
Basic Residue Clusters in Intrinsically Disordered Regions of Peripheral Membrane Proteins: Modulating 2D Diffusion on Cell Membranes


Abstract
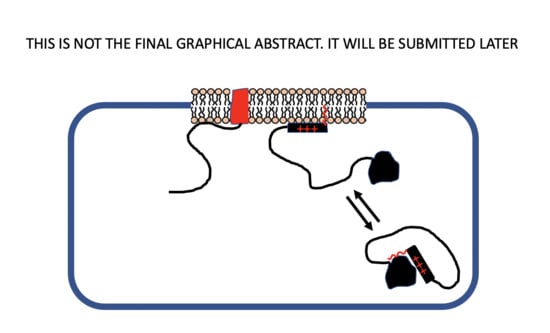
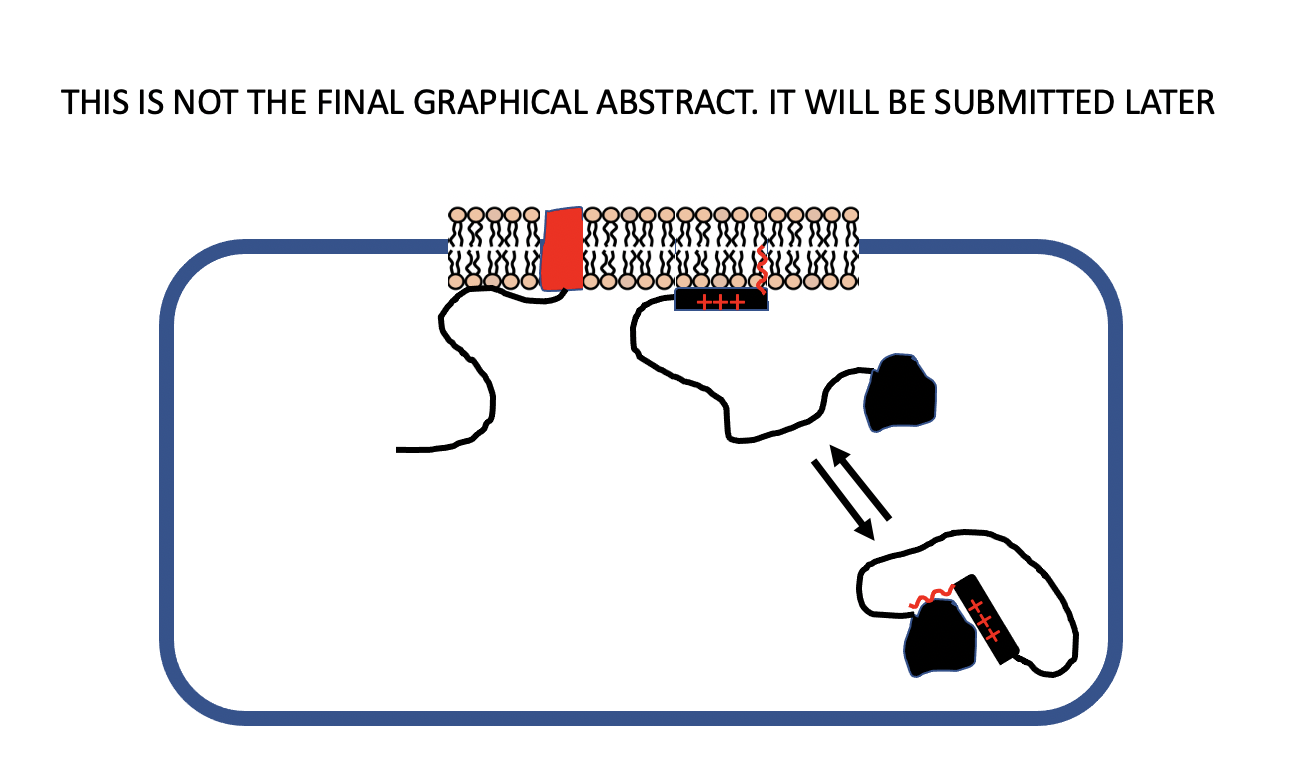
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Peripheral Membrane Proteins
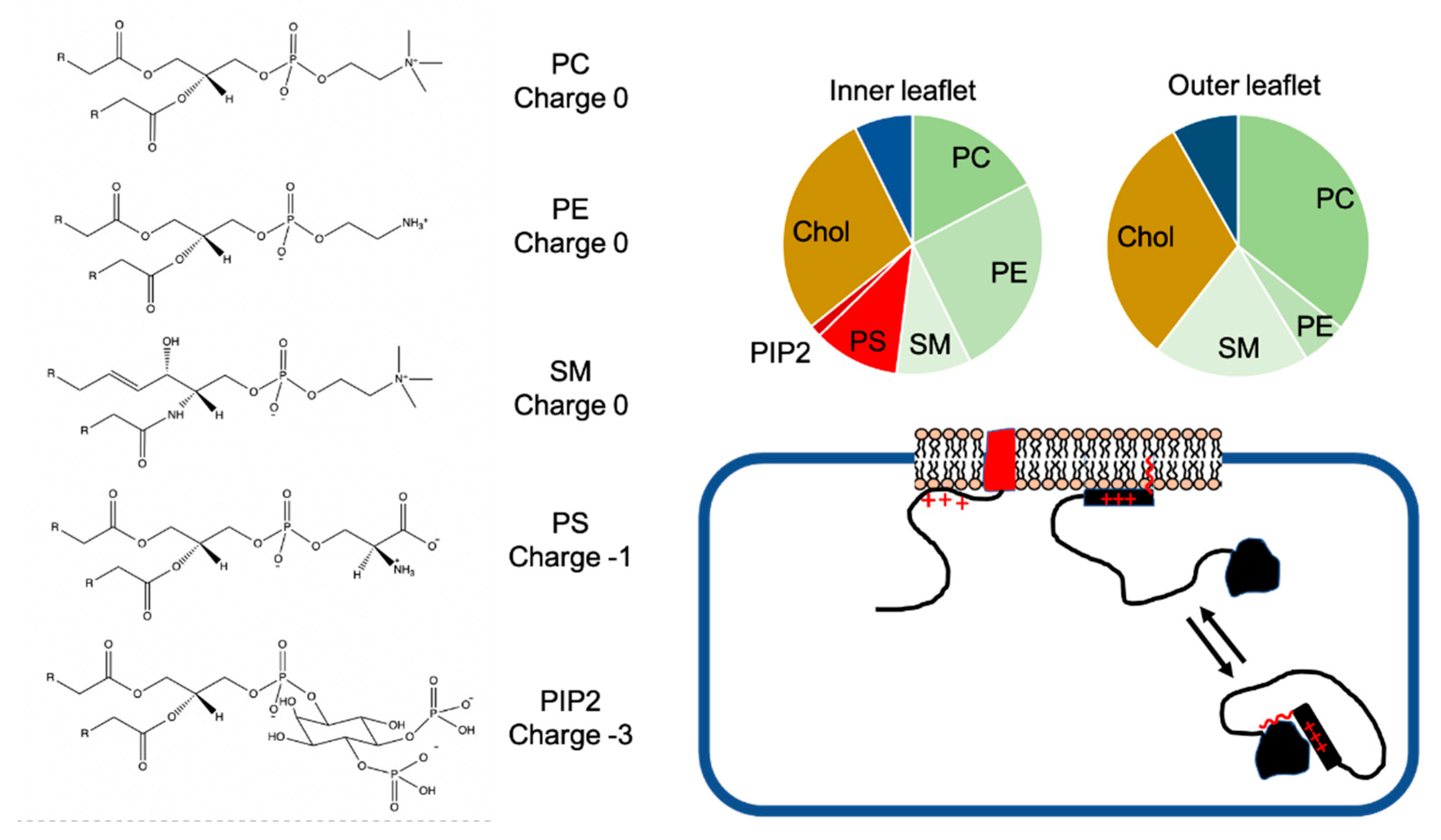
3. Asymmetric Distribution of Charged Lipids in Membranes
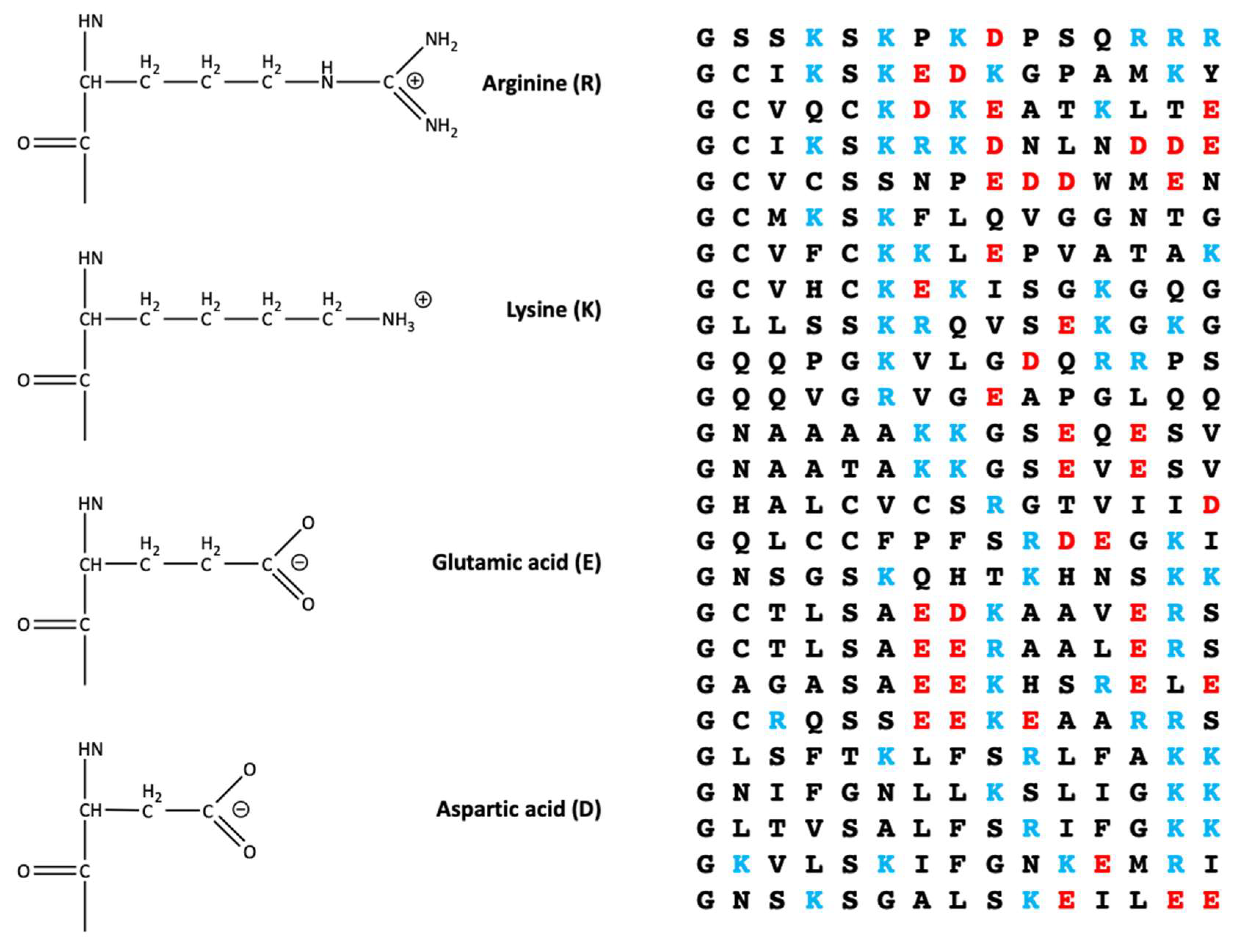
4. Positively Charged Disordered Regions in the Vicinity of Lipid Anchors Can Arise from Basic Residue Clusters or Depletion of Acidic Residues
5. Other Roles of Positively Charged Disordered Regions Bound to Membranes: PIP Modulation
6. Self-Association of Peripheral Membrane Proteins Involving Basic Residue Clusters
7. 3D versus 2D
- (i)
- Local concentration: the same number of proteins distributed uniformly in the interior of the cell or restricted to the narrow shell around the membrane would result in a much larger concentration in the vicinity of the membrane. If we assume a cell with a radius of 10 mm and a submembrane layer of around 3 nm, the increase in concentration is a factor of 1000. The increased concentrations affect the association-dissociation equilibrium with other partners or that of self-association and also affects the diffusion-driven mechanism of such encounters.
