
Morpho-Physiological Assessment of Tomato and Bell Pepper in Response to Nutrient Restriction


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Summary of Statistical Analysis
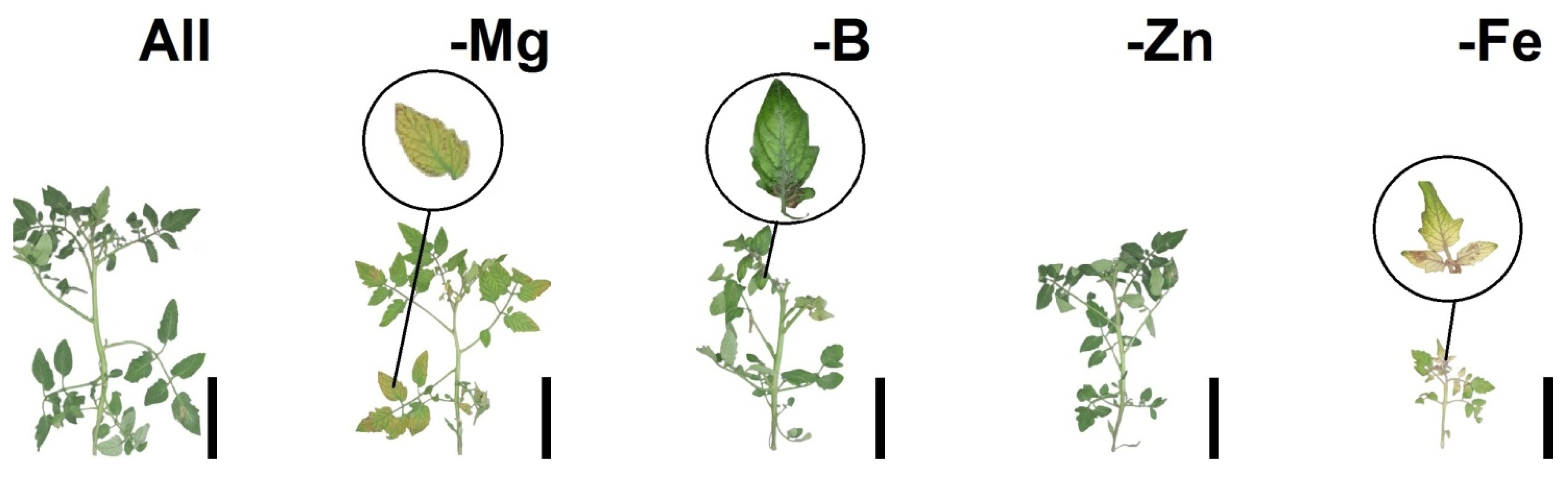
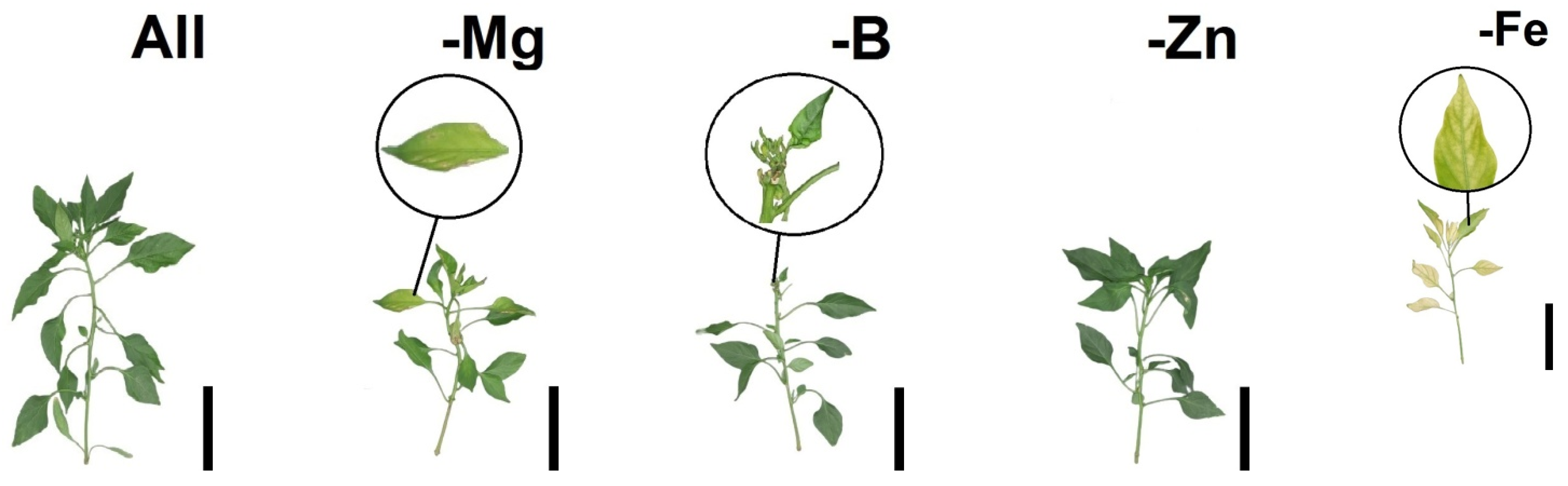
2.2. Morpho-Physiological Responses of Tomatoes and Bell Peppers to Nutrient Suppression
2.3. Changes in Biometric and Production Components as a Function of Nutrients Suppression in Tomatoes and Bell Peppers
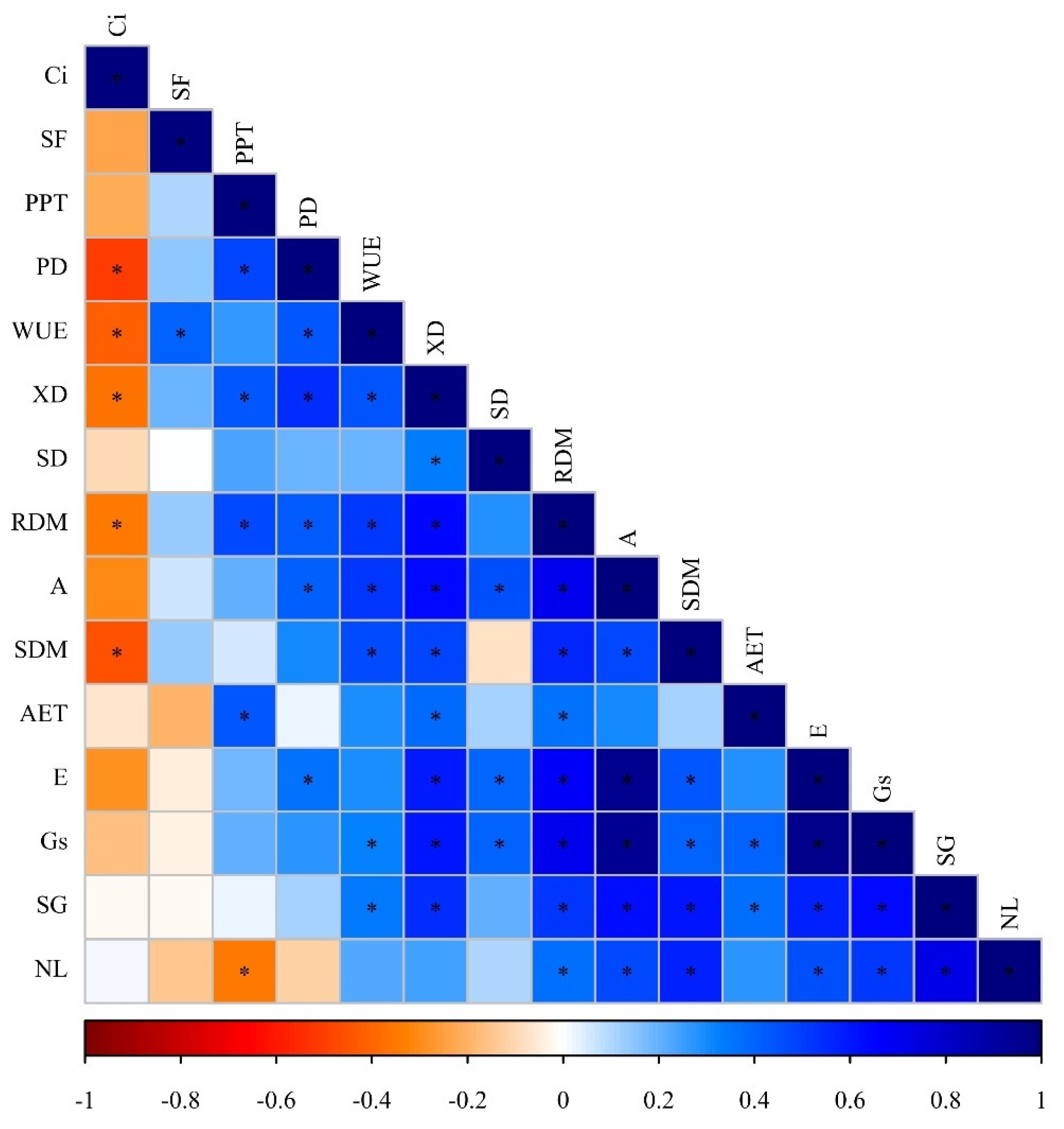
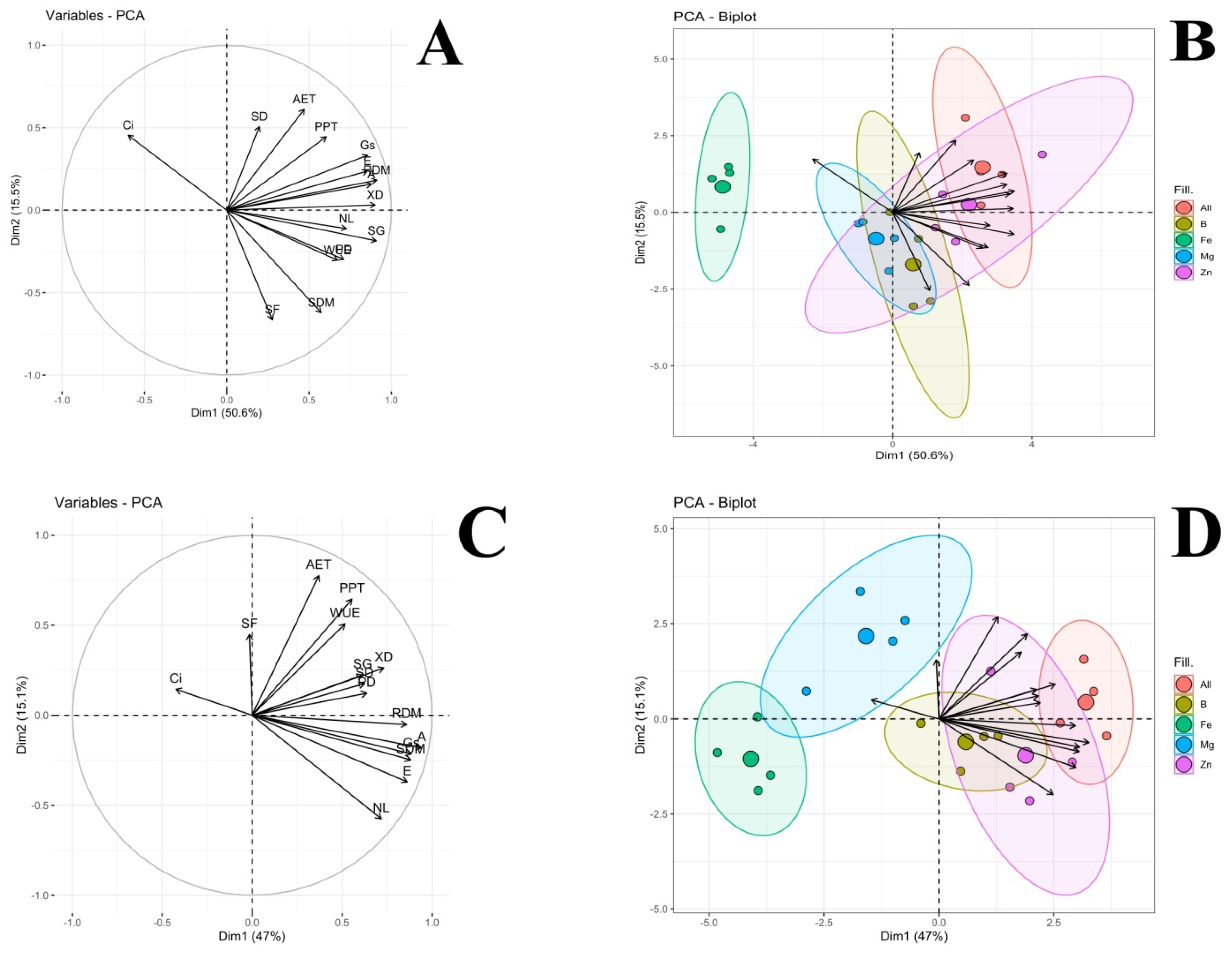
2.4. Nutritional Deficiency Symptoms, Pearson’s Correlation (Heatmap) and Principal Component Analysis (PCA) as a Function of Plant Species and Nutrient Suppression
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Treatments
4.2. Growth Parameters—Biometric Components
4.3. Physiological Parameters—Gas Exchange
4.4. Morphological Parameters
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brandão Filho, J.U.T.; Freitas, P.S.L.; Berian, L.O.S.; Goto, R. Hortaliças-Fruto; EDUEM: Maringá, Brazil, 2018; 535p. [Google Scholar] [CrossRef]
- Ali, M.Y.; Sina, A.A.I.; Khandker, S.S.; Neesa, L.; Tanvir, E.M.; Kabir, A.; Khalil, M.I.; Gan, S.H. Nutritional Composition and Bioactive Compounds in Tomatoes and Their Impact on Human Health and Disease: A Review. Foods 2021, 10, 45. [Google Scholar] [CrossRef]
- Franczuk, J.; Tartanus, M.; Rosa, R.; Zaniewicz-Bajkowska, A.; Dębski, H.; Andrejiová, A.; Dydiv, A. The Effect of Mycorrhiza Fungi and Various Mineral Fertilizer Levels on the Growth, Yield, and Nutritional Value of Bell Pepper (Capsicum annuum L.). Agriculture 2023, 13, 857. [Google Scholar] [CrossRef]
