods. Curr. Opin. Food Sci. 2018, 22, 160–166. [Google Scholar] [CrossRef]
- Sanders, M.E.; Benson, A.; Lebeer, S.; Merenstein, D.J.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr. Opin. Biotechnol. 2018, 49, 207–216. [Google Scholar] [CrossRef]
- Braude, R.; Fulford, R.J.; Low, A.G. Studies on digestion and absorption in the intestines of growing pigs. Measurements of the flow of digesta and pH. Br. J. Nutr. 1976, 36, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Andersson, S. Secretion of gastrointestinal hormones. Annu. Rev. Physiol. 1973, 35, 431–452. [Google Scholar] [CrossRef]
- Zakrzewski, R.; Lee, K.; Lye, G.J. Development of a miniature bioreactor model to study the impact of pH and DOT fluctuations on CHO cell culture performance as a tool to understanding heterogeneity effects at large-scale. Biotechnol. Prog. 2022, 38, e3264. [Google Scholar] [CrossRef]
- Bolduc, M.-P.; Raymond, Y.; Fustier, P.; Champagne, C.P.; Vuillemard, J.-C. Sensitivity of bifidobacteria to oxygen and redox potential in non-fermented pasteurized milk. Int. Dairy J. 2006, 16, 1038–1048. [Google Scholar] [CrossRef]
- Mohammadi, R.; Sohrabvandi, S.; Mohammad Mortazavian, A. The starter culture characteristics of probiotic microorganisms in fermented milks. Eng. Life Sci. 2012, 12, 399–409. [Google Scholar] [CrossRef]
- Turner, K.W.; Thomas, T.D. Uncoupling of growth and acid production in lactic streptococci. N. Z. J. Dairy Sci. Technol. 1975, 10, 162–167. [Google Scholar]
- Johnson, B.J.; Lin, B.; Rubin, R.A.; Malanoski, A.P. Media acidification by Escherichia coli in the presence of cranberry juice. BMC Res. Notes 2009, 2, 226. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.R.; Luli, G.W. A gradient-feed process for E. coli fermentations. Biopharm. Manuf. 1987, 1, 38–41. [Google Scholar]
- Anderson, K.W.; Grulke, E.; Gerhardt, P. Microfiltration culture process for enhanced production of rdna receptor cells of Escherichia coli. Bio/Technology 1984, 2, 891–896. [Google Scholar] [CrossRef]
- Bauer, S.; Ziv, E. Dense growth of aerobic bacteria in a bench scale fermentor. Biotechnol. Bioeng. 1976, 18, 81–94. [Google Scholar] [CrossRef]
- Blankenhorn, D.; Phillips, J.; Slonczewski, J.L. Acid- and base-induced proteins during aerobic and anaerobic growth of Escherichia coli revealed by two-dimensional gel electrophoresis. J. Bacteriol. 1999, 181, 2209–2216. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.D.K.; Jones-Mortimer, M.C.; Kornberg, H.L. The enzymatic interconversion of acetate and acetyl-coenzyme a in Escherichia coli. Microbiology 1977, 102, 327–336. [Google Scholar]
- Doelle, H.W.; Hollywood, N.; Westwood, A.W. Effect of glucose concentration on a number of enzymes involved in the aerobic and anaerobic utilization of glucose in turbidostat cultures of Escherichia coli. Microbios 1974, 9, 221–232. [Google Scholar]
- Hollywood, N.; Doelle, H.W. Effect of specific growth rate and glucose concentration on growth and glucose metabolism of Escherichia coli k 12. Microbios 1976, 17, 23–33. [Google Scholar]
- Landwall, P.; Holme, T. Influence of glucose and dissolved oxygen concentrations on yields of Escherichia coli b in dialysis culture. J. Gen. Microbiol. 1977, 103, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Mori, H.; Yano, T.; Kobayashi, T.; Shimizu, S. High density cultivation of biomass in fed-batch system with do-stat. J. Chem. Eng. Jpn. 1979, 12, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Reiling, H.E.; Laurila, H.; Fiechter, A. Mass culture of Escherichia coli: Medium development for low and high density cultivation of Escherichia coli b/r in minimal and complex media. J. Biotechnol. 1985, 2, 191–206. [Google Scholar] [CrossRef]
- Rinas, U.; Kracke-Helm, H.A.; Schügerl, K. Glucose as a substrate in recombinant strain fermentation technology-by-product formation, degradation and intracellular accumulation of recombinant protein. Appl. Microbiol. Biotechnol. 1989, 31, 163–167. [Google Scholar] [CrossRef]
- Smirnova, G.V.; Oktyabr’skii, O.N. Influence of acetate on the growth of Escherichia coli under aerobic and anaerobic conditions. Microbiology 1985, 54, 252–256. [Google Scholar]
- El-Mansi, E.M.T.; Holms, W.H. Control of carbon flux to acetate excretion during growth of Escherichia coli in batch and continuous cultures. J. Gen. Microbiol. 1989, 135, 2875–2883. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.K.; Lim, C.Y.; Teng, W.L.; Ouwehand, A.C.; Tuomola, E.M.; Salminen, S. Quantitative approach in the study of adhesion of lactic acid bacteria to intestinal cells and their competition with enterobacteria. Appl. Environ. Microbiol. 2000, 66, 3692–3697. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, P.; Esteban, M.A.; Cuesta, A. Heavy metals produce toxicity, oxidative stress ans apoptosis int he marine teleost fish saf-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef]
