Combined Biological and Chemical/Physicochemical Pretreatment Methods of Lignocellulosic Biomass for Bioethanol and Biomethane Energy Production—A Review

 ,
,  , , and
, , and
Abstract
:1. Introduction
1.1. Anaerobic Digestion of Lignocellulosic Biomass
1.2. Bioethanol Production from Lignocellulosic Biomass
2. Comparison of Combined Pretreatment Based on Biogas Yields
3. Comparison of Combined Pretreatment Based on Bioethanol Yields
3.1. Combined Biological–Alkaline Pretreatment
3.2. Combined Biological–Acid Pretreatment
3.3. Combined Biological—Organosolv Pretreatment
3.4. Combined Biological–Steam Explosion Pretreatment
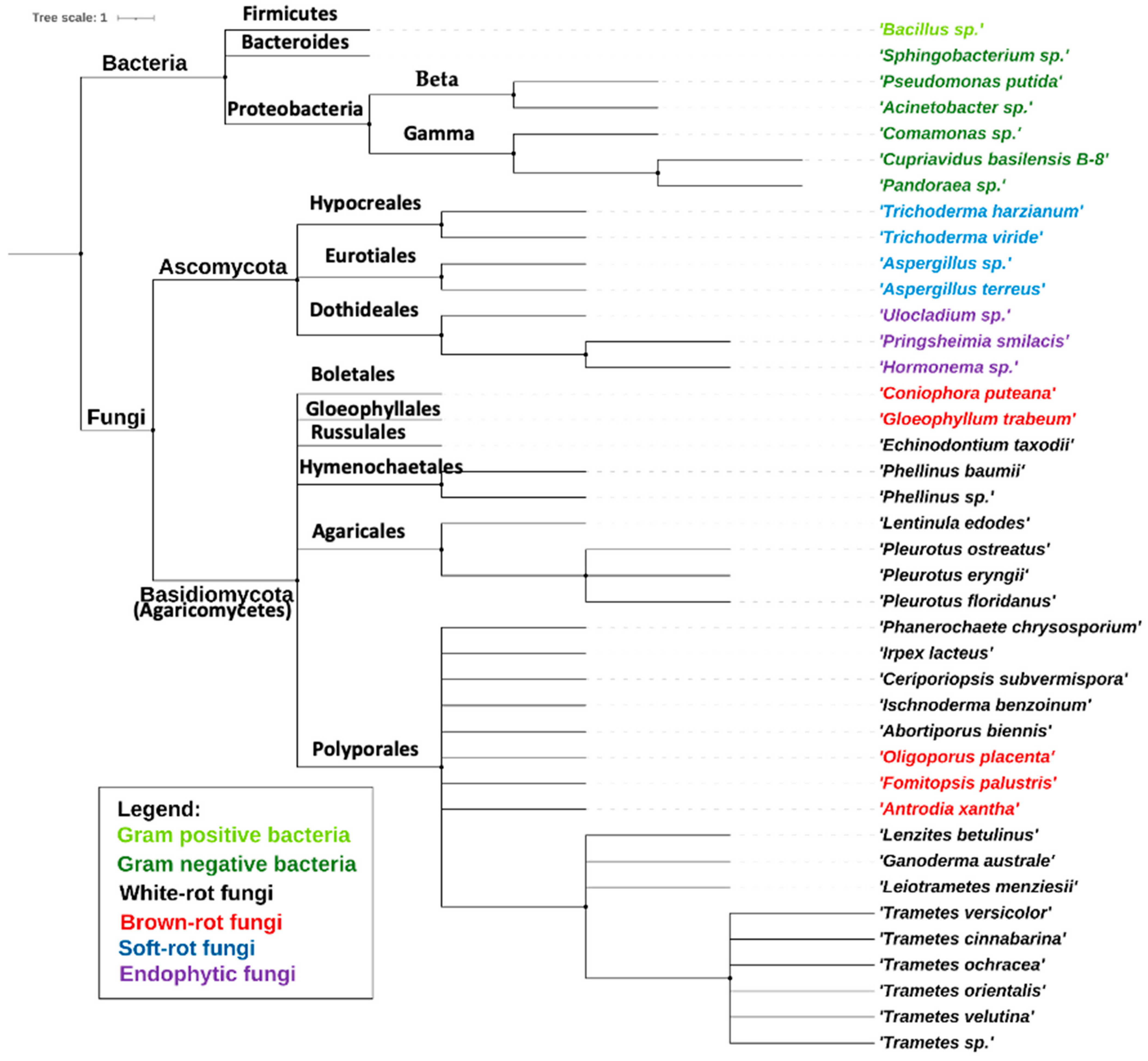
4. Properties of the Employed Microorganisms in the Pretreatment Step
5. New Trends in Microorganisms Used in Downstream Process
6. Challenges and Possible Solutions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Antonopoulou, G.; Kampranis, A.; Ntaikou, I.; Lyberatos, G. Enhancement of Liquid and Gaseous Biofuels Production from Agro-Industrial Residues after Thermochemical and Enzymatic Pretreatment. Front. Sustain. Food Syst. 2019, 3, 92. [Google Scholar] [CrossRef]
- World Bioenergy Association. Global Bioenergy Statistics 2021; World Bioenergy Association: Stockholm, Sweden, 2021; pp. 39–40. [Google Scholar]
- United States Environmental Protection Agency. Economics of Biofuels; EPA: Washington, DC, USA, 2022. [Google Scholar]
- IFP Energies Nouvelles. Biogas and Biomethane: Turning Our Waste into Energy. In Biogas, a Renewable Energy Source. Available online: https://www.ifpenergiesnouvelles.com/issues-and-foresight/decoding-keys/renewable-energies/biogas-and-biomethane-turning-our-waste-energy (accessed on 22 September 2022).