- (ii)
- As first suggested by Adam and Delbrück [42], reduction of dimensionality is expected to reduce the time between random encounters between two species diffusing in 2D. A similar argument has been used to explain how the high efficiency transcription factors find their target sequences in DNA using 1D searches [43].
- (iii)
- Anisotropic interactions: a non-spherical intracellular domain physically tethered to the membrane surface or experiencing strong anisotropic interactions (e.g., electrostatic) may become optimally oriented to present complementary contact points to other membrane anchored components. An example is provided by the formation of antiparallel b-sheet by peptides with alternating basic and neutral residues interacting electrostatically with anionic lipids [34].
8. Crowding and Restricted 2D Diffusion. Can Be Faster?
9. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Symons, J.; Cho, K.-J.; Chang, J.T.; Du, G.; Waxham, M.N.; Hancock, J.F.; Levental, I.; Levental, K.R. Lipidomic atlas of mammalian cell membranes reveals hierarchical variation induced by culture conditions, subcellular membranes, and cell lineages. Soft Matter 2021, 17, 288–297. [Google Scholar] [CrossRef]
- Mohammad, I.-L.; Mateos, B.; Pons, M. The disordered boundary of the cell: Emerging properties of membrane-bound intrinsically disordered proteins. Biomol. Concepts 2019, 10, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Weiss, M.; Simonovic, M.; Haertinger, G.; Schrimpf, S.P.; Hengartner, M.O.; von Mering, C. PaxDb, a database of protein abundance averages across all three domains of life. Mol. Cell. Proteom. 2012, 11, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornish, J.; Chamberlain, S.G.; Owen, D.; Mott, H.R. Intrinsically disordered proteins and membranes: A marriage of convenience for cell signalling? Biochem. Soc. Trans. 2020, 48, 2669–2689. [Google Scholar] [CrossRef] [PubMed]
- Burn, P. Talking point Amphitropic proteins: A new class of membrane proteins. Trends Biochem. Sci. 1988, 13, 79–83. [Google Scholar] [CrossRef]
- Nastou, K.; Tsaousis, G.N.; Kremizas, K.E.; Litou, Z.I.; Hamodrakas, S.J. The human plasma membrane peripherome: Visualization and analysis of interactions. BioMed Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lomize, M.A.; Lomize, A.L.; Pogozheva, I.D.; Mosberg, H.I. OPM: Orientations of proteins in membranes database. Bioinformatics 2006, 22, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.N.; Entova, S.; Ray, L.C.; Imperiali, B. Monotopic Membrane Proteins Join the Fold. Trends Biochem. Sci. 2019, 44, 7–20. [Google Scholar] [CrossRef]
- Resh, M.D. Trafficking and signaling by fatty-acylated and prenylated proteins. Nat. Chem. Biol. 2006, 2, 584–590. [Google Scholar] [CrossRef]
- Mosior, M.; McLaughlin, S. Binding of Basic Peptides to Acidic Lipids in Membranes: Effects of Inserting Alanine(s) Between the Basic Residues. Biochemistry 1992, 31, 1767–1773. [Google Scholar] [CrossRef]
- McLaughlin, S. The electrostatic properties of membranes. Annu. Rev. Biophys. Biophys. Chem. 1989, 18, 113–136. [Google Scholar] [CrossRef]
- Balach, M.M.; Casale, C.H.; Campetelli, A.N. Erythrocyte plasma membrane potential: Past and current methods for its measurement. Biophys. Rev. 2019, 11, 995–1005. [Google Scholar] [CrossRef]
- Gurtovenko, A.A.; Vattulainen, I. Lipid transmembrane asymmetry and intrinsic membrane potential: Two sides of the same coin. J. Am. Chem. Soc. 2007, 129, 5358–5359. [Google Scholar] [CrossRef] [PubMed]