- Fernandez-Duarte, K.P.; Olaya-Galán, N.N.; Salas-Cárdenas, S.P.; Lopez-Rozo, J.; Gutierrez-Fernandez, M.F. Bifidobacterium adolescentis (dsm 20083) and Lactobacillus casei (lafti l26-dsl): Probiotics able to block the in vitro adherence of rotavirus in ma104 cells. Probiotics Antimicrob. Proteins 2018, 10, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, Q.; Liu, C.M.; Gong, J. Issues deserve attention in encapsulating probiotics: Critical review of existing literature. Crit. Rev. Food Sci. Nutr. 2017, 57, 1228–1238. [Google Scholar] [CrossRef]
- Hering, N.A.; Richter, J.F.; Fromm, A.; Wieser, A.; Hartmann, S.; Günzel, D.; Bücker, R.; Fromm, M.; Schulzke, J.D.; Troeger, H. Tcpc protein from E. coli nissle improves epithelial barrier function involving pkcζ and erk1/2 signaling in ht-29/b6 cells. Mucosal Immunol. 2014, 7, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Rembacken, B.J.; Snelling, A.M.; Hawkey, P.M.; Chalmers, D.M.; Axon, A.T.R. Non-pathogenic Escherichia coli versus mesalazine for the treatment of ulcerative colitis: A randomised trial. Lancet 1999, 354, 635–639. [Google Scholar] [CrossRef]
- Altenhoefer, A.; Oswald, S.; Sonnenborn, U.; Enders, C.; Schulze, J.; Hacker, J.; Oelschlaeger, T.A. The probiotic Escherichia coli strain nissle 1917 interferes with invasion of human intestinal epithelial cells by different enteroinvasive bacterial pathogens. FEMS Immunol. Med. Microbiol. 2004, 40, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, S.; Vlasova, A.N.; Fischer, D.; Kumar, A.; Chattha, K.S.; Rauf, A.; Shao, L.; Langel, S.N.; Rajashekara, G.; Saif, L.J. Differential effects of Escherichia coli nissle and Lactobacillus rhamnosus strain gg on human rotavirus binding, infection, and b cell immunity. J. Immunol. 2016, 196, 1780–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyva Salas, M.; Mounier, J.; Maillard, M.B.; Valence, F.; Coton, E.; Thierry, A. Identification and quantification of natural compounds produced by antifungal bioprotective cultures in dairy products. Food Chem. 2019, 301, 125260. [Google Scholar] [CrossRef] [PubMed]
- Pyar, H.; Peh, K.K.; Liong, M.T. Inhibitory effect of metabolites from probiotics Lactobacillus acidophilus strains on growth of pathogenic bacteria. J. Pharmacol. Toxicol. 2011, 6, 533–540. [Google Scholar] [CrossRef]
- Bhat, M.I.; Singh, V.K.; Sharma, D.; Kapila, S.; Kapila, R. Adherence capability and safety assessment of an indigenous probiotic strain Lactobacillus rhamnosus mtcc-5897. Microb. Pathog. 2019, 130, 120–130. [Google Scholar] [CrossRef]
- Sharma, V.; Ranveer, R.C.; Jain, N.; Aseri, G.K. Bacteriocins: Production, different strategies of purification and applications. Int. J. Res. Pharm. Sci. 2019, 10, 1808–1817. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Wang, S.; Wang, Y.; Chu, X.; Ji, H. Lactobacillus plantarum exhibits antioxidant and cytoprotective activities in porcine intestinal epithelial cells exposed to hydrogen peroxide. Oxidative Med. Cell. Longev. 2021, 2021, 8936907. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Probability (p) of a Null Effect on | |||
|---|---|---|---|---|
| pH of Medium | IPEC-J2 Viability | |||
| LB 1 | E. coli 2 | LB | E. coli | |
| Strain | <0.01 | <0.01 | <0.01 | 0.01 |
| Inoculation level | <0.01 | <0.01 | <0.01 | <0.01 |
| Medium | <0.01 | <0.01 | <0.01 | <0.01 |
| Strain × inoculation level | <0.01 | 0.31 | 0.01 | 0.51 |
| Strain × medium | <0.01 | 0.34 | <0.01 | 0.04 |
| Medium × inoculation level | <0.01 | <0.01 | 0.01 | 0.02 |
| Strain × medium × inoculation level | <0.01 | 0.19 | <0.01 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemay, M.-J.; Raymond, Y.; Champagne, C.P.; Brassard, J. The Growth Medium Affects the Viability of IPEC-J2 Animal Cell Line in the Presence of Probiotic Bacteria. Appl. Microbiol. 2022, 2, 766-781. https://doi.org/10.3390/applmicrobiol2040058
Lemay M-J, Raymond Y, Champagne CP, Brassard J. The Growth Medium Affects the Viability of IPEC-J2 Animal Cell Line in the Presence of Probiotic Bacteria. Applied Microbiology. 2022; 2(4):766-781. https://doi.org/10.3390/applmicrobiol2040058
Chicago/Turabian StyleLemay, Marie-Josée, Yves Raymond, Claude P. Champagne, and Julie Brassard. 2022. "The Growth Medium Affects the Viability of IPEC-J2 Animal Cell Line in the Presence of Probiotic Bacteria" Applied Microbiology 2, no. 4: 766-781. https://doi.org/10.3390/applmicrobiol2040058