- Guilherme, R.; Reboredo, F.; Guerra, M.; Ressurreição, S.; Alvarenga, N. Elemental Composition and Some Nutritional Parameters of Bell Pepper from Organic and Conventional Agriculture. Plants 2020, 9, 863. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Cheraghi, M.; Motesharezadeh, B.; Alikhani, H.A.; Mousavi, S.M. Optimal management of plant nutrition in tomato (Lycopersicon esculent Mill) by using biologic, organic and inorganic fertilizers. J. Plant Nutr. 2023, 46, 1560–1579. [Google Scholar] [CrossRef]
- Cheraghi, M.; Motesharezadeh, B.; Alikhani, H.A. Nutritional and morpho-physiological responses of tomato plant (Lycopersicon esculentum Mill) affected by biological and chemical fertilizers. Iran. J. Soil Water Res. 2020, 51, 59–74. (In Persian) [Google Scholar] [CrossRef]
- Singh, P.; Singh, D.; Singh, A.K.; Singh, B.; Singh, T. Growth and yield of tomato grown under organic and inorganic nutrient management. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 365–375. [Google Scholar] [CrossRef]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. Biometals 2002, 15, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Lisboa, L.A.M.; Cavichioli, J.C.; Vitorino, R.; Figueiredo, P.A.M.; da Silva Viana, R. Nutrient suppression in passion fruit species: An approach to leaf development and morphology. Colloq. Agrar. 2021, 17, 89–102. [Google Scholar] [CrossRef]
- Fiorentini, D.; Cappadone, C.; Farruggia, G.; Prata, C. Magnesium: Biochemistry, nutrition, detection, and social impact of diseases linked to its deficiency. Nutrients 2021, 13, 1136. [Google Scholar] [CrossRef]
- Ishfaq, M.; Zhong, Y.; Wang, Y.; Li, X. Magnesium Limitation Leads to Transcriptional Down-Tuning of Auxin Synthesis, Transport, and Signaling in the Tomato Root. Front. Plant Sci 2021, 12, 802399. [Google Scholar] [CrossRef]
- Amirahmadi, E.; Ghorbani, M.; Moudrý, J.; Konvalina, P.; Kopecký, M. Impacts of Environmental Factors and Nutrients Management on Tomato Grown under Controlled and Open Field Conditions. Agronomy 2023, 13, 916. [Google Scholar] [CrossRef]