- Gairí, M.; Romi, R.; Fernández, I.; Rochat, H.; Martin-Eauclaire, M.F.; Van Rietschoten, J.; Pons, M.; Giralt, E. 3D structure of kaliotoxin: Is residue 34 a key for channel selectivity? J. Pept. Sci. 1997, 3, 314–319. [Google Scholar] [CrossRef]
- Weber, D.; Yao, S.; Rojko, N.; Anderluh, G.; Lybrand, T.P.; Downton, M.T.; Wagner, J.; Separovic, F. Characterization of the lipid-binding site of equinatoxin II by NMR and molecular dynamics simulation. Biophys. J. 2015, 108, 1987–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.M.; He, T.; Fuglebakk, E.; Grauffel, C.; Yang, B.; Roberts, M.F.; Gershenson, A.; Reuter, N. A Role for Weak Electrostatic Interactions in Peripheral Membrane Protein Binding. Biophys. J. 2016, 110, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Branduardi, D.; Gervasio, F.L.; Cavalli, A.; Recanatini, A.M.; Parrinello, M. The role of the peripheral anionic site and cation−π interactions in the ligand penetration of the human AChE gorge. J. Am. Chem. Soc. 2005, 127, 9147–9155. [Google Scholar] [CrossRef]
- Zhenyu, L.; Lai, J.; Yingkai, Z. Importance of charge independent effects in readout of the trimethyllysine mark by HP1 chromodomain. J. Am. Chem. Soc. 2009, 131, 14928–14931. [Google Scholar]
- Gianni, S.; Freiberger, M.I.; Jemth, P.; Ferreiro, D.U.; Wolynes, P.G.; Fuxreiter, M. Fuzziness and Frustration in the Energy Landscape of Protein Folding, Function, and Assembly. Acc. Chem. Res. 2021, 54, 1251–1259. [Google Scholar] [CrossRef]
- McLaughlin, S.; Aderem, A. The myristoyl-electrostatic switch: A modulator of reversible protein-membrane interactions. Trends Biochem. Sci. 1995, 20, 272–276. [Google Scholar] [CrossRef]
- Kim, J.; Mosior, M.; Chung, L.; Wu, H.; McLaughlin, S. Binding of peptides with basic residues to membranes containing acidic phospholipids. Biophys. J. 1991, 60, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Ben-Tal, N.; Honig, B.; Peitzsch, R.; Denisov, G.; McLaughlin, S. Binding of small basic peptides to membranes containing acidic lipids: Theoretical models and experimental results. Biophys. J. 1996, 71, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Deryusheva, E.; Nemashkalova, E.; Galloux, M.; Richard, C.-A.; Eléouët, J.-F.; Kovacs, D.; Van Belle, K.; Tompa, P.; Uversky, V.N.; Permyakov, S. Does Intrinsic Disorder in Proteins Favor Their Interaction with Lipids? Proteom. 2019, 19, e1800098. [Google Scholar] [CrossRef]
- Piovesan, D.; Tabaro, F.; Mičetić, I.; Necci, M.; Quaglia, F.; Oldfield, C.J.; Aspromonte, M.C.; Davey, N.E.; Davidovic, R.; Dosztanyi, Z.; et al. DisProt 7.0: A major update of the database of disordered proteins. Nucleic Acids Res. 2017, 45, D219–D227. [Google Scholar] [CrossRef] [Green Version]
- Resh, M.D. Fatty acylation of proteins: New insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim. Biophys. Acta Mol. Cell Res. 1999, 1451, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Castrec, B.; Dian, C.; Ciccone, S.; Ebert, C.L.; Bienvenut, W.V.; Le Caer, J.-P.; Steyaert, J.-M.; Giglione, C.; Meinnel, T. Structural and genomic decoding of human and plant myristoylomes reveals a definitive recognition pattern. Nat. Chem. Biol. 2018, 14, 671–679. [Google Scholar] [CrossRef]
- Charron, G.; Li, M.M.H.; MacDonald, M.R.; Hang, H.C. Prenylome profiling reveals S-farnesylation is crucial for membrane targeting and antiviral activity of ZAP long-isoform. Proc. Natl. Acad. Sci. USA 2013, 110, 11085–11090. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, S.; Murray, D. Plasma membrane phosphoinositide organization by protein electrostatics. Nature 2005, 438, 605–611. [Google Scholar] [CrossRef]