- Hirani, A.H.; Javed, N.; Asif, M.; Basu, S.K.; Kumar, A. A Review on First- and Second-Generation Biofuel Productions. In Biofuels: Greenhouse Gas Mitigation and Global Warming; Kumar, A., Ogita, S., Yau, Y.-Y., Eds.; Springer: New Delhi, India, 2018; pp. 141–154. ISBN 978-81-322-3761-7. [Google Scholar]
- Beig, B.; Riaz, M.; Raza Naqvi, S.; Hassan, M.; Zheng, Z.; Karimi, K.; Pugazhendhi, A.; Atabani, A.E.; Thuy Lan Chi, N. Current Challenges and Innovative Developments in Pretreatment of Lignocellulosic Residues for Biofuel Production: A Review. Fuel 2021, 287, 119670. [Google Scholar] [CrossRef]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Combination of Fungal and Physicochemical Processes for Lignocellulosic Biomass Pretreatment—A Review. Renew. Sustain. Energy Rev. 2016, 54, 217–234. [Google Scholar] [CrossRef]
- Meenakshisundaram, S.; Fayeulle, A.; Leonard, E.; Ceballos, C.; Pauss, A. Fiber Degradation and Carbohydrate Production by Combined Biological and Chemical/Physicochemical Pretreatment Methods of Lignocellulosic Biomass—A Review. Bioresour. Technol. 2021, 331, 125053. [Google Scholar] [CrossRef]
- Abraham, A.; Mathew, A.K.; Park, H.; Choi, O.; Sindhu, R.; Parameswaran, B.; Pandey, A.; Park, J.H.; Sang, B.-I. Pretreatment Strategies for Enhanced Biogas Production from Lignocellulosic Biomass. Bioresour. Technol. 2020, 301, 122725. [Google Scholar] [CrossRef]
- Atelge, M.R.; Krisa, D.; Kumar, G.; Eskicioglu, C.; Nguyen, D.D.; Chang, S.W.; Atabani, A.E.; Al-Muhtaseb, A.H.; Unalan, S. Biogas Production from Organic Waste: Recent Progress and Perspectives. Waste Biomass Valorization 2020, 11, 1019–1040. [Google Scholar] [CrossRef]
- Khan, I.U.; Othman, M.H.D.; Hashim, H.; Matsuura, T.; Ismail, A.F.; Arzhandi, M.R.-D.; Azelee, I.W. Biogas as a Renewable Energy Fuel—A Review of Biogas Upgrading, Utilisation and Storage. Energy Convers. Manag. 2017, 150, 277–294. [Google Scholar] [CrossRef]
- Zion Market Research: Global Biomethane Market Will Reach to USD 2.61 Billion By 2025. Available online: https://www.globenewswire.com/news-release/2019/07/22/1885680/0/en/Global-Biomethane-Market-Will-Reach-to-USD-2-61-Billion-By-2025-Zion-Market-Research.html (accessed on 12 January 2022).
- Paudel, S.R.; Banjara, S.P.; Choi, O.K.; Park, K.Y.; Kim, Y.M.; Lee, J.W. Pretreatment of Agricultural Biomass for Anaerobic Digestion: Current State and Challenges. Bioresour. Technol. 2017, 245, 1194–1205. [Google Scholar] [CrossRef]
- Tabatabaei, M.; Aghbashlo, M.; Valijanian, E.; Kazemi Shariat Panahi, H.; Nizami, A.-S.; Ghanavati, H.; Sulaiman, A.; Mirmohamadsadeghi, S.; Karimi, K. A Comprehensive Review on Recent Biological Innovations to Improve Biogas Production, Part 1: Upstream Strategies. Renew. Energy 2020, 146, 1204–1220. [Google Scholar] [CrossRef]
- Cesaro, A.; Belgiorno, V. Combined Biogas and Bioethanol Production: Opportunities and Challenges for Industrial Application. Energies 2015, 8, 8121–8144. [Google Scholar] [CrossRef]
- Sawatdeenarunat, C.; Surendra, K.C.; Takara, D.; Oechsner, H.; Khanal, S.K. Anaerobic Digestion of Lignocellulosic Biomass: Challenges and Opportunities. Bioresour. Technol. 2015, 178, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Sherrard, A. Breaking the Straw to Biogas Conundrum. Available online: https://bioenergyinternational.com/feedstock/breaking-the-straw-to-biogas-conundrum (accessed on 10 January 2022).
