
Landscape Characteristics Affecting Small Mammal Occurrence in Heterogeneous Olive Grove Agro-Ecosystems


Abstract
:1. Introduction
2. Materials and Methods
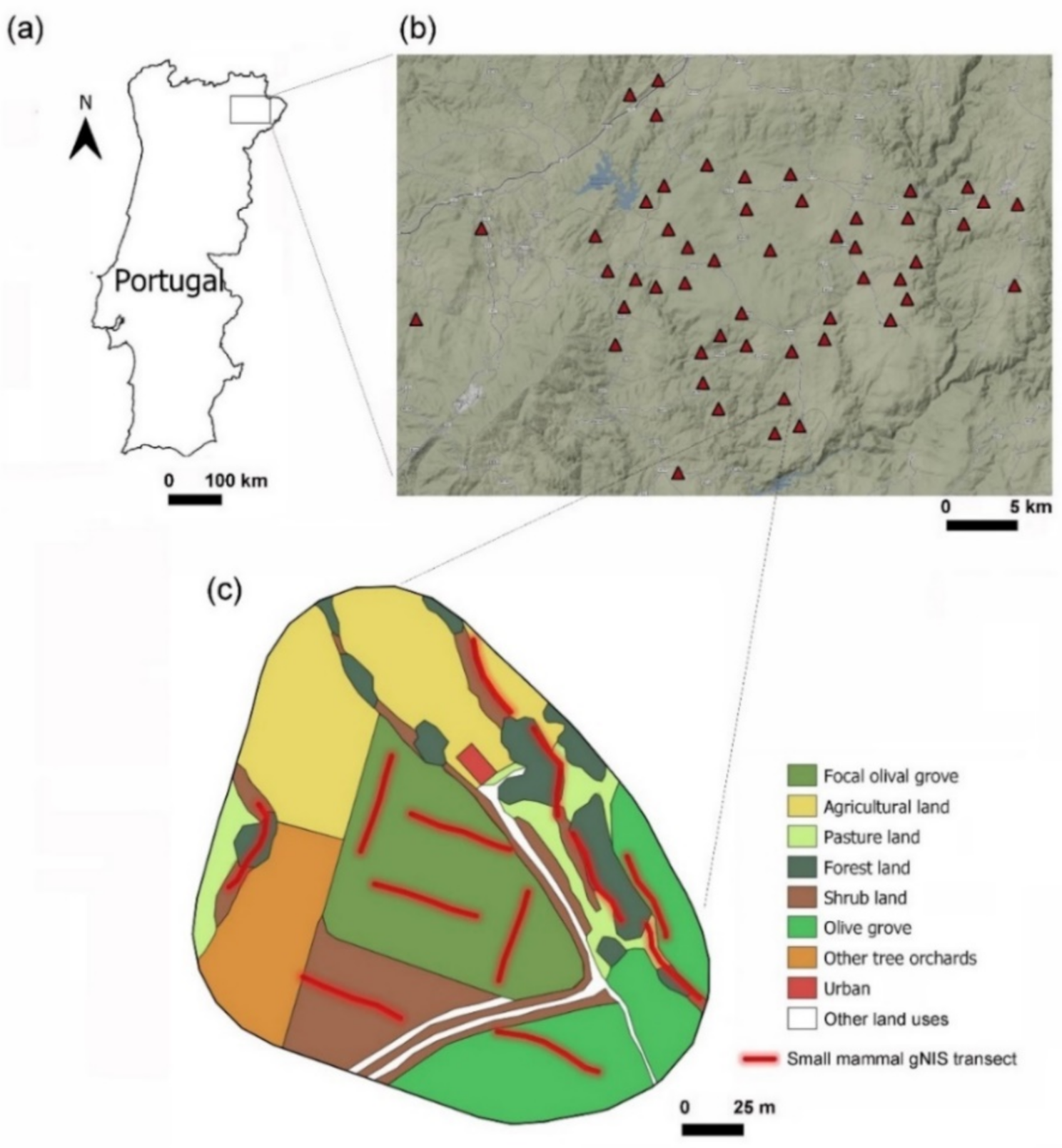
2.1. Study Area and Design
2.2. Small Mammal Genetic Non-Invasive Sampling
2.3. Molecular Analyses
2.4. Bioinformatics
2.5. Landscape Metrics
2.6. Occupancy Modelling
3. Results
3.1. Small Mammal Detection Based on Genetic Non-Invasive Sampling
3.2. Landscape Variability
3.3. Landscape Effects on Microtus cabrerae and Microtus lusitanicus Occupancy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, S.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schildler, D.; Schlesinger, W.H.; Simberloff, D.; Swackharmer, D. Forecasting agriculturally driven environmental change. Science 2001, 292, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoate, C.; Boatman, N.D.; Borralho, R.; Rio Carvalho, C.; de Snoo, G.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; De Fries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Loumou, A.; Giourga, C. Olive groves: “The life and identity of the Mediterranean”. Agric. Hum. Values 2003, 20, 87–95. [Google Scholar] [CrossRef]
- European Commission. LIFE among the Olives: Good Practice in Improving Environmental Performance in the Olive Oil Sector; Office for Official Publications of the European Union: Luxembourg, 2010; Available online: https://publications.europa.eu/en/publication-detail/-/publication/53cd8cd1-272f-4cb8-b7b5-5c100c267f8f (accessed on 31 July 2021).
