
Native Carnivore Diversity and Relative Abundance in Landscapes of the Coast Range in Central Chile: Insights for Conservation Decision-Making
 , , ,
, , ,
Abstract
:1. Introduction
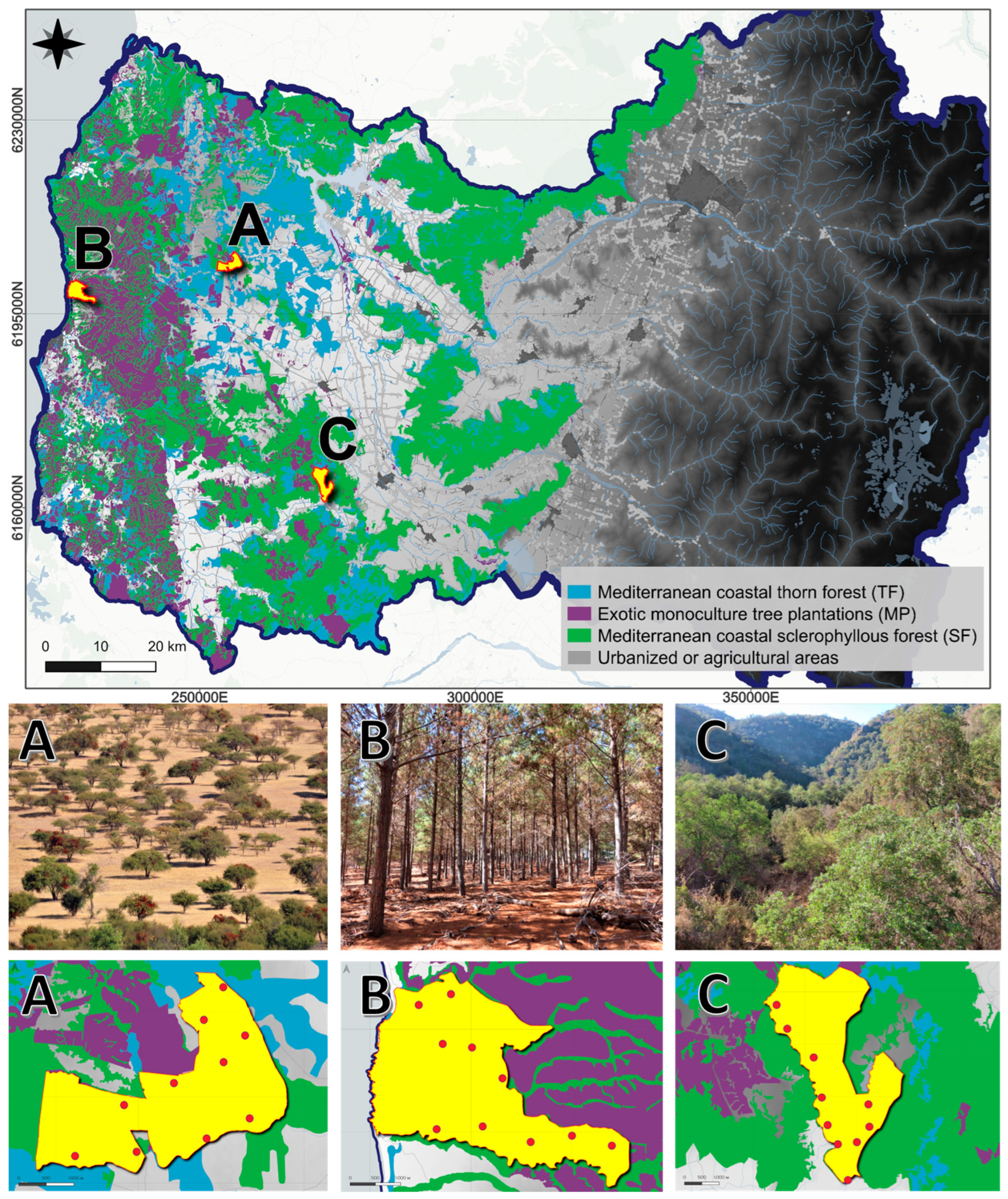
2. Methods
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferreira, A.S.; Peres, C.A.; Dodonov, P.; Cassano, C.C. Multi-scale mammal responses to agroforestry landscapes in the Brazilian Atlantic Forest: The conservation value of forest and traditional shade plantations. Agrofor. Syst. 2020, 94, 2331–2341. [Google Scholar] [CrossRef]
- de Abreu Pestana, L.F.; Martello, F.; Fonseca, R.C.B. Richness and composition of terrestrial mammals vary in eucalyptus plantations due to stand age. Austral Ecol. 2023, 48, 743–760. [Google Scholar] [CrossRef]
- Cravino, A.; Martínez-Lanfranco, J.A.; Brazeiro, A. Community structure of medium-large mammals across a tree plantation cycle in natural grasslands of Uruguay. For. Ecol. Manag. 2023, 529, 120713. [Google Scholar] [CrossRef]
- Miranda, A.; Altamirano, A.; Cayuela, L.; Lara, A.; González, M. Native forest loss in the Chilean biodiversity hotspot: Revealing the evidence. Reg. Environ. Chang. 2017, 17, 285–297. [Google Scholar] [CrossRef]
- Zamorano-Elgueta, C.; Benayas, J.M.R.; Cayuela, L.; Hantson, S.; Armenteras, D. Native forest replacement by exotic plantations in southern Chile (1985–2011) and partial compensation by natural regeneration. For. Ecol. Manag. 2015, 345, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Cunill, P. Geografía de Chile, 9th ed.; Editorial Universitaria: Santiago, Chile, 1979; p. 557. [Google Scholar]
- Suarez, M.; Naranjo, J.; Puig, A. Estratigrafía de la Cordillera de la Costa, al sur de Taltal, Chile: Etapas iniciales de la evolución Andina. Rev. Geológica Chile 2010, 24, 19–28. [Google Scholar] [CrossRef]
- Armesto, J.J.; Aravena, J.C.; Villagran, C.; Perez, C.; Parker, G. Bosques Templados de la Cordillera de la Costa; Editorial Universitaria: Santiago, Chile, 1996. [Google Scholar]
- Smith-Ramírez, C. The Chilean coastal range: A vanishing center of biodiversity and endemism in South American temperate rainforests. Biodivers. Conserv. 2004, 13, 373–393. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Weinzettel, J.; Vačkář, D.; Medková, H. Human footprint in biodiversity hotspots. Front. Ecol. Environ. 2018, 16, 447–452. [Google Scholar] [CrossRef]
- Miller, B.; Dugelby, B.; Foreman, D.; Del Rio, C.M.; Noss, R.; Phillips, M.; Soulé, M.; Terborgh, J.; Willcox, L. The importance of large carnivores to healthy ecosystems. Endanger. Species Update 2001, 18, 202–210. [Google Scholar]
- Prevosti, F.J.; Pereira, J.A. Community Structure of South American Carnivores in the Past and Present. J. Mamm. Evol. 2014, 21, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Noss, R.F.; Quigley, H.B.; Hornocker, M.G.; Merrill, T.; Paquet, P.C. Conservation biology and carnivore conservation in the Rocky Mountains. Conserv. Biol. 1996, 10, 949–963. [Google Scholar] [CrossRef]
- Thorne, J.H.; Cameron, D.; Quinn, J.F. A conservation design for the central coast of California and the evaluation of mountain lion as an umbrella species. Nat. Areas J. 2006, 26, 137–148. [Google Scholar] [CrossRef]