- Ma, S.; Wang, H.; Li, J.; Fu, Y.; Zhu, W. Methane Production Performances of Different Compositions in Lignocellulosic Biomass through Anaerobic Digestion. Energy 2019, 189, 116190. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Xu, F.; Li, Y. Pretreatment of Lignocellulosic Biomass for Enhanced Biogas Production. Prog. Energy Combust. Sci. 2014, 42, 35–53. [Google Scholar] [CrossRef]
- Hagos, K.; Zong, J.; Li, D.; Liu, C.; Lu, X. Anaerobic Co-Digestion Process for Biogas Production: Progress, Challenges and Perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1485–1496. [Google Scholar] [CrossRef]
- Song, Z.; Yang, G.; Liu, X.; Yan, Z.; Yuan, Y.; Liao, Y. Comparison of Seven Chemical Pretreatments of Corn Straw for Improving Methane Yield by Anaerobic Digestion. PLoS ONE 2014, 9, e93801. [Google Scholar] [CrossRef] [PubMed]
- Comino, E.; Rosso, M.; Riggio, V. Investigation of Increasing Organic Loading Rate in the Co-Digestion of Energy Crops and Cow Manure Mix. Bioresour. Technol. 2010, 101, 3013–3019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, G.; Li, W.; Li, C.; Xu, G. Enhanced Biogas Production from Sorghum Stem by Co-Digestion with Cow Manure. Int. J. Hydrogen Energy 2016, 41, 9153–9158. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Sun, Y.; Yuan, Z. Bioaugmentation Strategy for Enhancing Anaerobic Digestion of High C/N Ratio Feedstock with Methanogenic Enrichment Culture. Bioresour. Technol. 2018, 261, 188–195. [Google Scholar] [CrossRef]
- Zieliński, M.; Kisielewska, M.; Dębowski, M.; Elbruda, K. Effects of Nutrients Supplementation on Enhanced Biogas Production from Maize Silage and Cattle Slurry Mixture. Water Air Soil Pollut. 2019, 230, 117. [Google Scholar] [CrossRef]
- Amin, F.R.; Khalid, H.; Zhang, H.; Rahman, S.u.; Zhang, R.; Liu, G.; Chen, C. Pretreatment Methods of Lignocellulosic Biomass for Anaerobic Digestion. AMB Express 2017, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Tesfaw, A.; Assefa, F. Current Trends in Bioethanol Production by Saccharomyces Cerevisiae: Substrate, Inhibitor Reduction, Growth Variables, Coculture, and Immobilization. Int. Sch. Res. Not. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tuong An Tran, T.; Kim Phung Le, T.; Phong Mai, T.; Quan Nguyen, D. Bioethanol Production from Lignocellulosic Biomass. In Alcohol Fuels—Current Technologies and Future Prospect; Yun, Y., Ed.; IntechOpen: London, UK, 2020; ISBN 978-1-78985-043-7. [Google Scholar]
- Zhang, Q.; Wei, Y.; Han, H.; Weng, C. Enhancing Bioethanol Production from Water Hyacinth by New Combined Pretreatment Methods. Bioresour. Technol. 2018, 251, 358–363. [Google Scholar] [CrossRef]
- Li, W.; Khalid, H.; Zhu, Z.; Zhang, R.; Liu, G.; Chen, C.; Thorin, E. Methane Production through Anaerobic Digestion: Participation and Digestion Characteristics of Cellulose, Hemicellulose and Lignin. Appl. Energy 2018, 226, 1219–1228. [Google Scholar] [CrossRef]
- Paul, S.; Dutta, A. Challenges and Opportunities of Lignocellulosic Biomass for Anaerobic Digestion. Resour. Conserv. Recycl. 2018, 130, 164–174. [Google Scholar] [CrossRef]
- Alexandropoulou, M.; Antonopoulou, G.; Fragkou, E.; Ntaikou, I.; Lyberatos, G. Fungal Pretreatment of Willow Sawdust and Its Combination with Alkaline Treatment for Enhancing Biogas Production. J. Environ. Manag. 2017, 203, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Schanbacher, F.L. Production of Methane Biogas as Fuel through Anaerobic Digestion. In Sustainable Biotechnology; Singh, O.V., Harvey, S.P., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 105–127. ISBN 978-90-481-3294-2. [Google Scholar]
- Mao, C.; Feng, Y.; Wang, X.; Ren, G. Review on Research Achievements of Biogas from Anaerobic Digestion. Renew. Sustain. Energy Rev. 2015, 45, 540–555. [Google Scholar] [CrossRef]
- Taherdanak, M.; Zilouei, H. Improving Biogas Production from Wheat Plant Using Alkaline Pretreatment. Fuel 2014, 115, 714–719. [Google Scholar] [CrossRef]