- Sharma, H.; Sharma, A.; Yashvika; Sidhu, S.; Upadhyay, S.K. A glimpse of boron transport in plants. In Cation Transporters in Plants; Academic Press: Cambridge, MA, USA, 2022; pp. 281–306. [Google Scholar] [CrossRef]
- Kohli, S.K.; Kaur, H.; Khanna, K.; Handa, N.; Bhardwaj, R.; Rinklebe, J.; Ahmad, P. Boron in plants: Uptake, deficiency and biological potential. Plant Growth Regul. 2022, 97, 267–282. [Google Scholar] [CrossRef]
- Xu, W.; Wang, P.; Yuan, L.; Chen, X.; Hu, X. Effects of Application Methods of Boron on Tomato Growth, Fruit Quality and Flavor. Horticulturae 2021, 7, 223. [Google Scholar] [CrossRef]
- Clemens, S. The cell biology of zinc. J. Exp. Bot. 2021, 73, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Ohnishi, M.; Maekawa, S.; Wada, S.; Ifuku, K.; Miyake, C. Evaluating the Oxidation Rate of Reduced Ferredoxin in Arabidopsis thaliana Independent of Photosynthetic Linear Electron Flow: Plausible Activity of Ferredoxin-Dependent Cyclic Electron Flow around Photosystem I. Int. J. Mol. Sci. 2023, 24, 12145. [Google Scholar] [CrossRef] [PubMed]
- Stutz, S.S.; Hanson, D.T. What is the fate of xylem-transported CO2 in Kranz-type C4 plants? New Phytol. 2019, 223, 1241–1252. [Google Scholar] [CrossRef]
- Guo, B.; Liu, J.; Liu, C.; Lin, Y.; Li, H.; Zhu, D.; Zhang, Q.; Chen, X.; Qiu, G.; Fu, Q. Shade and iron plaque of Sesbania affect cadmium accumulation in rice: A new strategy for safe production in contaminated soil. Environ. Technol. Innov. 2023, 29, 102964. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Greer, D.H.; Moroni, F.J.; Baby, T. Potassium and Magnesium Mediate the Light and CO2 Photosynthetic Responses of Grapevines. Biology 2020, 9, 144. [Google Scholar] [CrossRef]