- Tartaglini, N.; Calabrese, G.; Servadei, L. Ancient olive orchards as high nature value farmland: A shared vision at Euro-Mediterranean level. In A Multi-Scale and Multi-Level Approach for Conservation of Ancient Olive Orchards in the Euro-Mediterranean Region; Posta, L., Lacirignola, C., Mimiola, G., Eds.; CIHEAM—Mediterranean Agronomic Institute of Bari: Bari, Italy, 2012; pp. 25–39. [Google Scholar]
- Herrera, J.M.; Costa, P.; Medinas, D.; Marques, J.T.; Mira, A. Community composition and activity of insectivorous bats in Mediterranean olive farms. Anim. Conserv. 2015, 18, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martínez, J.D.; Cabrera, A.P. The olive monoculture in the south of Spain. Eur. J. Geogr. 2015, 6, 16–29. [Google Scholar]
- Infante-Amate, J.; Villa, I.; Aguilera, E.; Torremocha, E.; Guzmán, G.; Cid, A.; Molina, M.G. The making of olive landscapes in the south of Spain. A history of continuous expansion and intensification. In Biocultural Diversity in Europe. Environmental History; Agnoletti, M., Emanueli, F., Eds.; Springer: Cham, Switzerland, 2016; Volume 5, pp. 157–179. [Google Scholar] [CrossRef]
- Gómez, J.A.; Llewellyn, C.; Basch, G.; Sutton, P.B.; Dyson, J.S.; Jones, C.A. The effects of cover crops and conventional tillage on soil and runoff loss in vineyards and olive groves in several Mediterranean countries. Soil Use Manag. 2011, 27, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Weissteiner, C.J.; Strobl, P.; Sommer, S. Assessment of status and trends of olive farming intensity in EU-Mediterranean countries using remote sensing time series and land cover data. Ecol. Indic. 2011, 11, 601–610. [Google Scholar] [CrossRef]
- Rey, P.J.; Manzaneda, A.J.; Valera, F.; Alcántara, J.M.; Tarifa, R.; Isla, J.; Molina-Pardo, J.; Calvo, G.; Salido, T.; Gutiérrez, E.; et al. Landscape-moderated biodiversity effects of ground herb cover in olive groves: Implications for regional Biodiversity conservation. Agric. Ecosyst. Environ. 2019, 277, 61–73. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Delibes-Mateos, M.; Smith, A.T.; Slobodchikoff, C.N.; Swenson, J.E. The paradox of keystone species persecuted as pests: A call for the conservation of abundant small mammals in their native range. Biol. Conserv. 2011, 144, 1335–1346. [Google Scholar] [CrossRef]
- Fischer, C.; Gayer, C.; Kurucz, K.; Riesch, F.; Tscharntke, T.; Batáry, P. Ecosystem services and disservices provided by small rodents in arable fields: Effects of local and landscape management. J. Appl. Ecol. 2017, 55, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Gliwicz, J.; Taylor, J.R.E. Comparing life histories of shrews and rodents. Acta Theriol. 2002, 47, 185–208. [Google Scholar] [CrossRef]
- Giraudoux, P.; Delattre, P.; Habert, M.; Quéré, J.P.; Deblay, S.; Defaut, R.; Duhamel, R.; Moissenet, M.F.; Salvi, D.; Truchetet, D. Population dynamics of fossorial water vole (Arvicola terrestris scherman): A land use and landscape perspective. Agric. Ecosyst. Environ. 1997, 66, 47–60. [Google Scholar] [CrossRef]