- Al Seadi, T.; Rutz, D.; Prassl, H.; Köttner, M.; Finsterwalder, T.; Volk, S.; Janssen, R. Biogas Handbook; University of Southern Denmark Esbjerg: Esbjerg, Denmark, 2008; ISBN 978-87-992962-0-0. [Google Scholar]
- Yang, D.; Zheng, Y.; Zhang, R. Alkali Pretreatment of Rice Straw for Increasing the Biodegradability. In Proceedings of the 2009 Reno, Nevada, 21 June–24 June 2009; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2009. [Google Scholar]
- He, Y.; Pang, Y.; Liu, Y.; Li, X.; Wang, K. Physicochemical Characterization of Rice Straw Pretreated with Sodium Hydroxide in the Solid State for Enhancing Biogas Production. Energy Fuels 2008, 22, 2775–2781. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Guo, X.M.; Latrille, E.; Trably, E.; Steyer, J.-P.; Carrere, H. Predictive Models of Biohydrogen and Biomethane Production Based on the Compositional and Structural Features of Lignocellulosic Materials. Environ. Sci. Technol. 2012, 46, 12217–12225. [Google Scholar] [CrossRef]
- Amirta, R.; Tanabe, T.; Watanabe, T.; Honda, Y.; Kuwahara, M.; Watanabe, T. Methane Fermentation of Japanese Cedar Wood Pretreated with a White Rot Fungus, Ceriporiopsis Subvermispora. J. Biotechnol. 2006, 123, 71–77. [Google Scholar] [CrossRef]
- Lalak, J.; Kasprzycka, A.; Martyniak, D.; Tys, J. Effect of Biological Pretreatment of Agropyron Elongatum ‘BAMAR’ on Biogas Production by Anaerobic Digestion. Bioresour. Technol. 2016, 200, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Mackuľak, T.; Prousek, J.; Švorc, Ľ.; Drtil, M. Increase of Biogas Production from Pretreated Hay and Leaves Using Wood-Rotting Fungi. Chem. Pap. 2012, 66, 649–653. [Google Scholar] [CrossRef]
- Ali, S.S.; Sun, J. Physico-Chemical Pretreatment and Fungal Biotreatment for Park Wastes and Cattle Dung for Biogas Production. SpringerPlus 2015, 4, 712. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Luo, K.; Zhang, Y.; Zheng, Z.; Cai, Y.; Wen, B.; Cui, Z.; Wang, X. Improving the Methane Yield of Maize Straw: Focus on the Effects of Pretreatment with Fungi and Their Secreted Enzymes Combined with Sodium Hydroxide. Bioresour. Technol. 2018, 250, 204–213. [Google Scholar] [CrossRef]
- Scharer, J.M.; Moo-Young, M. Methane Generation by Anaerobic Digestion of Cellulose-Containing Wastes. Adv. Biochem. Eng. 1979, 11, 85–101. [Google Scholar]
- Monrroy, M.; Ibañez, J.; Melin, V.; Baeza, J.; Mendonça, R.T.; Contreras, D.; Freer, J. Bioorganosolv Pretreatments of P. Radiata by a Brown Rot Fungus (Gloephyllum Trabeum) and Ethanolysis. Enzyme Microb. Technol. 2010, 47, 11–16. [Google Scholar] [CrossRef]
- Muñoz, C.; Mendonça, R.; Baeza, J.; Berlin, A.; Saddler, J.; Freer, J. Bioethanol Production from Bio- Organosolv Pulps OfPinus Radiata AndAcacia Dealbata. J. Chem. Technol. Biotechnol. 2007, 82, 767–774. [Google Scholar] [CrossRef]
- Fissore, A.; Carrasco, L.; Reyes, P.; Rodríguez, J.; Freer, J.; Mendonça, R.T. Evaluation of a Combined Brown Rot Decay–Chemical Delignification Process as a Pretreatment for Bioethanol Production from Pinus Radiata Wood Chips. J. Ind. Microbiol. Biotechnol. 2010, 37, 893–900. [Google Scholar] [CrossRef]
- Salvachúa, D.; Prieto, A.; López-Abelairas, M.; Lu-Chau, T.; Martínez, Á.T.; Martínez, M.J. Fungal Pretreatment: An Alternative in Second-Generation Ethanol from Wheat Straw. Bioresour. Technol. 2011, 102, 7500–7506. [Google Scholar] [CrossRef]
- Ma, F.; Yang, N.; Xu, C.; Yu, H.; Wu, J.; Zhang, X. Combination of Biological Pretreatment with Mild Acid Pretreatment for Enzymatic Hydrolysis and Ethanol Production from Water Hyacinth. Bioresour. Technol. 2010, 101, 9600–9604. [Google Scholar] [CrossRef]
- Gui, X.; Wang, G.; Hu, M.; Yan, Y. Combined Fungal and Mild Acid Pretreatment of Glycyrrhiza Uralensis Residue for Enhancing Enzymatic Hydrolysis and Oil Production. BioResources 2013, 8, 5485–5499. [Google Scholar] [CrossRef]