- Inoue, S.; Hayashi, M.; Huang, S.; Yokosho, K.; Gotoh, E.; Ikematsu, S.; Okumura, M.; Suzuki, T.; Kamura, T.; Kinoshita, T. A tonoplast localized magnesium transporter is crucial for stomatal opening in Arabidopsis under high Mg2+ conditions. New Phytol. 2022, 236, 864–877. [Google Scholar] [CrossRef]
- Santos, M.S.; Sanglard, L.M.P.V.; Martins, S.C.V.; Barbosa, M.L.; Melo, D.C.; Gonzaga, W.F.; Damatta, F.M. Silicon alleviates the impairments of iron toxicity on the rice photosynthetic performance via alterations in leaf diffusive conductance with minimal impacts on carbon metabolism. Plant Physiol. Biochem. 2019, 143, 275–285. [Google Scholar] [CrossRef]
- Kouas, S.; Slatni, T.; Chihaoui, S.; Abdelly, C.; Mhadhbi, H. Differential behavior of Medicago truncatula to calcareous soil is explained by modulation of stomatal responses, antioxidant activity, and iron use efficiency. Arab. J. Geosci. 2021, 14, 1–12. [Google Scholar] [CrossRef]
- Alrashidi, A.A.; Alhaithloul, H.A.S.; Soliman, M.H.; Attia, M.S.; Elsayed, S.M.; Sadek, A.M.; Fakhr, M.A. Role of calcium and magnesium on dramatic physiological and anatomical responses in tomato plants. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12614. [Google Scholar] [CrossRef]
- Viswanath, K.K.; Varakumar, P.; Pamuru, R.R.; Basha, S.J.; Mehta, S.; Rao, A.D. Plant Lipoxygenases and Their Role in Plant Physiology. J. Plant Biol. 2020, 63, 83–95. [Google Scholar] [CrossRef]
- Assefa, T.; Zhang, J.; Chowda-Reddy, R.V.; Lauter, A.N.M.; Singh, A.; O’rourke, J.A.; Graham, M.A.; Singh, A.K. Deconstructing the genetic architecture of iron deficiency chlorosis in soybean using genome-wide approaches. Bmc Plant Biol. 2020, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.; Bacaicoa, E.; Rivero, M.; Zamarreño, Á.M.; García-Mina, J.M. Complementary Evaluation of Iron Deficiency Root Responses to Assess the