- Aschwanden, J.; Birrer, S.; Jenni, L. Are ecological compensation areas attractive hunting sites for common kestrels (Falco tinnunculus) and long-eared owls (Asio otus)? J. Ornithol. 2005, 146, 279–286. [Google Scholar] [CrossRef]
- Sálek, M.; Kreisinger, J.; Sedlacek, F.; Albrecht, T. Do prey densities determine preferences of mammalian predators for habitat edges in an agricultural landscape? Landsc. Urban Plan. 2010, 98, 86–91. [Google Scholar] [CrossRef]
- Luque-Larena, J.J.; Mougeot, F.; Viñuela, J.; Jareño, D.; Arroyo, L.; Lambin, X.; Arroyo, B. Recent large-scale range expansion and eruption of common vole (Microtus arvalis) outbreaks in NW Spain. Basic Appl. Ecol. 2003, 14, 432–441. [Google Scholar] [CrossRef] [Green Version]
- Renwick, A.R.; Lambin, X. Host-parasite interactions in a fragmented landscape. Int. J. Parasitol. 2013, 43, 27–35. [Google Scholar] [CrossRef]
- Miñarro, M.; Somoano, A.; Ventura, J. Intra-annual continuous reproduction of the apple pest Microtus lusitanicus: Implications for management. Crop Prot. 2017, 96, 164–172. [Google Scholar] [CrossRef]
- Miñarro, M.; Montiel, C.; Dapena, E. Vole pests in apple orchards: Use of presence signs to estimate the abundance of Arvicola terrestris cantabriae and Microtus lusitanicus. J. Pest Sci. 2012, 85, 477–488. [Google Scholar] [CrossRef]
- Pita, R.; Beja, P.; Mira, A. Spatial population structure of the Cabrera vole in Mediterranean farmland: The relative role of patch and matrix effects. Biol. Conserv. 2007, 134, 383–392. [Google Scholar] [CrossRef]
- Dee, L.E.; Cowles, J.; Isbell, F.; Pau, S.; Gaines, S.D.; Reich, P.B. When do ecosystem services depend on rare species? Trends Ecol. Evol. 2019, 34, 746–758. [Google Scholar] [CrossRef]
- Bondi, N.; White, J.G.; Stevens, M.; Cooke, R. A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities. Wildl. Res. 2010, 37, 456–465. [Google Scholar] [CrossRef]
- Beja-Pereira, A.; Oliveira, R.; Alves, P.C.; Schwartz, M.K.; Luikart, G. Advancing ecological understandings through technological transformations in noninvasive genetics. Mol. Ecol. Resour. 2009, 9, 1279–1301. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.M.; Sabino-Marques, H.; Barbosa, S.; Costa, P.; Encarnação, C.; Alpizar-Jara, R.; Pita, R.; Beja, P.; Mira, A.; Searle, J.B.; et al. Genetic non-invasive sampling (gNIS) as a cost-effective tool for monitoring elusive small mammals. Eur. J. Wildl. Res. 2018, 64, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Duarte, F.; Jones, N.; Lúcio, C.; Nunes, A. The reform of the olive oil regime and its impacts on the olive and olive oil sector: A case study in Northern Portugal—Trás-os-Montes. New Medit. 2006, 5, 4–15. [Google Scholar]
- Fernández-Salvador, R. Microtus cabrerae (Thomas, 1906). In Atlas y Libro Rojo de los Mamíferos Terrestres de España; Palomo, L.J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 429–431. [Google Scholar]
- Pita, R.; Mira, A.; Beja, P. Microtus cabrerae (Rodentia: Cricetidae). Mamm. Species 2014, 46, 48–70. [Google Scholar] [CrossRef] [Green Version]