- Garcia, C.B.; Svensson, G.L.; Bravo, C.; Undurraga, M.I.; Diaz-Forestier, J.; Godoy, K.; Neaman, A.; Barbosa, O.; Abades, S.; Celis-Diez, J.L. Remnants of native forests support carnivore diversity in the vineyard landscapes of central Chile. Oryx 2021, 55, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Glen, A.; Dickman, C. Complex interactions among mammalian carnivores in Australia, and their implications for wildlife management. Biol. Rev. 2005, 80, 387–401. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.S.; Hebblewhite, M.; Boitani, L.; Powell, R.A. Carnivore habitat ecology: Integrating theory and application. In Carnivore Ecology and Conservation: A Handbook of Techniques; Boitani, L., Powell, R.A., Eds.; Oxford University Press: London, UK, 2012; pp. 218–255. ISBN 978-019-177-454-6. [Google Scholar]
- Long, R.A.; Donovan, T.M.; MacKay, P.; Zielinski, W.J.; Buzas, J.S. Predicting carnivore occurrence with noninvasive surveys and occupancy modeling. Landsc. Ecol. 2011, 26, 327–340. [Google Scholar] [CrossRef]
- Moreira-Arce, D.; Silva-Rodríguez, E.A.; Napolitano, C.; D’Elía, G.; Cabello, J.; Millán, J.; Hidalgo-Hermoso, E.; Farías, A.A. A forest-specialist carnivore in the middle of the desert? Comments on Anabalon et al. 2019. Ecol. Evol. 2020, 10, 3825–3830. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.S.; Bailey, L.L.; Van de Woude, S.; Crooks, K.R. Interspecific interactions between wild felids vary across scales and levels of urbanization. Ecol. Evol. 2015, 5, 5946–5961. [Google Scholar] [CrossRef]
- Chanchani, P.; Noon, B.R.; Bailey, L.; Warrier, R.A. Conserving tigers in working landscapes. Conserv. Biol. 2016, 30, 649–660. [Google Scholar] [CrossRef]
- Grassel, S.M.; Rachlow, J.L.; Williams, C.J. Spatial interactions between sympatric carnivores: Asymmetric avoidance of an intraguild predator. Ecol. Evol. 2015, 5, 2762–2773. [Google Scholar] [CrossRef] [Green Version]
- Moreira-Arce, D.; Cabello, J.; Meneses, L.O.; Norambuena, K.; Perez-Hernandez, C.G.; Hidalgo-Hermoso, E.; Alaniz, A.J.; Vergara, P.M. Scale-dependent habitat use from an individual-based perspective: The case of the endangered Darwin’s fox living in heterogeneous forest landscapes. Landsc. Ecol. 2021, 36, 513–526. [Google Scholar] [CrossRef]
- Linnell, J.D.C.; Strand, O. Interference interactions, co-existence and conservation of mammalian carnivores. Divers. Distrib. 2000, 6, 169–176. [Google Scholar] [CrossRef]
- Gil-Sánchez, J.M.; Mañá-Varela, B.; Herrera-Sánchez, F.J.; Urios, V. Spatio-temporal ecology of a carnivore community in middle atlas, NW of Morocco. Zoology 2021, 146, 125904. [Google Scholar] [CrossRef] [PubMed]
- Arias-Alzate, A.; Arroyave, F.; Romero, O.; Hurtado, R.; Gonzalez-Maya, J.F.; Arroyo-Cabrales, J.; Townsend, A.; Martinez-Meyer, A. Functional niche constraints on carnivore assemblages (Mammalia: Carnivora) in the Americas: What facilitates coexistence through space and time? J. Biogeogr. 2022, 49, 445–575. [Google Scholar] [CrossRef]
- Andrade-Núñez, M.J.; Aide, T.M. Effects of Habitat and Landscape Characteristics on Medium and Large Mammal Species Richness and Composition in Northern Uruguay. Zoologia 2010, 27, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Lyra-Jorge, M.C.; Ribeiro, M.C.; Ciocheti, G.; Tambosi, L.R.; Pivello, V.R. Influence of multi-scale landscape structure on the occurrence of carnivorous mammals in a human-modified savanna, Brazil. Eur. J. Wildl. Res. 2010, 56, 359–368. [Google Scholar] [CrossRef]