Effectiveness of Different Iron Foliar Applications for Chlorosis Remediation. Front. Plant Sci. 2018, 9, 351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The Adaptive Mechanism of Plants to Iron Deficiency via Iron Uptake, Transport, and Homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef] [PubMed]
- Furlani, P.R. Instruções Para o Cultivo de Hortaliças de Folhas Pela Técnica de Hidroponia—NFT; Instituto Agronômico de Campinas: Campinas, Brazil, 1997; p. 30. [Google Scholar]
- Segatto, F.B.; Bisognin, D.A.; Benedetti, M.; Costa, L.C.; Rampelotto, M.V.; Nicoloso, F.T. A technique for the anatomical study of potato leaf epidermis. Ciência Rural 2004, 34, 1597–1601. [Google Scholar] [CrossRef]
- Castro, E.M.; Pereira, F.J.; Paiva, R. Histologia Vegetal: Estrutura e Função de Órgãos Vegetativos; UFLA: Lavras, Brazil, 2009; p. 234. [Google Scholar]
- Galindo, F.S.; Rodrigues, W.L.; Fernandes, G.C.; Boleta, E.H.M.; Jalal, A.; Rosa, P.A.L.; Buzetti, S.; Lavres, J.; Teixeira Filho, M.C.M. Enhancing agronomic efficiency and maize grain yield with Azospirillum brasilense inoculation under Brazilian savannah conditions. Eur. J. Agron. 2022, 134, 126471. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org/ (accessed on 3 December 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Value | A | E | Gs | Ci | WUE |
| Vegetable (V) | 0.0911 ns | 0.0665 ns | 0.0039 ** | 0.0112 * | 0.6665 ns |
| Nutrient (N) | 0.0001 ** | 0.0001 ** | 0.0001 ** | 0.0112 * | 0.0005 ** |
| V × N | 0.0893 ns | 0.2140 ns | 0.1462 ns | 0.6855 ns | 0.8408 ns |
| p-value | PD | XD | PPT | AET | SD |
| Vegetable (V) | 0.0001 ** | 0.7194 ns | 0.0001 ** | 0.0168 * | 0.7227 ns |
| Nutrient (N) | 0.0001 ** | 0.0001 ** | 0.0001 ** | 0.0001 ** | 0.0213 * |
| V × N | 0.3990 ns | 0.1668 ns | 0.0265 * | 0.3176 ns | 0.3519 ns |