- Cabral, M.; Almeida, J.; Almeida, P.; Dellinger, T.; Ferrand de Almeida, N.; Oliveira, M.; Palmeirim, J.; Queiroz, A.; Rogado, L.; Santos-Reis, M. Livro Vermelho dos Vertebrados de Portugal; Instituto de Conservação da Natureza: Lisbon, Portugal, 2005; p. 659. [Google Scholar]
- Pita, R.; Mira, A.; Beja, P. Conserving the Cabrera vole, Microtus cabrerae, in intensively used Mediterranean landscapes. Agric. Ecosyst. Environ. 2006, 115, 1–5. [Google Scholar] [CrossRef]
- Pita, R.; Mira, A.; Beja, P. Assessing habitat differentiation between coexisting species: The role of spatial scale. Acta Oecol. 2011, 37, 124–132. [Google Scholar] [CrossRef]
- Santos, S.M.; Simões, M.P.; Mathias, M.; Mira, A. Vegetation analysis in colonies of an endangered rodent, the Cabrera vole (Microtus cabrerae), in southern Portugal. Ecol. Res. 2006, 21, 197–207. [Google Scholar] [CrossRef]
- Santos, S.M.; Mathias, M.; Mira, A.; Simões, M.P. Vegetation structure and composition of road verge and meadow sites colonized by Cabrera vole (Microtus cabrerae Thomas). Pol. J. Ecol. 2007, 55, 481–493. [Google Scholar]
- Luque-Larena, J.J.; López, P. Microhabitat use by wild-ranging Cabrera voles Microtus cabrerae as revealed by live trapping. Eur. J. Wildl. Res. 2007, 53, 221–225. [Google Scholar] [CrossRef]
- Santos, S.M.; Lourenço, R.F.; Mathias, M.L.; Mira, A. Spatial and temporal ecology of the Lusitanian pine vole (Microtus lusitanicus) in a Mediterranean polyculture. Anim. Biol. 2010, 60, 209–227. [Google Scholar] [CrossRef]
- Giannoni, S.M.; Borghi, C.E.; Martínez-Rica, J.P. Comparing the burrowing behaviour of the Iberian mole voles (Microtus (Terricola) lusitanicus, M.(T.) pyrenaicus and M.(T.) duodecimcostatus). Mammalia 1993, 57, 483–490. [Google Scholar] [CrossRef]
- Vinhas, A. Microtus lusitanicus (Rato cego) e Microtus duodecimcostatus (Rato cego-mediterrânico), roedores pragas das culturas. Rev. Cienc. Agrar. 1993, 17, 375–382. [Google Scholar]
- Mira, A.; Mathias, M.L. Microtus lusitanicus (Gerbe, 1879). In Atlas y Libro Rojo de los Mamíferos Terrestres de España; Palomo, L.J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 418–421. [Google Scholar]
- Ponsà, M.; Cabré, L.; Ferrando, A.; Bosch, M.; Ventura, J. Approach on the genetic diversity in a Lusitanian pine vole population. Mammalia 2011, 75, 305–309. [Google Scholar] [CrossRef]
- Rodriguez-Pastor, R.; Luque-Larena, J.J.; Lambin, X.; Mougeot, F. “Living on the edge”: The role of field margins for common vole (Microtus arvalis) populations in recently colonised Mediterranean farmland. Agric. Ecosyst. Environ. 2016, 231, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Sukma, H.T.; Stefano, J.D.; Swan, M.; Sitters, H. Mammal functional diversity increases with vegetation structural complexity in two forest types. For. Ecol. Manag. 2019, 433, 85–92. [Google Scholar] [CrossRef]
- Instituto da Conservação da Natureza e das Florestas (ICNF). Parque Natural de Montesinho. Available online: http://www2.icnf.pt/portal/ap/p-nat/pnm (accessed on 1 September 2021).