- Lantschner, M.V.; Rusch, V.; Hayes, J.P. Habitat use by carnivores at different spatial scales in a plantation forest landscape in Patagonia, Argentina. For. Ecol. Manag. 2012, 269, 271–278. [Google Scholar] [CrossRef]
- Ramírez-Alvarez, D. Fauna Nativa de la Región de O’Higgins: Vertebrados Terrestres; Grafica Andes: O’Higgins Region, Chile, 2018; p. 504. ISBN 978-956-398-064-6. [Google Scholar]
- Jaksic, F.M. Ecología de los Vertebrados de Chile; Ediciones Universidad Católica de Chile: Santiago, Chile, 1997; p. 262. ISBN 956-140-440-0. [Google Scholar]
- Salvatori, V.; Vaglio-Laurin, G.; Meserve, P.L.; Boitani, L.; Campanella, A. Spatial organization, activity, and social interactions of culpeo foxes (Pseudalopex culpaeus) in north central Chile. J. Mammal. 1999, 80, 980–985. [Google Scholar] [CrossRef] [Green Version]
- Zuñiga, A.; Muñoz-Pedrero, A.; Fierro, A. Dieta de Lycalopex griseus (Gray, 1837) (Mammalia: Canidae) en la depresión intermedia del Sur de Chile. Gayana 2008, 72, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Silva-Rodríguez, E.A.; Ortega-Solís, G.R.; Jiménez, J.E. Conservation and ecological implications of the use of space by chilla foxes and free-ranging dogs in a human-dominated landscape in southern Chile. Austral Ecol. 2010, 35, 765–777. [Google Scholar] [CrossRef]
- Silva, F. Filogeografia e História Populacional de Lycalopex vetulus (Carnivora, Canidae), Incluindo Sua Hibridação Com L. gymnocercus. Master’s Dissertation, Faculdade de Biociências, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brasil, 2015. Available online: https://tede2.pucrs.br/tede2/handle/tede/6409#preview-link0 (accessed on 23 May 2022).
- Tchaicka, L.; Freitas, T.R.; Bager, A.; Vidal, S.L.; Lucherini, M.; Iriarte, A.; Novaro, A.; Geffen, E.; Garcez, F.S.; Johnson, W.E.; et al. Molecular assessment of the phylogeny and biogeography of a recently diversified endemic group of South American canids (Mammalia: Carnivora: Canidae). Genet. Mol. Biol. 2016, 39, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Chemisquy, M.A.; Prevosti, F.J.; Martinez, P.; Raimondi, V.; Stom, J.E.C.; Acosta-Jamett, G.; Montoya-Burgos, J.I. How many species of grey foxes (Canidae, Carnivora) are there in southern South America? Mastozoología Neotrop. 2019, 26, 81–97. [Google Scholar] [CrossRef]
- Ramírez-Álvarez, D.; Napolitano, C.; Salgado, I. Puma (Puma concolor) in the neighborhood? Records near human settlements and insights into human-carnivore coexistence in central Chile. Animals 2021, 11, 965. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, C.; Pino, R.; Donoso, D.; Iriarte, A. El Puma en la Región de Coquimbo: Develando Su Ecología e Interacción con la Ganadería, 2nd ed.; Ediciones SAG Región de Coquimbo y Flora & Fauna Chile: Santiago, Chile, 2017; p. 62. [Google Scholar]
- Napolitano, C.; Johnson, W.E.; Sanderson, J.; O’Brien, S.J.; Hoelzel, A.R.; Freer, R.; Dunstone, N.; Ritland, K.; Ritland, C.E.; Poulin, E. Phylogeography and population history of Leopardus guigna, the smallest American felid. Conserv. Genet. 2014, 15, 631–653. [Google Scholar] [CrossRef]
- Napolitano, C.; Gálvez, N.; Bennett, M.; Acosta-Jamett, G.; Sanderson, J. Leopardus guigna, Guiña. The IUCN Red List of Threatened Species 2015b: E.T15311A50657245. Available online: https://doi.org/10.2305/IUCN.UK.2015-2.RLTS.T15311A50657245.en (accessed on 9 June 2023).