- Gui, X.; Wang, G.; Li, X.; Yan, Y. Fungus-Assisted Mild Acid Pretreatment of Glycyrrhiza Uralensis Residues to Enhance Enzymatic Hydrolysis and Oil Production by Green Microalga Chlorella Protothecoides. Ind. Crops Prod. 2014, 62, 466–473. [Google Scholar] [CrossRef]
- Ishola, M.M.; Isroi; Taherzadeh, M.J. Effect of Fungal and Phosphoric Acid Pretreatment on Ethanol Production from Oil Palm Empty Fruit Bunches (OPEFB). Bioresour. Technol. 2014, 165, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Si, M.; Liu, D.; Zhuo, S.; Liu, M.; Liu, H.; Yan, X.; Shi, Y. A Bionic System with Fenton Reaction and Bacteria as a Model for Bioprocessing Lignocellulosic Biomass. Biotechnol. Biofuels 2018, 11, 31. [Google Scholar] [CrossRef]
- Itoh, H.; Wada, M.; Honda, Y.; Kuwahara, M.; Watanabe, T. Bioorganosolve Pretreatments for Simultaneous Saccharification and Fermentation of Beech Wood by Ethanolysis and White Rot Fungi. J. Biotechnol. 2003, 103, 273–280. [Google Scholar] [CrossRef]
- Baba, Y.; Tanabe, T.; Shirai, N.; Watanabe, T.; Honda, Y.; Watanabe, T. Pretreatment of Japanese Cedar Wood by White Rot Fungi and Ethanolysis for Bioethanol Production. Biomass Bioenergy 2011, 35, 320–324. [Google Scholar] [CrossRef]
- Asada, C.; Asakawa, A.; Sasaki, C.; Nakamura, Y. Characterization of the Steam-Exploded Spent Shiitake Mushroom Medium and Its Efficient Conversion to Ethanol. Bioresour. Technol. 2011, 102, 10052–10056. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin Degradation: Microorganisms, Enzymes Involved, Genomes Analysis and Evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products. Int. J. Environ. Res. Public. Health 2021, 18, 6001. [Google Scholar] [CrossRef]
- Yang, H.; Wang, K.; Wang, W.; Sun, R.-C. Improved Bioconversion of Poplar by Synergistic Treatments with White-Rot Fungus Trametes Velutina D10149 Pretreatment and Alkaline Fractionation. Bioresour. Technol. 2013, 130, 578–583. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Wróbel, B.; Górska, E.B. Cellulolytic Properties of a Potentially Lignocellulose-Degrading Bacillus Sp. 8E1A Strain Isolated from Bulk Soil. Agronomy 2022, 12, 665. [Google Scholar] [CrossRef]
- Zhuo, S.; Yan, X.; Liu, D.; Si, M.; Zhang, K.; Liu, M.; Peng, B.; Shi, Y. Use of Bacteria for Improving the Lignocellulose Biorefinery Process: Importance of Pre-Erosion. Biotechnol. Biofuels 2018, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose Degradation: An Overview of Fungi and Fungal Enzymes Involved in Lignocellulose Degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Strobel, G. The Emergence of Endophytic Microbes and Their Biological Promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Krah, F.-S.; Bässler, C.; Heibl, C.; Soghigian, J.; Schaefer, H.; Hibbett, D.S. Evolutionary Dynamics of Host Specialization in Wood-Decay Fungi. BMC Evol. Biol. 2018, 18, 119. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Valášková, V. Degradation of Cellulose by Basidiomycetous Fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Li, Y. Fungal Pretreatment of Lignocellulosic Biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef]
- Wei, S. The Application of Biotechnology on the Enhancing of Biogas Production from Lignocellulosic Waste. Appl. Microbiol. Biotechnol. 2016, 100, 9821–9836. [Google Scholar] [CrossRef]
- Dollhofer, V.; Podmirseg, S.M.; Callaghan, T.M.; Griffith, G.W.; Fliegerová, K. Anaerobic Fungi and Their Potential for Biogas Production. In Biogas Science and Technology; Guebitz, G.M., Bauer, A., Bochmann, G., Gronauer, A., Weiss, S., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Switzerland, 2015; Volume 151, pp. 41–61. ISBN 978-3-319-21992-9. [Google Scholar]
- Cheng, Y.F.; Edwards, J.E.; Allison, G.G.; Zhu, W.-Y.; Theodorou, M.K. Diversity and Activity of Enriched Ruminal Cultures of Anaerobic Fungi and Methanogens Grown Together on Lignocellulose in Consecutive Batch Culture. Bioresour. Technol. 2009, 100, 4821–4828. [Google Scholar] [CrossRef]