- Bencatel, J.; Sabino-Marques, H.; Álvares, F.; Moura, A.E.; Barbosa, A.M. Atlas de Mamíferos de Portugal, 2nd ed.; Universidade de Évora: Évora, Portugal, 2019; p. 271. [Google Scholar]
- de Graaff, J.; Duarte, F.; Fleskens, L.; De Figueiredo, T. The future of olive groves on sloping land and ex-ante assessment of cross compliance for erosion control. Land Use Policy 2010, 27, 33–41. [Google Scholar] [CrossRef]
- Sutherland, G.D.; Harestad, A.S.; Price, K.; Lertzman, K.P. Scaling of natal dispersal distances in terrestrial birds and mammals. Conserv. Ecol. 2000, 4, 16. [Google Scholar] [CrossRef]
- Bowman, J.; Jaeger, J.A.G.; Fahrig, L. Dispersal distance of mammals is proportional to home range size. Ecology 2002, 83, 2049–2055. [Google Scholar] [CrossRef]
- Sabino-Marques, H.; Fereira, C.M.; Paupério, J.; Costa, P.; Barbosa, S.; Encarnacao, C.; Alpizar-Jara, R.; Alves, P.C.; Searle, J.B.; Mira, P.; et al. Combining genetic non-invasive sampling with spatially explicit capture-recapture models for density estimation of a patchily distributed small mammal. Eur. J. Wildl. Res. 2018, 64, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Martins, F.M.S.; Galhardo, M.; Filipe, A.F.; Teixeira, A.; Pinheiro, P.; Paupério, J.; Alves, P.C.; Beja, P. Have the cake and eat it: Optimizing nondestructive DNA metabarcoding of macroinvertebrate samples for freshwater biomonitoring. Mol. Ecol. Resour. 2019, 19, 863–876. [Google Scholar] [CrossRef]
- Barbosa, S.; Paupério, J.; Searle, J.B.; Alves, P.C. Genetic identification of Iberian rodent species using both mitochondrial and nuclear loci: Application to noninvasive sampling. Mol. Ecol. Resour. 2013, 13, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Rosenbom, S.; Monteiro, R.; O‘Rourke, S.M.; Beja-Pereira, A. Improving DNA 515 quality extracted from fecal samples—A method to improve DNA yield. Eur. J. Wildl. Res. 2017, 63, 1–7. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef] [PubMed]
- Riaz, T.; Shehzad, W.; Viari, A.; Pompanon, F.; Taberlet, P.; Coissac, E. ecoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucl. Acids Res. 2011, 39, e145. [Google Scholar] [CrossRef]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. Obitools: A unix-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2008, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.; Lewin-Koh, N. Maptools: Tools for Handling Spatial Objects. R Package. Available online: https://cran.r-project.org/web/packages/maptools/index.html (accessed on 30 September 2021).
- Bivand, R.; Keitt, T.; Rowlingson, B.; Pebesma, E.; Sumner, M.; Hijmans, R.; Rouault, E. Rgdal: Bindings for the Geospatial Data Abstraction Library. R Package. Available online: https://cran.r-project.org/web/packages/rgdal/index.html (accessed on 30 September 2021).
- Wickham, H.; Francois, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. R Package. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 30 September 2021).
- Sherrill-Mix, S. Taxonomizr: Functions to Work with NCBI Accessions and Taxonomy. R Package. Available online: https://CRAN.R-project.org/package=taxonomizr (accessed on 30 September 2021).
- Müller, K.; Wickham, H. Tibble: Simple Data Frames. R Package. Available online: https://CRAN.R-project.org/package=tibble (accessed on 30 September 2021).
- Allaire, J. RStudio: Integrated Development Environment for R; RStudio Team: Boston, MA, USA, 2012; Volume 770, p. 394. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 14 August 2021).
- QGIS Development Team. QGIS Geographic Information System. QGIS Association. Available online: https://www.qgis.org/en/site/ (accessed on 30 July 2021).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package. Available online: https://CRAN.R-project.org/package=raster (accessed on 2 August 2021).
- Sciaini, M.; Fritsch, M.; Scherer, C.; Simpkins, C.E. NLMR and Landscapetools: An Integrated Environment for Simulating and Modifying Neutral Landscape Models in R. Methods Ecol. Evol. 2018, 9, 2240–2248. [Google Scholar] [CrossRef] [Green Version]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.J.; Gowda, S.V.; Gururaja, K.V.; Sudhira, H.S. Effect of Landscape Metrics on Varied Spatial Extents of Bangalore, India. Asian J. Geoinformatics 2012, 12, 1–11. [Google Scholar]
- Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droedge, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carre, G.; García Marquéz, J.R.; Gruber, B.; Lafourcade, B.; Leitão, P.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2012, 36, 27–46. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package. Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 30 October 2021).