- Napolitano, C.; Larraguibel-González, C.; Cepeda-Mercado, A.; Vial, P.; Sanderson, J. New records of Leopardus guigna in its northern-most distribution in Chile: Implications for conservation. Rev. Chil. Hist. Nat. 2020, 93, 7. [Google Scholar] [CrossRef]
- Dunstone, N.; Durbin, L.; Wyllie, I.; Freer, R.; Acosta, G.; Mazzolli, M.; Rose, S. Spatial organization, ranging behavior and habitat use of the kodkod (Oncifelis guigna) in southern Chile. J. Zool. 2002, 257, 1–11. [Google Scholar] [CrossRef]
- Sanderson, J.; Sunquist, M.E.; Iriarte, A.W. Natural history and landscape-use of guignas (Oncifelis guigna) on Isla Grande de Chiloé, Chile. J. Mammal. 2002, 83, 608–613. [Google Scholar] [CrossRef]
- Schüttler, E.; Klenke, R.; Galuppo, S.; Castro, R.A.; Bonacic, C.; Laker, J.; Henle, K. Habitat use and sensitivity to fragmentation in America’s smallest wildcat. Mamm. Biol. 2017, 86, 1–8. [Google Scholar] [CrossRef]
- Gálvez, N.; Hernández, F.; Laker, J.; Gilabert, H.; Petitpas, R.; Bonacic, C.; Gimona, A.; Hester, A.; MacDonald, D. Forest cover outside protected areas plays an important role in the conservation of the vulnerable guiña Leopardus guigna. Oryx 2013, 47, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Galvez, N.; Infante, J.; Fernandez, A.; Diaz, J.; Petracca, L. Land use intensification coupled with free-roaming dogs as potential defaunation drivers of mesocarnivores in agricultural landscapes. J. Appl. Ecol. 2021, 58, 2962–2974. [Google Scholar] [CrossRef]
- Napolitano, C.; Díaz, D.; Sanderson, J.; Johnson, W.E.; Ritland, K.; Ritland, C.E.; Poulin, E. Reduced genetic diversity and increased dispersal in Guigna (Leopardus guigna) in Chilean fragmented landscapes. J. Hered. 2015, 106 (Suppl. 1), 522–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-Jamett, G.; Simonetti, J.A. Habitat use by Oncifelis guigna and Pseudalopex culpaeus in a fragmented forest landscape in Central Chile. Biodivers. Conserv. 2004, 13, 1135–1151. [Google Scholar] [CrossRef]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group. Cat News Spec. Issue 2017, 11, 80. [Google Scholar]
- Nascimento, F.O.D.; Cheng, J.; Feijó, A. Taxonomic revision of the pampas cat Leopardus colocola complex (Carnivora: Felidae): An integrative approach. Zool. J. Linn. Soc. 2021, 191, 575–611. [Google Scholar] [CrossRef]
- Castro-Pastene, C.; González, C.; Carrasco, H.; Soto-Gamboa, M. 2021. Registros de diferentes hábitats usados por el Colo-Colo (Leopardus colocola colocola) Molina 1782, en la zona central de Chile. Gayana 2001, 85, 84–89. [Google Scholar] [CrossRef]
- Tellaeche, C. Ecología y Uso del Espacio de dos Especies de Félidos, Gato Andino (Leopardus jacobita) y Gato del Pajonal (L. colocolo) En La Región Altoandina, Provincia de Jujuy. Ph.D. Thesis, Universidad Nacional Del Sur, Bahía Blanca, Argentina, 2015. [Google Scholar]
- Castillo, D.F.; Vidal, E.M.L.; Caruso, N.C.; Lucherini, M.; Casanave, E.B. Denning ecology of Conepatus chinga (Carnivora: Mephitidae) in a grassland relict of central Argentina. Mastozoología Neotrop. 2013, 20, 373–379. [Google Scholar]
- Donadio, E.; Di Martino, S.; Aubone, M.; Novaro, A.J. Activity patterns, home-range, and habitat selection of the common hog-nosed skunk, Conepatus chinga (Mammmalia, Mustelidae), in northwestern Patagonia. Mammalia 2001, 65, 49–54. [Google Scholar] [CrossRef]
- Tellaeche, C.G.; Reppucci, J.I.; Vidal, E.M.L.; Lucherini, M. New data on the distribution and natural history of the lesser grison (Galictis cuja), hog-nosed skunk (Conepatus chinga), and culpeo (Lycalopex culpaeus) in northwestern Argentina. Mammalia 2014, 78, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile, 2nd ed.; Editorial Universitaria: Santiaog, Chile, 2018; p. 384. ISBN 978-956-112-575-9. [Google Scholar]
- MINAGRI. Catastro y Actualización de los Recursos Vegetacionales y Uso de la Tierra de la Región de O’Higgins (VI) CONAF. IDE Infraestructura de Datos Espaciales del Ministerio de Agricultura de Chile. 2013. Available online: https://ide.minagri.gob.cl/geoweb/2019/11/22/planificacion-catastral/ (accessed on 14 June 2022).