- Jin, W.; Cheng, Y.-F.; Mao, S.-Y.; Zhu, W.-Y. Isolation of Natural Cultures of Anaerobic Fungi and Indigenously Associated Methanogens from Herbivores and Their Bioconversion of Lignocellulosic Materials to Methane. Bioresour. Technol. 2011, 102, 7925–7931. [Google Scholar] [CrossRef]
- Procházka, J.; Dolejš, P.; Máca, J.; Dohányos, M. Stability and Inhibition of Anaerobic Processes Caused by Insufficiency or Excess of Ammonia Nitrogen. Appl. Microbiol. Biotechnol. 2012, 93, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Ferdeș, M.; Dincă, M.N.; Moiceanu, G.; Zăbavă, B.Ș.; Paraschiv, G. Microorganisms and Enzymes Used in the Biological Pretreatment of the Substrate to Enhance Biogas Production: A Review. Sustainability 2020, 12, 7205. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wang, F.; Shi, X.-S.; Guo, R.-B. Impacts of Microaeration on the Anaerobic Digestion of Corn Straw and the Microbial Community Structure. Chem. Eng. J. 2016, 287, 523–528. [Google Scholar] [CrossRef]
- Hu, Y.; Hao, X.; Wang, J.; Cao, Y. Enhancing Anaerobic Digestion of Lignocellulosic Materials in Excess Sludge by Bioaugmentation and Pre-Treatment. Waste Manag. 2016, 49, 55–63. [Google Scholar] [CrossRef]
- Mulat, D.G.; Huerta, S.G.; Kalyani, D.; Horn, S.J. Enhancing Methane Production from Lignocellulosic Biomass by Combined Steam-Explosion Pretreatment and Bioaugmentation with Cellulolytic Bacterium Caldicellulosiruptor Bescii. Biotechnol. Biofuels 2018, 11, 19. [Google Scholar] [CrossRef]
- Sträuber, H.; Bühligen, F.; Kleinsteuber, S.; Nikolausz, M.; Porsch, K. Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms. Bioengineering 2015, 2, 66–93. [Google Scholar] [CrossRef]
- Sarkar, N.; Ghosh, S.K.; Bannerjee, S.; Aikat, K. Bioethanol Production from Agricultural Wastes: An Overview. Renew. Energy 2012, 37, 19–27. [Google Scholar] [CrossRef]
- Meethit, P.; Ratanaprasit, P.; Sakdaronnarong, C. Candida Shehatae and Saccharomyces Cerevisiae Work Synergistically to Improve Ethanol Fermentation from Sugarcane Bagasse and Rice Straw Hydrolysate in Immobilized Cell Bioreactor. Eng. Life Sci. 2016, 16, 706–719. [Google Scholar] [CrossRef]
- Kun, R.S.; Gomes, A.C.S.; Hildén, K.S.; Salazar Cerezo, S.; Mäkelä, M.R.; de Vries, R.P. Developments and Opportunities in Fungal Strain Engineering for the Production of Novel Enzymes and Enzyme Cocktails for Plant Biomass Degradation. Biotechnol. Adv. 2019, 37, 107361. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Sankaran, R.; Show, P.L.; Ibrahim, T.N.B.T.; Chew, K.W.; Culaba, A.; Jo-Shu, C. Pretreatment Methods for Lignocellulosic Biofuels Production: Current Advances, Challenges and Future Prospects. Biofuel Res. J. 2020, 7, 1115–1127. [Google Scholar] [CrossRef]
- Zhou, M.; Tian, X. Development of Different Pretreatments and Related Technologies for Efficient Biomass Conversion of Lignocellulose. Int. J. Biol. Macromol. 2022, 202, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Pham, L.T.M.; Choudhary, H.; Gauttam, R.; Singer, S.W.; Gladden, J.M.; Simmons, B.A.; Singh, S.; Sale, K.L. Revisiting Theoretical Tools and Approaches for the Valorization of Recalcitrant Lignocellulosic Biomass to Value-Added Chemicals. Front. Energy Res. 2022, 10, 863153. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Substrate | Step 1 | Step 2 | % Increase Compared to Sole Biological Pretreatment | Methane Yield |
|---|---|---|---|---|---|
| Ali and Sun (2015) [43] | Park waste (dry and fresh leaves) + cattle manure | 2.5 % NaOH and 2.5% NH4OH (15 d) | Aspergillus terreus and Trichoderma viride (25 °C, 7 d) | 30 (compared to untreated biomass) | 79.8 L/kgVS (125.9 L/kgVS biogas production) |
| Alexandropoulou et al. (2017) [32] | Willow sawdust | Leiotrametes menziesii (27 °C, 30 d) | 1% (w/v) NaOH (80 °C, 24 h) | 48.9 | 142.2 ± 0.3 L/kg TS (L. menziesii) |
| Abortiporus biennis (27 °C, 30 d) | 50.1 | 205.3 ± 0.3 L /kg TS (A. biennis) | |||