- Michailidis, I.E.; Rusinova, R.; Georgakopoulos, A.; Chen, Y.; Iyengar, R.; Robakis, N.K.; Logothetis, D.E.; Baki, L. Phosphatidylinositol-4,5-bisphosphate regulates epidermal growth factor receptor activation. Pflügers Arch. Eur. J. Physiol. 2011, 461, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkhipov, A.; Shan, Y.; Das, R.; Endres, N.F.; Eastwood, M.P.; Wemmer, D.E.; Kuriyan, J.; Shaw, D.E. Architecture and membrane interactions of the EGF receptor. Cell 2013, 152, 557–569. [Google Scholar] [CrossRef] [Green Version]
- Halim, K.B.A.; Koldsø, H.; Sansom, M.S. Interactions of the EGFR juxtamembrane domain with PIP2-containing lipid bilayers: Insights from multiscale molecular dynamics simulations. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1017–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedger, G.; Sansom, M.S.P.; Koldsø, H. The juxtamembrane regions of human receptor tyrosine kinases exhibit conserved interaction sites with anionic lipids. Sci. Rep. 2015, 5, 9198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Bogaart, G.; Meyenberg, K.; Risselada, H.J.; Amin, H.; Willig, K.I.; Hubrich, B.E.; Dier, M.; Hell, S.W.; Grubmuller, H.; Diederichsen, U.; et al. Membrane protein sequestering by ionic protein-lipid interactions. Nature 2011, 479, 552–555. [Google Scholar] [CrossRef]
- Hädicke, A.; Blume, A. Binding of cationic model peptides (KX) 4 K to anionic lipid bilayers: Lipid headgroup size influences secondary structure of bound peptides. Biochim. Biophys. Acta Biomembr. 2017, 1859, 415–424. [Google Scholar] [CrossRef]
- Sinthuvanich, C.; Veiga, A.S.; Gupta, K.; Gaspar, D.; Blumenthal, R.; Schneider, J.P. Anticancer β-hairpin peptides: Membrane-induced folding triggers activityc. J. Am. Chem. Soc. 2012, 134, 6210–6217. [Google Scholar] [CrossRef] [Green Version]
- Almo, S.C.; Bonanno, J.B.; Sauder, J.M.; Emtage, S.; DiLorenzo, T.P.; Malashkevich, V.; Wasserman, S.R.; Swaminathan, S.; Eswaramoorthy, S.; Agarwal, R.; et al. Structural genomics of protein phosphatases. J. Struct. Funct. Genom. 2007, 8, 121–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Roux, A.L.; Busquets, M.A.; Sagués, F.; Pons, M. Kinetics characterization of c-Src binding to lipid membranes: Switching from labile to persistent binding. Colloids Surf. B Biointerfaces 2016, 138, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, A.L.; Castro, B.; Garbacik, E.T.; Garcia Parajo, M.F.; Pons, M. Single molecule fluorescence reveals dimerization of myristoylated Src N-terminal region on supported lipid bilayers. ChemistrySelect 2016, 1, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Couet, J.; Lisanti, M.P. Src tyrosine kinases, G(α) subunits, and H-Ras share a common membrane-anchored scaffolding protein, caveolin: Caveolin binding negatively regulates the auto-activation of Src tyrosine kinases. J. Biol. Chem. 1996, 271, 29182–29190. [Google Scholar] [CrossRef] [Green Version]
- Barker, S.; Kassel, D.B.; Weigl, D.; Huang, X.; Luther, M.; Knight, W.B. Characterization of pp60c-src Tyrosine Kinase Activities Using a Continuous Assay: Autoactivation of the Enzyme Is an Intermolecular Autophosphorylation Process. Biochemistry 1995, 34, 14843–14851. [Google Scholar] [CrossRef]
- Le Roux, A.L.; Mohammad, I.L.; Mateos, B.; Arbesú, M.; Gairí, M.; Khan, F.A.; Farman, A.; Teixeira, J.; Pons, M. A Myristoyl-Binding Site in the SH3 Domain Modulates c-Src Membrane Anchoring. iScience 2019, 12, 194–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, G.; Delbrück, M. Reduction of Dimensionality in Biological Diffusion Processes. In Structural Chemistry and Molecular Biology; Rich, A., Davidson, N., Eds.; W.H. Freeman and Co.: San Francisco, CA, USA, 1968. [Google Scholar]