- Heilmayr, R.; Echeverría, C.; Fuentes, R.; Lambin, E.F. A plantation-dominated forest transition in Chile. Appl. Geogr. 2016, 75, 71–82. [Google Scholar] [CrossRef]
- Astorga-Schneider, L.; Burschel, H. Chile Necesita un Nuevo Modelo Forestal Ante Los Desafíos Climáticos, Sociales y Ambientales; LOM Ediciones: Santiago, Chile, 2019; ISBN 978-956-00-1239-5. [Google Scholar]
- Becerra, P.; Simonetti-Zambelli, J.; Native and Exotic Plant Species Diversity in Forest Fragments and Forestry Plantations of a Coastal Landscape of Central Chile. Repositorio Académico Universidad de Chile. Available online: https://repositorio.uchile.cl/handle/2250/177895 (accessed on 21 October 2022).
- Trolliet, F.; Vermeulen, C.; Huynen, M.C.; Hambuckers, A. Use of camera traps for wildlife studies: A review. Biotechnol. Agron. Société Environ. 2014, 18, 446–454. [Google Scholar]
- Sanderson, J.; Harris, G. Automatic data organization, storage, and analysis of camera trap pictures. J. Indones. Nat. Hist. 2013, 1, 11–19. [Google Scholar]
- SWCCF. Small Wild Cat Conservation Foundation Webpage. Available online: https://smallcats.org/resources/#camerasweet (accessed on 15 June 2022).
- Vásquez-Ibarra, V.; Cortés, E.I.; Silva-Rodríguez, E.A. Tutorial para la Clasificación de Imágenes Obtenidas Mediante Cámaras Trampa; Universidad Austral de Chile: Valdivia, Chile, 2021. [Google Scholar]
- Yoshizaki, J.; Pollock, K.H.; Brownie, C.; Webster, R.A. Modeling misidentification errors in capture-recapture studies using photographic identification of evolving marks. Ecology 2009, 90, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.J.; Harmsen, B.J. A critique of density estimation from camera-trap data. J. Wildl. Manag. 2012, 76, 224–236. [Google Scholar] [CrossRef]
- Johansson, Ö.; Samelius, G.; Wikberg, E.; Chapron, G.; Mishra, C.; Low, M. Identification errors in camera-trap studies result in systematic population overestimation. Sci. Rep. 2020, 10, 6393. [Google Scholar] [CrossRef] [Green Version]
- Dohoo, I.; Martin, W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; AVC Inc.: Charlottetown, PE, Canada, 2009; p. 865. ISBN 978-091-901-360-5. [Google Scholar]
- Zuñiga, A.; Muñoz-Pedreros, A.; Fierro, A. Habitat use of four terrestrial carnivores in southern Chile. Gayana 2009, 73, 200–210. [Google Scholar]
- Zúñiga, A.; Fuenzalida, V.; Sandoval, R. Diet of the gray fox Lycalopex griseus in an agroecosystem of southern-central Chile. Therya 2018, 9, 179–183. [Google Scholar] [CrossRef]
- Lagos, N.; Villalobos, R.; Vianna, J.A.; Espinosa-Miranda, C.; Rau, J.R.; Iriarte, A. The spatial and trophic ecology of culpeo foxes (Lycalopex culpaeus) in the high Andes of northern Chile. Stud. Neotrop. Fauna Environ. 2021, 1–10. [Google Scholar] [CrossRef]
- SAG. Actas de Movimiento Animal para Rescate y Rehabilitación de Fauna Silvestre. Periodo 2018–2022; Servicio Agrícola y Ganadero de la Región de O’Higgins: Región de O’Higgins, Chile, 2022. [Google Scholar]
- Yensen, E.; Tarifa, T. Galictis cuja. Mamm. Species 2003, 728, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Beltrami, E.; Gálvez, N.; Osorio, C.; Kelly, M.J.; Morales-Moraga, D.; Bonacic, C. Ravines as conservation strongholds for small wildcats under pressure from free-ranging dogs and cats in Mediterranean landscapes of Chile. Stud. Neotrop. Fauna Environ. 2021, 58, 138–154. [Google Scholar] [CrossRef]
- Palomares, F.; Delibes, M.; Ferreras, P.; Fedriani, J.M.; Calzada, J.; Revilla, E. Iberian Lynx in a fragmented landscape: Predispersal, dispersal and postdispersal habitats. Conserv. Biol. 2000, 14, 809–818. [Google Scholar] [CrossRef]
- Schneider, A.; Henegar, C.; Day, K.; Absher, D.; Napolitano, C.; Silveira, L.; David, V.A.; O’Brien, S.J.; Menotti-Raymond, M.; Barsh, G.S.; et al. Recurrent Evolution of Melanism in South American Felids. PLoS Genet. 2015, 11, e1004892. [Google Scholar] [CrossRef]
- MMA Ministerio del Medio Ambiente de Chile. Inventario Nacional de Especies de Chile: Leopardus colocolo (Molina, 1782). Available online: http://especies.mma.gob.cl/CNMWeb/Web/WebCiudadana/ficha_indepen.aspx?EspecieId=11&Version=1 (accessed on 17 August 2022).