| Zhao et al. (2018) [44] | Maize straw | 1% (w/v) NaOH (room temperature, 48 h) | Aspergillus sp. (30 °C, 10 d) | 13.34 * | 276.29 L/kg TS |
| T. harzianum (30 °C, 10 d) | 22.88 * | 261.63 L/kg TS | |||
| T. harzianum & Aspergillus sp. (30 °C, 10 d) | 31.77 * | 277.99 L/kg TS | |||
| Enzyme from Aspergillus sp. (KY644131) (50 °C, 10 d) | 25.02 * | 300.85 L/kg TS | |||
| Enzyme from T. harzianum (KY644130) (50 °C, 10 d) | 4.14 * | 285.09 L/kg TS | |||
| Enzyme from T. harzianum & Aspergillus sp. (50 °C, 10 d) | −6.71 * (decrease) | 258.45 L/kg TS |
| Substrate | 1st Step | 2nd Step | Hydrolysis and Fermentation (Ethanol Concentration Detection Technique) | Ethanol Yield Increase (%) | Ethanol Concentration (% Theoretical Ethanol Yield) | Reference |
|---|---|---|---|---|---|---|
| Biological—Alkaline Pretreatment | ||||||
| Pinus Radiata | Gloeophyllum trabeum (27 °C, 28 d) | 25% w/w NaOH (180 °C, 5 h) | SHF: 5% substrate consistency. Celluclast (20 FPU/g) and β-glucosidase (40 UI/g) (50 °C, 24 h, 150 rpm). 3 g/L Saccharomyces cerevisiae IR2-9a (40 °C, 96 h, 150 rpm). (GC-FID) | −37.5% * compared to alkaline alone | 99.55 mL/kg wood (33.98%) | Fissore et al. (2010) [48] |
| Wheat straw (Triticum aestivum) | P. subvermispora (28 °C, 21 d) | 0.1% NaOH (5% w/v) (50 °C, 1 h, 165 rpm) | SHF: Cellulase (15 FPU/g) and xylanase (30 U/g) (50 °C, 60 h, 165 rpm). 0.5 g/L Saccharomyces cerevisiae (Fermentis LPA 3035) (32 °C, 24 h, 200 rpm). (GC-FID) | 79.41% * compared to alkaline alone | 122 ± 8 mg/g substrate (62%) | Salvachúa et al. (2011) [49] |
| I. lacteus (28 °C, 21 d) | 80.88% * compared to alkaline alone | 123 ± 5 mg/g substrate (62%) | ||||
| Biological—Acid Pretreatment | ||||||
| Water hyacinth (Eichhornia crassipes) | Echinodontium Taxodii (28 °C, 10 d) | 0.25% H2SO4 (100 °C, 1 h) | SHF: 2% substrate consistency; Cellulase (30 FPU/g) (50 °C, 48 h). 0.3% v/v activated Saccharomyces cerevisiae (40 °C, 72 h, 100 rpm for first 8 h). (HPLC) | 31.51% * (1.34-fold increase) compared to acid alone | 0.192 g/g of dry material (sole acid = 0.146 g/g of dry material) | Ma et al. (2010) [50] |
| Glycyrrhiza uralensis Fisch. Ex DC (GUR) | Phanerochaete chrysosporium (28 °C, 21 d) | 2.5% H2SO4 (100 °C, 2.5 h) | Cellulase (30 FPU/g) (50 °C, 48 h); heterotrophic cultivation of C. protothecoides for microalgal oil production. (28 °C, 7 d, 200 rpm) (GC-MS) | 1.34-fold increase relative to acid treatment alone | 1.66 g/L (oil content) | Gui et al. (2013) [51] |
| Glycyrrhiza uralensis Fisch. Ex DC (GUR) | Phanerochaete chrysosporium (28 °C, 21 d) | 2M acetic acid (100 °C, 3 h) | Cellulase (40 FPU/g) (50 °C, 48 h); heterotrophic cultivation of C. protothecoides (28 °C, 120 h, 200 rpm) (GC-MS) | 1.54 *-fold increase relative to acid treatment alone | 1.91 g/L (oil content) | Gui et al. (2014) [52] |
| Oil palm empty fruit bunches (OPEFB) | Pleurotus floridanus LIPIMC996 (31 °C, 28 d) | Ball-milled at 29.6/s for 4 min; phosphoric acid treatment (50 °C, 5 h) | SSF: Enzymatic hydrolysis (60 FPU/g cellulose). Saccharomyces cerevisiae CBS 8066 (35 °C, 48 h, 130 rpm) (HPLC) | 7.39% * increase relative to acid treatment alone | 21.8 g/L (62.8%) | Ishola et al. (2014) [53] |
| Water hyacinth | Phanerochaete chrysosporium (30 °C, 60 h, 150 rpm) | 1% H2SO4 (100 °C, 1 h) | 6 g/L Saccharomyces cerevisiae (30 °C, 24 h, 120 rpm) (GC) | 8.61% increase relative to acid treatment alone | 1.40 g/L | Zhang et al. (2018) [54] |
| Biological—Organosolv Pretreatment | ||||||
| Sapwood of beech (Fagus crenata) | C. subvermispora FP90031 (28 °C, 56 d) | 60% (v/v) ethanol solution (180 °C, 2 h) | SSF: Cellulase (10 FPU/0.25 g). 10% v/v S. cerevisiae AM12 (40 °C, 96 h, 100 rpm) (GC-FID) | 1.6-fold increase relative to ethanolysis treatment alone | 0.176 g/g of wood | Itoh et al. (2003) [55] |