- Legendre, P. Spatial autocorrelation: Trouble or new paradigm? Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Montoya, D.; Gaba, S.; de Mazancourt, C.; Bretagnolle, V.; Loreau, M. Reconciling biodiversity conservation, food production and farmers’ demand in agricultural landscapes. Ecol. Modell. 2020, 416, 108889–108909. [Google Scholar] [CrossRef] [PubMed]
- Millán de la Peña, N.; Butet, A.; Delettre, Y.; Paillat, G.; Morant, P.; Le Du, L.; Burel, F. Response of the small mammal community to changes in western French agricultural landscapes. Landsc. Ecol. 2003, 18, 265–278. [Google Scholar] [CrossRef]
- Pita, R.; Lambin, X.; Mira, A.; Beja, P. Hierarchical spatial segregation of two Mediterranean vole species: The role of patch-network structure and matrix composition. Oecologia 2016, 182, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, B.J.; Fox, M.D. Factors determining mammal species richness on habitat islands and isolates: Habitat diversity, disturbance, species interactions and guild assembly rules. Glob. Ecol. Biogeogr. 2000, 9, 19–37. [Google Scholar] [CrossRef]
- Cramer, M.J.; Willig, M.R. Habitat heterogeneity, habitat associations, and rodent species diversity in a sand–shinnery-oak landscape. J. Mammal. 2002, 83, 743–753. [Google Scholar] [CrossRef]
- Rigaux, P.; Vaslin, M.; Noblet, J.F.; Amori, G.; Muñoz, L.J.P. Arvicola sapidus. In The IUCN Red List of Threatened Species; e.T2150A9290712. 2008. Available online: https://www.iucnredlist.org/species/2150/9290712 (accessed on 10 November 2021).
- Paupério, J.; Herman, J.S.; Melo-Ferreira, J.; Jaarola, M.; Alves, P.C.; Searle, J.B. Cryptic speciation in the field vole: A multilocus approach confirms three highly divergent lineages in Eurasia. Mol. Ecol. 2012, 21, 6015–6032. [Google Scholar] [CrossRef]
- Balestrieri, A.; Gazzola, A.; Formenton, G.; Canova, L. Long-term impact of agricultural practices on the diversity of small mammal communities: A case study based on owl pellets. Environ. Monit. Assess. 2019, 191, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Morgado, R.; Santana, J.; Porto, M.; Sánchez-Oliver, J.S.; Reino, L.; Herrera, J.M.; Rego, F.; Beja, P.; Moreira, F. A Mediterranean silent spring? The effects of olive farming intensification on breeding bird communities. Agric. Ecosyst. Environ. 2020, 288, 106694. [Google Scholar] [CrossRef]
- Castro, J.; Tortosa, F.S.; Carpio, A.J. Structure of canopy and ground-dwelling arthropod communities in olive orchards is determined by the type of soil cover. Eur. J. Entomol. 2021, 118, 159–170. [Google Scholar] [CrossRef]
- Tarifa, R.; Martínez-Núñez, C.; Valera, F.; González-Varo, J.P.; Salido, T.; Rey, P.J. Agricultural intensification erodes taxonomic and functional diversity in Mediterranean olive groves by filtering out rare species. J. Appl. Ecol. 2021, 58, 2266–2276. [Google Scholar] [CrossRef]
- Paz, D.B.; Henderson, K.; Loreau, M. Agricultural land use and the sustainability of social-ecological systems. Ecol. Modell. 2020, 437, 109312. [Google Scholar] [CrossRef] [PubMed]
- Serafini, V.N.; Priotto, J.W.; Gomez, M.D. Effects of agroecosystem landscape complexity on small mammals: A multi-species approach at different spatial scales. Landsc. Ecol. 2019, 34, 1117–1129. [Google Scholar] [CrossRef]