- Tirelli, F.P.; Trigo, T.C.; Queirolo, D.; Kasper, C.B.; Bou, N.; Peters, F.; Mazim, F.D.; Martínez-Lanfranco, J.A.; González, E.M.; Espinosa, C. High extinction risk and limited habitat connectivity of Muñoa’s pampas cat, an endemic felid of the Uruguayan Savanna ecoregion. J. Nat. Conserv. 2021, 62, 126009. [Google Scholar] [CrossRef]
- Guzmán-Marín, B.C.; Hernández-Hernández, J.C.; de Aguilera, N.S.O.; Muñoz-Peña, P.A.; Kaiser, M. Felino pequeño, ciudad grande: Leopardus colocola (Carnivora: Felidae) en la Region Metropolitana de Santiago de Chile y amenazas para su conservacion. Bol. Mus. Nac. Hist. Nat. 2022, 71, 13–21. [Google Scholar] [CrossRef]
- Pereira, P.; da Silva, A.A.; Alves, J.; Matos, M.; Fonseca, C. Coexistence of carnivores in a heterogeneous landscape: Habitat selection and ecological niches. Ecol. Res. 2012, 27, 745–753. [Google Scholar] [CrossRef]
- Benson, J.; Chamberlain, M. Space use and habitat selection by female Louisiana black bears in the Tensas river basin of Louisiana. J. Wildl. Manag. 2007, 71, 117–126. [Google Scholar] [CrossRef]
- Fernández, N.; Delibes, M.; Palomares, F. Habitat-related heterogeneity in breeding in a metapopulation of the Iberian lynx. Ecography 2007, 30, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Pulliam, H.R. Sources, sinks, and population regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Robinson, H.S.; Wielgus, R.B.; Cooley, H.S.; Cooley, S.W. Sink populations in carnivore management: Cougar demography and immigration in a hunted population. Ecol. Appl. 2008, 18, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Coon, C.A.C.; Mahoney, P.J.; Edelblutte, E.; McDonald, Z.; Stoner, D.C. Predictors of puma occupancy indicate prey vulnerability is more important than prey availability in a highly fragmented landscape. Wildl. Biol. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hopcraft, J.G.C.; Sinclair, A.R.E.; Packer, C. Planning for success: Serengeti lions seek prey accessibility rather than abundance. J. Anim. Ecol. 2005, 74, 559–566. [Google Scholar] [CrossRef]
- Villaseñor, N.R.; Escobar, M.A.; Estades, C.F. There is no place like home: High homing rate and increased mortality after translocation of a small mammal. Eur. J. Wildl. Res. 2013, 59, 749–760. [Google Scholar] [CrossRef]
- Pliscoff, P.; Simonetti, J.A.; Grez, A.A.; Vergara, P.M.; Barahona-Segovia, R.M. Defining corridors for movement of multiple species in a forest-plantation landscape. Glob. Ecol. Conserv. 2020, 23, e01108. [Google Scholar] [CrossRef]
- Aravena, J.C. Análisis de la Estructura y la Flora de los Bosques de Chiloe Continental e Insular. Master’s Thesis, Facultad de Ciencias, Universidad de Chile, Santiago, Chile, 1991. [Google Scholar]
- Murúa, R. Comunidades de Mamiferos del Bosque Templado de Chile. In Ecologia de los Bosques Nativos de Chile; Armesto, J.J., Villagran, C., Kalin, M., Eds.; Editorial Universitaria: Santiago, Chile, 1996; ISBN 978-956-111-180-6. [Google Scholar]
- CONAF. Listado Sistema Nacional de Áreas Silvestres Protegidas del Estado (SNASPE). Diciembre 2020. Corporación Nacional Forestal de Chile. Available online: https://www.conaf.cl/wp-content/files_mf/1610052227ListadoSNASPEActualizado21122020.pdf (accessed on 23 March 2022).