| p-value | SF | SG | NL | SDM | RDM |
| Vegetable (V) | 0.1285 ns | 0.0001 ** | 0.0001 ** | 0.0455 * | 0.0172 * |
| Nutrient (N) | 0.3175 ns | 0.0001 ** | 0.0001 ** | 0.0001 ** | 0.0001 ** |
| V × N | 0.2174 ns | 0.1484 ns | 0.0001 ** | 0.0019 ** | 0.0068 ** |
| Physiological Parameters | |||||
| Vegetable | A | E | Gs | Ci | WUE |
| Tomato (average over all treatments) | 12.0 a | 2.38 a | 0.173 a | 254 a | 4.81 a |
| Bell pepper (average over all treatments) | 10.5 a | 2.09 a | 0.129 b | 231 b | 4.93 a |
| Nutrients | |||||
| All | 16.8 a | 3.41 a | 0.275 a | 225 b | 4.95 a |
| -Mg | 6.1 c | 1.13 c | 0.075 c | 259 a | 5.29 a |
| -Zn | 17.8 a | 2.28 b | 0.250 a | 254 a | 5.42 a |
| -B | 11.7 b | 3.29 a | 0.117 b | 199 b | 5.25 a |
| -Fe | 3.84 c | 1.07 c | 0.037 c | 276 a | 3.45 b |
| C.V. (%) | 24.2 | 21.4 | 29.4 | 10.8 | 18.1 |
| Average mean | 11.2 | 2.23 | 0.151 | 243 | 4.87 |
| Morphological parameters | |||||
| Vegetable | PD | XD | AET | SD | SF |
| Tomato (average over all treatments) | 3.17 b | 13.0 a | 23.0 a | 178 a | 1.74 a |
| Bell pepper (average over all treatments) | 4.40 a | 13.3 a | 19.5 b | 173 a | 1.85 a |
| Nutrients | |||||
| All | 4.39 a | 17.8 a | 29.0 a | 193 a | 1.73 a |
| -Mg | 3.30 b | 13.0 b | 24.0 b | 140 b | 1.86 a |
| -Zn | 4.13 a | 14.3 b | 18.8 c | 212 a | 1.87 a |
| -B | 4.67 a | 15.2 b | 17.8 c | 178 a | 1.84 a |
| -Fe | 2.44 b | 5.5 c | 16.4 c | 156 b | 1.69 a |
| C.V. (%) | 22.3 | 17.9 | 20.5 | 25.0 | 11.6 |
| Average mean | 3.79 | 13.2 | 21.2 | 176 | 1.80 |
| PPT (µm) | |||||
|---|---|---|---|---|---|
| Vegetable/Nutrient | All | -Mg | -Zn | -B | -Fe |
| Tomato | 7.96 bA | 5.35 bB | 4.65 bB | 4.58 bB | 5.23 aB |
| Bell pepper | 15.2 aA | 11.6 aB | 8.26 aC | 7.49 aC | 3.62 aD |
| Vegetable | SG | NL |
|---|---|---|
| Tomato (average over all treatments) | 11.1 a | 38.4 a |