- Moreira, F.; Herrera, J.M.; Beja, P. Making olive oil sustainable. Science 2019, 365, 873. [Google Scholar] [CrossRef]
- Krishna, V.; Drucker, A.; Pascual, U.; Raghu, P.E.D.; King, I.O. Estimating compensation payments for on-farm conservation of agricultural biodiversity in developing countries. Ecol. Econ. 2013, 87, 110–123. [Google Scholar] [CrossRef]
- Proença-Ferreira, A.; Ferreira, C.; Leitão, I.; Paupério, J.; Sabino-Marques, H.; Barbosa, S.; Lambin, X.; Alves, P.C.; Beja, P.; Moreira, F.; et al. Drivers of survival in a small mammal of conservation concern: An assessment using extensive genetic non-invasive sampling in fragmented farmland. Biol. Conserv. 2019, 230, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Reiners, T.E.; Encarnação, J.A.; Wolters, V. An optimized hair trap for non-invasive genetic studies of small cryptic mammals. Eur. J. Wildl. Res. 2011, 57, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Van Veen, F.J.; Memmott, J.; Godfray, H.C.J. Indirect effects, apparent competition and biological control. In Trophic and Guild Interactions in Biological Control; Brodeur, J., Boivin, G., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 145–169. [Google Scholar]
- Waits, L.P. Using noninvasive genetic sampling to detect and estimate abundance of rare wildlife species. In Sampling Rare or Elusive Species: Concepts, Designs, and Techniques for Estimating Population Parameters; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 211–228. [Google Scholar]
- Benedek, A.M.; Sîrbu, I. Responses of small mammal communities to environment and agriculture in a rural mosaic landscape. Mamm. Biol. 2018, 90, 55–65. [Google Scholar] [CrossRef]


{kind=link}
| Species | p Submodel | AICc | ΔAICc | Wt-AICc |
|---|---|---|---|---|
| Microtus cabrerae | ~1 | 553.916 | 24.783 | 0.00 |
| ~TransType | 529.133 | 1.00 | ||
| Microtus lusitanicus | ~1 | 268.363 | 7.040 | 0.03 |
| ~TransType | 261.323 | 0.97 | ||
| Apodemus sylvaticus | ~1 | 750.621 | 32.786 | 0.00 |
| ~TransType | 717.835 | 1.00 | ||
| Mus spretus | ~1 | 467.628 | 7.495 | 0.02 |
| ~TransType | 460.133 | 0.98 | ||
| Crocidura russula | ~1 | 486.666 | 18.523 | 0.00 |
| ~TransType | 468.144 | 1.00 |
| Species | Mean (95%CI) p (within Olive Groves) | Mean (95%CI) p (Surrounding Habitats) | Mean (95%CI) Psi |
|---|---|---|---|
| Microtus cabrerae | 0.037 (0.012–0.108) | 0.267 (0.223–0.317) | 0.765 (0.611–0.871) |
| Microtus lusitanicus | 0.029 (0.004–0.181) | 0.219 (0.158–0.296) | 0.366 (0.236–0.519) |
| Apodemus sylvaticus | 0.085 (0.045–0.156) | 0.347 (0.304–0.392) | 0.984 (0.654–0.999) |
| Mus spretus | 0.066 (0.030–0.141) | 0.163 (0.126–0.210) | 0.868 (0.635–0.961) |
| Crocidura russula | 0.019 (0.005–0.072) | 0.158 (0.129–0.193) | 1.000 (0.000–1.000) 1 |
| Covariable (Units) | Code | Mean ± se | Range |
|---|---|---|---|
| Cover metrics | |||
| Olive grove cover (%) | OliveCover | 53.16 ± 3.26 | 11.5–100 |
| Pasture land cover (%) | PastCover | 15.84 ± 1.82 | 0–47.6 |
| Shrub cover (%) | ShrubCover | 13.28 ± 2.08 | 0–58.6 |
| Agricultural land cover (%) | AgricCover | 7.01 ± 1.30 | 0–37.7 |
| Forest cover (%) | ForestCover | 6.17 ± 1.75 | 0–63.8 |
| Edge density metrics | |||
| Olive edge density /m/ha) | OliveEdgeD | 202.05 ± 11.95 | 0–443 |
| Pasture land edge density /m/ha) | PastEdgeD | 114.16 ± 12.25 | 0–353 |