- Acosta-Jamett, G.; Simonetti, J.A.; Bustamante, R.O.; Dunstone, N. Metapopulation approach to assess survival of Oncifelis guigna in fragmented forests of central Chile: A theoretical model. Mastozoología Neotrop. 2003, 10, 217–229. [Google Scholar]
- Sweanor, L.L.; Logan, K.A.; Hornocker, M.G. Cougar dispersal patterns, metapopulation dynamics, and conservation. Conserv. Biol. 2000, 14, 798–808. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
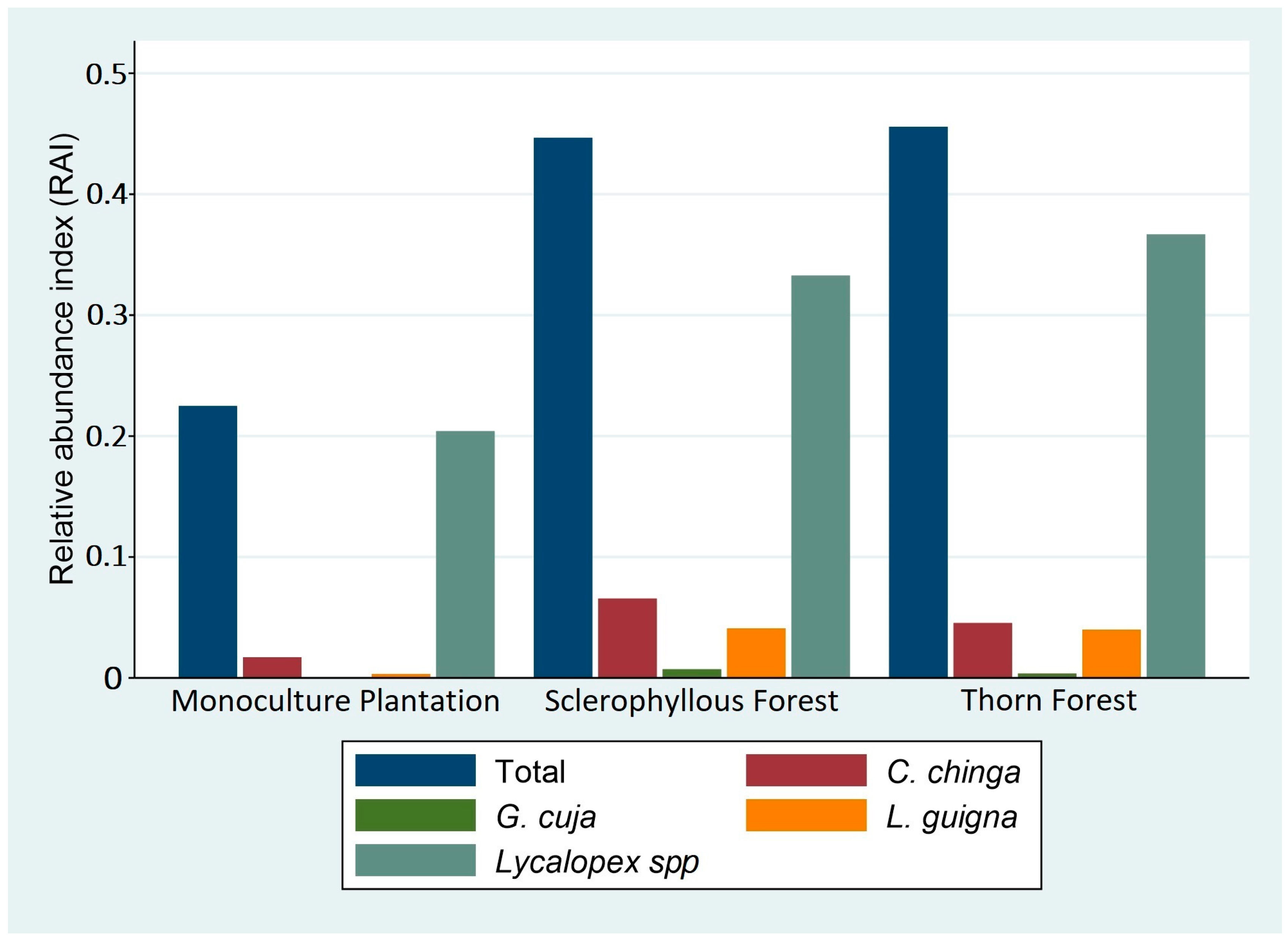
| Location | Landscape | Total Sampling Effort (Days × Camera) | N° of Cameras | Total Number of Independent Events (RAI = n/days) | ||||
|---|---|---|---|---|---|---|---|---|
| C. chinga | G. cuja | L. guigna | Lycalopex spp. | All Species | ||||
| Alto Colorado | Monoculture tree plantation (MP) | 3446 | 10 | 59 (0.0171) | 2 (0.006) | 11 (0.0032) | 703 (0.2040) | 775 (0.2249) |
| Callihue | Sclerophyllous forest (SF) | 3502 | 10 | 230 (0.0657) | 25 (0.0071) | 144 (0.0411) | 1165 (0.3327) | 1564 (0.4466) |
| La Estrella | Thorn forest (TF) | 3098 | 10 | 141 (0.0455) | 11 (0.0036) | 124 (0.0400) | 1136 (0.3667) | 1412 (0.4558) |
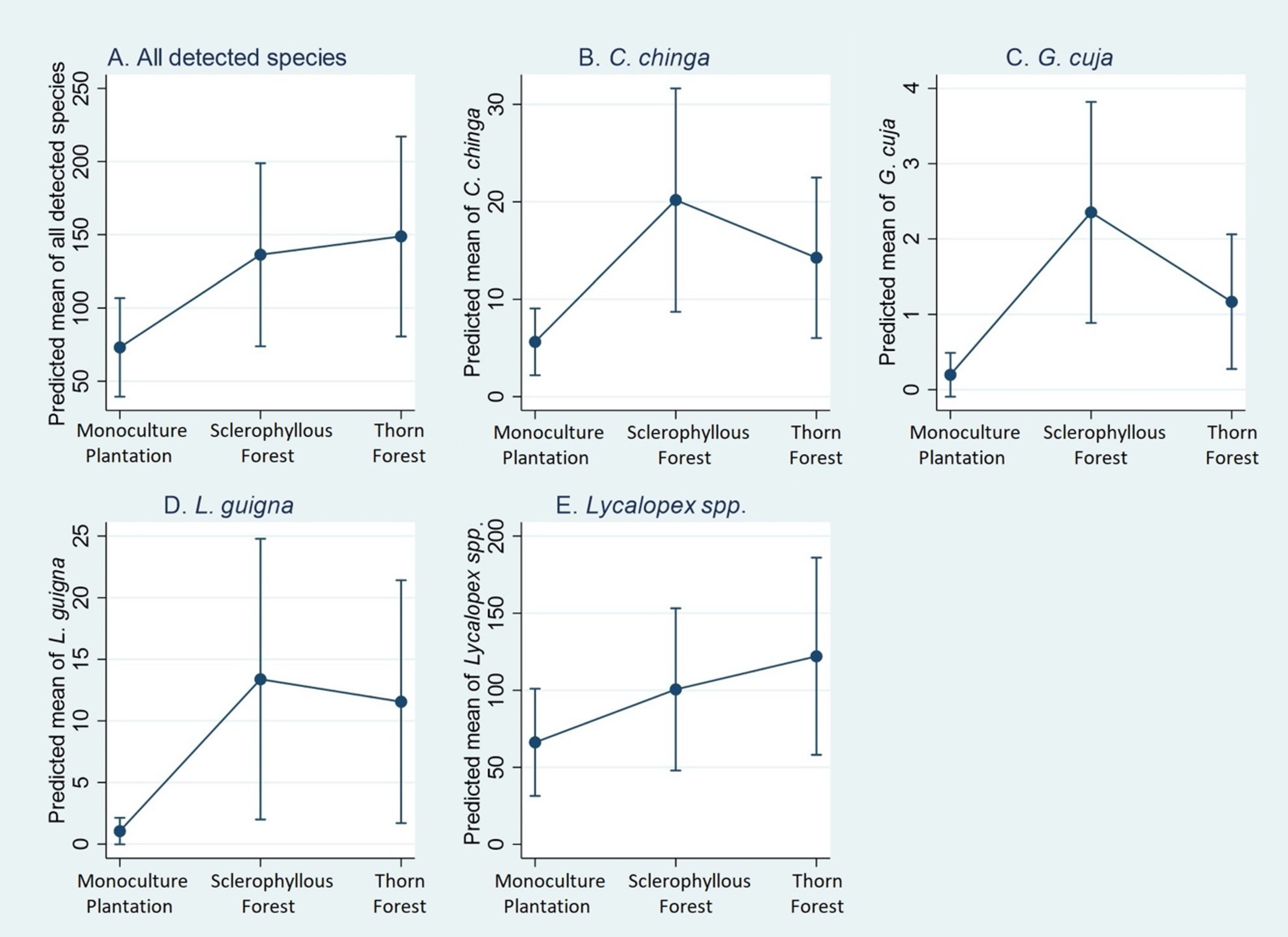
| Variable Name | All Detected Species | C. chinga | G. cuja | L. guigna | Lycalopex spp. |
|---|---|---|---|---|---|
| Intercept | −1.522 | −4.085 | −7.431 | −5.763 | −1.621 |
| (0.235) | (0.311) | (0.749) | (0.522) | (0.268) | |
| <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Location | 0.066 | 0.009 | 0.008 | <0.001 | 0.256 |