- Berg, O.G.; Winter, R.B.; Von Hippel, P.H. Diffusion-Driven Mechanisms of Protein Translocation on Nucleic Acids. 1. Models and Theory. Biochemistry 1981, 20, 6929–6948. [Google Scholar] [CrossRef] [PubMed]
- Cherry, R.J. Rotational and lateral diffusion of membrane proteins. Biochim. Biophys. Acta Rev. Biomembr. 1979, 559, 289–327. [Google Scholar] [CrossRef]
- Kholodenko, B.N.; Brown, G.C.; Hoek, J.B. Diffusion control of protein phosphorylation in signal transduction pathways. Biochem. J. 2000, 350, 901–907. [Google Scholar] [CrossRef]
- Kholodenko, B.N.; Hoek, J.; Westerhoff, H. Why cytoplasmic signalling proteins should be recruited to cell membranes. Trends Cell Biol. 2000, 10, 173–178. [Google Scholar] [CrossRef]
- Saffman, P.G.; Delbruck, M. Brownian motion in biological membranes. Proc. Natl. Acad. Sci. USA 1975, 72, 3111–3113. [Google Scholar] [CrossRef] [Green Version]
- Hughes, B.D.; Pailthorpe, B.A.; White, L.R. The Translational and Rotational Drag on a Cylinder Moving in a Membrane. J. Fluid Mech. 1981, 110, 349–372. [Google Scholar] [CrossRef]
- Petrov, E.P.; Schwille, P. Translational diffusion in lipid membranes beyond the Saffman-Delbrück approximation. Biophys. J. 2008, 94, L41–L43. [Google Scholar] [CrossRef] [Green Version]
- Goutaland, Q.; Fournier, J.-B. Saffman-Delbrück and beyond: A pointlike approach. Eur. Phys. J. E 2019, 42, 156. [Google Scholar] [CrossRef] [Green Version]
- Javanainen, M.; Martinez-Seara, H.; Metzler, R.; Vattulainen, I. Diffusion of Integral Membrane Proteins in Protein-Rich Membranes. J. Phys. Chem. Lett. 2017, 8, 4308–4313. [Google Scholar] [CrossRef]
- Ziemba, B.P.; Falke, J.J. Lateral diffusion of peripheral membrane proteins on supported lipid bilayers is controlled by the additive frictional drags of (1) bound lipids and (2) protein domains penetrating into the bilayer hydrocarbon core. Chem. Phys. Lipids 2013, 172-173, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, G.; Im, W.; Sugita, Y.; Feig, M. Clustering and dynamics of crowded proteins near membranes and their influence on membrane bending. Proc. Natl. Acad. Sci. USA 2019, 116, 24562–24567. [Google Scholar] [CrossRef]
- Schleicher, K.D.; Dettmer, S.L.; Kapinos, L.E.; Pagliara, S.; Keyser, U.; Jeney, S.; Lim, R. Selective transport control on molecular velcro made from intrinsically disordered proteins. Nat. Nanotechnol. 2014, 9, 525–530. [Google Scholar] [CrossRef]
- Sil, P.; Mateos, N.; Nath, S.; Buschow, S.; Manzo, C.; Suzuki, K.G.N.; Fujiwara, T.; Kusumi, A.; Garcia-Parajo, M.F.; Mayor, S. Dynamic actin-mediated nano-scale clustering of CD44 regulates its meso-scale organization at the plasma membrane. Mol. Biol. Cell 2020, 31, 561–579. [Google Scholar] [CrossRef]
- Loverdo, C.; Bénichou, O.; Moreau, M.; Voituriez, R. Enhanced reaction kinetics in biological cells. Nat. Phys. 2008, 4, 134–137. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pons, M. Basic Residue Clusters in Intrinsically Disordered Regions of Peripheral Membrane Proteins: Modulating 2D Diffusion on Cell Membranes. Physchem 2021, 1, 152-162. https://doi.org/10.3390/physchem1020010
Pons M. Basic Residue Clusters in Intrinsically Disordered Regions of Peripheral Membrane Proteins: Modulating 2D Diffusion on Cell Membranes. Physchem. 2021; 1(2):152-162. https://doi.org/10.3390/physchem1020010
Chicago/Turabian StylePons, Miquel. 2021. "Basic Residue Clusters in Intrinsically Disordered Regions of Peripheral Membrane Proteins: Modulating 2D Diffusion on Cell Membranes" Physchem 1, no. 2: 152-162. https://doi.org/10.3390/physchem1020010