| Shrub edge density /m/ha) | ShrubEdgeD | 124.60 ± 16.65 | 0–483 |
| Agricultural land edge density (m/ha) | AgricEdgeD | 64.48 ± 11.21 | 0–303 |
| Forest edge density /m/ha) | ForestEdgeD | 46.37 ± 9.99 | 0–293 |
| Patch density metrics | |||
| Olive patch density (number/ha) | OlivePatchD | 65.18 ± 5.34 | 21.7–189 |
| Pasture patch density (number/ha) | PastPatchD | 61.70 ± 7.05 | 0–218 |
| Shrub patch density (number/ha) | ShrubPatchD | 60.89 ± 5.56 | 0–203 |
| Agricultural patch density (number/ha) | AgricPatchD | 35.94 ± 5.56 | 0–172 |
| Forest patch density (number/ha) | ForestPatchD | 20.32 ± 4.27 | 0–110 |
| Aggregation metrics | |||
| Olive normalized landscape shape index | OliveNLSI | 0.04 ± 0.02 | 0–1.00 |
| Pasture normalized landscape shape index | PastNLSI | 0.02 ± 0.01 | 0–0.27 |
| Shrub normalized landscape shape index | ShrubNLSI | 0.02 ± 0.00 | 0–0.06 |
| Agricultural normalized landscape shape index | AgricNLSI | 0.01 ± 0.00 | 0–0.10 |
| Forest normalized landscape shape index | ForestNLSI | 0.01 ± 0.00 | 0–0.07 |
| Diversity metrics | |||
| Number of habitat types | PatchRichness | 4.67 ± 0.20 | 1–7 |
| Shannon diversity index | ShanDiv | 1.04 ± 0.05 | 0–1.73 |
| Shannon evenness index | ShanEvn | 0.68 ± 0.03 | 0–0.91 |
| Species | Psi Submodel | AICc | ΔAICc | Wt-AICc |
|---|---|---|---|---|
| Microtus cabrerae | ~AgricEdgeD + PastNLSI | 522.87 | 0 | 0.55 |
| ~AgricEdgeD + AgricNLSI + PastNLSI | 525.22 | 2.35 | 0.17 | |
| ~AgricNLSI + PastNLSI | 525.95 | 3.08 | 0.12 | |
| ~AgricEdgeD | 527.25 | 4.38 | 0.06 | |
| ~AgricNLSI | 528.08 | 5.21 | 0.04 | |
| ~PastNLSI | 528.35 | 5.48 | 0.04 | |
| ~AgricEdgeD + AgricNLSI | 529.22 | 6.35 | 0.02 | |
| Microtus lusitanicus | ~PastPatchD + OliveNLSI | 258.1 | 0 | 0.24 |
| ~PastPatchD | 258.28 | 0.18 | 0.22 | |
| ~PastPatchD + ForestNLSI | 258.7 | 0.6 | 0.18 | |
| ~PastPatchD + OliveNLSI + ForestNLSI | 259.31 | 1.21 | 0.13 | |
| ~OliveNLSI | 259.57 | 1.47 | 0.11 | |
| ~OliveNLSI + ForestNLSI | 260.52 | 2.41 | 0.07 | |
| ~ForestNLSI | 261.2 | 3.1 | 0.05 |
| Species | Parameter | Covariable | Effect Size | 5th, 95th Percentiles |
|---|---|---|---|---|
| Microtus cabrerae | p | TransType:SurroundHabitat | 2.26 | 1.27, 3.25 |
| Psi | AgricEdgeD | 1.68 | 0.31, 3.05 | |
| Psi | PastNLSI | −2.23 | −5.00, 0.54 | |
| Microtus lusitanicus | p | TransType:SurroundHabitat | 2.24 | 0.54, 3.93 |
| Psi | PastPatchD | 0.75 | 0.07, 1.43 | |
| Psi | OliveNLSI | −7.89 | −17.8, 2.03 | |
| Psi | ForestNLSI | 0.44 | −0.15, 1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barão, I.; Queirós, J.; Vale-Gonçalves, H.; Paupério, J.; Pita, R. Landscape Characteristics Affecting Small Mammal Occurrence in Heterogeneous Olive Grove Agro-Ecosystems. Conservation 2022, 2, 51-67. https://doi.org/10.3390/conservation2010005
Barão I, Queirós J, Vale-Gonçalves H, Paupério J, Pita R. Landscape Characteristics Affecting Small Mammal Occurrence in Heterogeneous Olive Grove Agro-Ecosystems. Conservation. 2022; 2(1):51-67. https://doi.org/10.3390/conservation2010005
Chicago/Turabian StyleBarão, Isabel, João Queirós, Hélia Vale-Gonçalves, Joana Paupério, and Ricardo Pita. 2022. "Landscape Characteristics Affecting Small Mammal Occurrence in Heterogeneous Olive Grove Agro-Ecosystems" Conservation 2, no. 1: 51-67. https://doi.org/10.3390/conservation2010005