| (Monoculture Plantations (MP) (reference) | -- | -- | -- | -- | -- |
| Sclerophyllous Forest (SF) | 0.623 | 1.276 | 2.473 | 2.544 | 0.418 |
| (0.332) | (0.425) | (0.814) | (0.679) | (0.378) | |
| 0.060 | 0.003 | 0.002 | <0.001 | 0.269 | |
| Thorn Forest (TF) | 0.711 | 0.929 | 1.772 | 2.397 | 0.612 |
| (0.332) | (0.428) | (0.845) | (0.680) | (0.379) | |
| 0.032 | 0.030 | 0.036 | <0.001 | 0.106 | |
| Alpha | 0.540 | 0.796 | 0.601 | 1.813 | 0.705 |
| (0.133) | (0.209) | (0.423) | (0.558) | (0.173) | |
| <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Álvarez, D.; Napolitano, C.; Arriagada, G.; Salgado, I.; Cox, S.; Céspedes-Parada, B. Native Carnivore Diversity and Relative Abundance in Landscapes of the Coast Range in Central Chile: Insights for Conservation Decision-Making. Conservation 2023, 3, 379-393. https://doi.org/10.3390/conservation3030026
Ramírez-Álvarez D, Napolitano C, Arriagada G, Salgado I, Cox S, Céspedes-Parada B. Native Carnivore Diversity and Relative Abundance in Landscapes of the Coast Range in Central Chile: Insights for Conservation Decision-Making. Conservation. 2023; 3(3):379-393. https://doi.org/10.3390/conservation3030026
Chicago/Turabian StyleRamírez-Álvarez, Diego, Constanza Napolitano, Gabriel Arriagada, Iván Salgado, Simón Cox, and Belén Céspedes-Parada. 2023. "Native Carnivore Diversity and Relative Abundance in Landscapes of the Coast Range in Central Chile: Insights for Conservation Decision-Making" Conservation 3, no. 3: 379-393. https://doi.org/10.3390/conservation3030026