| Pinus radiata wood chips | Ceriporiopsis subvermispora (27 °C, 30 d) | 60% ethanol in water solvent (200 °C, 1 h) (H-factor: 11,360); cold alkaline wash: 1% NaOH for 10 min; hot alkaline wash: 1% NaOH (75 °C, 1 h) | SHF: Cellulase (20 FPU/g glucan) and β-glucosidase (40 CBU/g glucan) (50 °C, 72 h, 150 rpm); Saccharomyces cerevisiae Y-1528 (30 °C, 48 h, 150 rpm) (GC-FID); SSF: 2% substrate consistency; Cellulase (20 FPU/g glucan) and β-glucosidase (40 CBU/g glucan) (50 °C, 72 h, 150 rpm). Saccharomyces cerevisiae Y-1528 (37 °C, 48 h, 150 rpm) (GC-FID) | SHF: 61.90% *; SSF: 458% * compared to control | SHF: 136 g/kg wood (37%) SSF: 162 g/kg wood (44%) | Muñoz et al. (2007) [47] |
| Acacia dealbata wood chips | Ganoderma australe (27 °C, 30 d) | 60% ethanol in water solvent (200 °C, 1 h) (H-factor: 10,920); cold alkaline wash: 1% NaOH for 10 min; hot alkaline wash: 1% NaOH (75 °C, 1 h) | SHF: Cellulase (20 FPU/g glucan) and β-glucosidase (40 CBU/g glucan) (50 °C, 72 h, 150 rpm); Saccharomyces cerevisiae Y-1528 (30 °C, 48 h, 150 rpm) (GC-FID); SSF: 2% substrate consistency; Cellulase (20 FPU/g glucan) and β-glucosidase (40 CBU/g glucan). Saccharomyces cerevisiae Y-1528 (37 °C, 48 h, 150 rpm) (GC-FID) | SHF: −7.14% *; SSF: 4.28% * compared to control | SHF: 143 g/kg wood (48%) SSF: 195 g/kg wood (65%) | Muñoz et al. (2007) [47] |
| Pinus radiata | Gloeophyllum trabeum (27 °C, 28 d) | 60% ethanol in water solvent (200 °C, 1 h) | SSF: Celluclast (20 FPU/g) and β-glucosidase (40 UI/g) (50 °C, 24 h, 150 rpm); 3 g/L Saccharomyces cerevisiae IR2-9a (40 °C, 96 h, 150 rpm) (GC-FID) | 44.6% | 210 mL/kg wood (72%) | Fissore et al. (2010) [48] |
| Pinus radiata wood chips | Gloephyllum trabeum ATCC 11539 (25 °C, 21 d) | Biopulp: 95% ethanol in water solvent (60:40 v/v ratio) with 0.13% H2SO4 (w/v) (185 °C, 18 min) Control pulp: 95% ethanol in water solvent (60:40 v/v ratio) with 0.13% H2SO4 (w/v) (200 °C, 32 min) | SSF: 10% substrate consistency; Celluclast (20 FPU/g), β-glucosidase Novozymes 188 (40 IU/g); 6.0 g/L Saccharomyces cerevisiae IR2T9 (40 °C, 96 h, 150 rpm) (GC-FID) | Similar yield in both control pulp and biopulp | 161 g/kg wood (63.8%) | Monrroy et al. (2010) [46] |
| Japanese cedar (Cryptomeria japonica) | Phellinus sp. SKM2102 (28 °C, 56 d) | Ethanol/lactic acid/water (40:10:50, w/w) (190 °C, 30 min) | SSF: Meicelase (10 FPU/0.25 g); 10% v/v S. cerevisiae AM12 (35 °C, 72 h, 100 rpm) (GC-FID) | NA | 8.94 g/L (28.3%) | Baba et al. (2011) [56] |
| C. subvermispora FP-90031-sp (28 °C, 56 d) | Ethanol/lactic acid/water (40:10:50, w/w) (200 °C, 1 h) | 9.82 g/L (31.1%) | ||||
| Biological—Steam Explosion Pretreatment | ||||||
| Sawtooth oak, corn, and bran | Lentinula edodes (120 d) | Steam explosion (214 °C, 5 min, 20 atm) | SSF: 0.1 g enzyme/g substrate Meicelase (45 °C, 48 h, 140 strokes/min); Saccharomyces cerevisiae AM 12 (40 °C, 24 h, 100 rpm) (HPLC) | 49.68% * increase compared to spent shitake mushroom medium | 23.8 g/L (87.6%) | Asada et al. (2011) [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meenakshisundaram, S.; Fayeulle, A.; Léonard, E.; Ceballos, C.; Liu, X.; Pauss, A. Combined Biological and Chemical/Physicochemical Pretreatment Methods of Lignocellulosic Biomass for Bioethanol and Biomethane Energy Production—A Review. Appl. Microbiol. 2022, 2, 716-734. https://doi.org/10.3390/applmicrobiol2040055
Meenakshisundaram S, Fayeulle A, Léonard E, Ceballos C, Liu X, Pauss A. Combined Biological and Chemical/Physicochemical Pretreatment Methods of Lignocellulosic Biomass for Bioethanol and Biomethane Energy Production—A Review. Applied Microbiology. 2022; 2(4):716-734. https://doi.org/10.3390/applmicrobiol2040055
Chicago/Turabian StyleMeenakshisundaram, Shruthi, Antoine Fayeulle, Estelle Léonard, Claire Ceballos, Xiaojun Liu, and André Pauss. 2022. "Combined Biological and Chemical/Physicochemical Pretreatment Methods of Lignocellulosic Biomass for Bioethanol and Biomethane Energy Production—A Review" Applied Microbiology 2, no. 4: 716-734. https://doi.org/10.3390/applmicrobiol2040055