| Bell pepper (average over all treatments) | 5.88 b | 18.0 b |
| Nutrients | ||
| All | 12.6 a | 30.8 a |
| -Mg | 9.47 b | 29.0 a |
| -Zn | 10.4 b | 35.5 a |
| -B | 8.72 b | 28.7 a |
| -Fe | 1.30 c | 16.8 b |
| C.V. (%) | 28.4 | 19.8 |
| Average Mean | 8.53 | 28.2 |
| NL | |||||
| Vegetable/Nutrient | All | -Mg | -Zn | -B | -Fe |
| Tomato | 40.5 aB | 48.0 aA | 47.0 aA | 35.7 aB | 20.7 aC |
| Bell pepper | 21.2 bA | 10.0 bB | 24.0 bA | 21.7 bA | 13.0 aB |
| SDM (g) | |||||
| Vegetable/Nutrient | All | -Mg | -Zn | -B | -Fe |
| Tomato | 1.63 aA | 2.39 aA | 1.71 aA | 2.07 aA | 0.37 aB |
| Bell pepper | 1.83 aA | 0.95 bB | 1.75 aA | 1.65 aA | 0.60 aB |
| RDM (g) | |||||
| Vegetable/Nutrient | All | -Mg | -Zn | -B | -Fe |
| Tomato | 0.78 aA | 0.54 aB | 0.60 bB | 0.42 aC | 0.12 bD |
| Bell pepper | 0.69 aA | 0.50 aB | 0.76 aA | 0.55 aB | 0.32 aC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisboa, L.A.M.; Galindo, F.S.; Pagliari, P.H.; Goncalves, J.I.U.P.; Okazuka, M.H.; Cunha, M.L.O.; de Figueiredo, P.A.M. Morpho-Physiological Assessment of Tomato and Bell Pepper in Response to Nutrient Restriction. Stresses 2024, 4, 172-184. https://doi.org/10.3390/stresses4010010
Lisboa LAM, Galindo FS, Pagliari PH, Goncalves JIUP, Okazuka MH, Cunha MLO, de Figueiredo PAM. Morpho-Physiological Assessment of Tomato and Bell Pepper in Response to Nutrient Restriction. Stresses. 2024; 4(1):172-184. https://doi.org/10.3390/stresses4010010
Chicago/Turabian StyleLisboa, Lucas Aparecido Manzani, Fernando Shintate Galindo, Paulo Humberto Pagliari, João Igor Ussifati Pessoa Goncalves, Matheus Haruichi Okazuka, Matheus Luís Oliveira Cunha, and Paulo Alexandre Monteiro de Figueiredo. 2024. "Morpho-Physiological Assessment of Tomato and Bell Pepper in Response to Nutrient Restriction" Stresses 4, no. 1: 172-184. https://doi.org/10.3390/stresses4010010