
Binge-like Alcohol Administration Alters Decision Making in an Adolescent Rat Model: Role of N-Methyl-D-Aspartate Receptor Signaling



Abstract
:1. Introduction
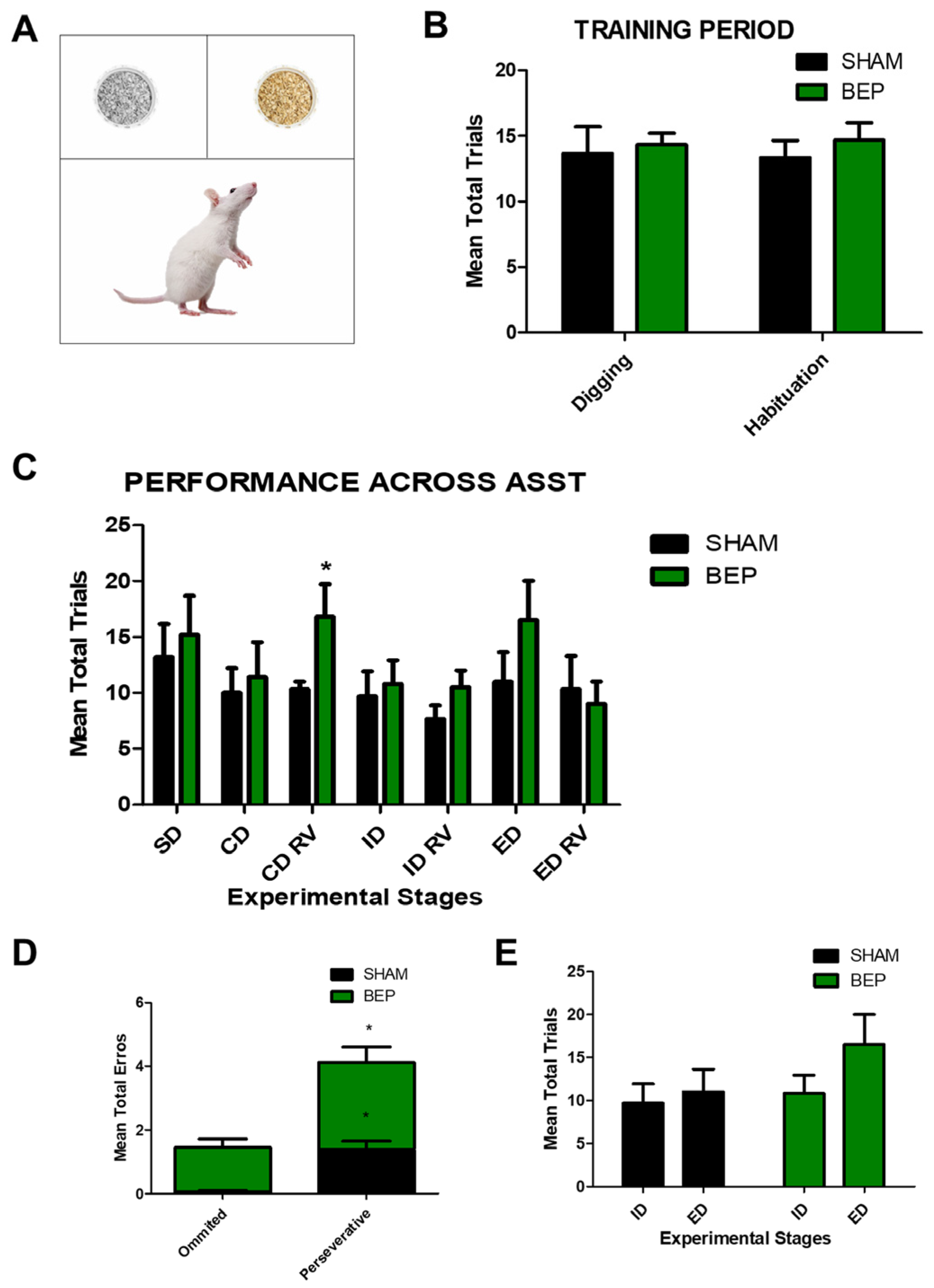
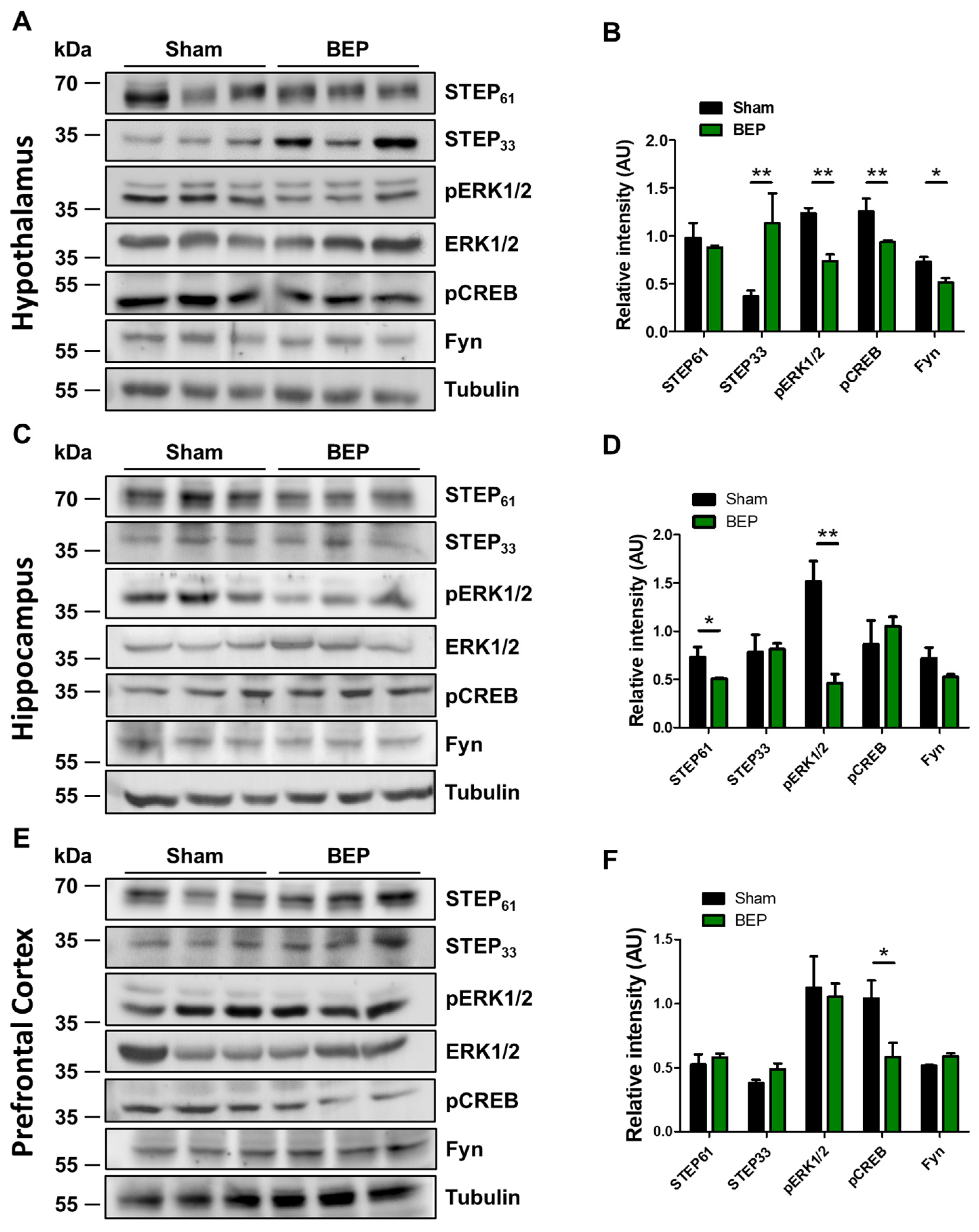
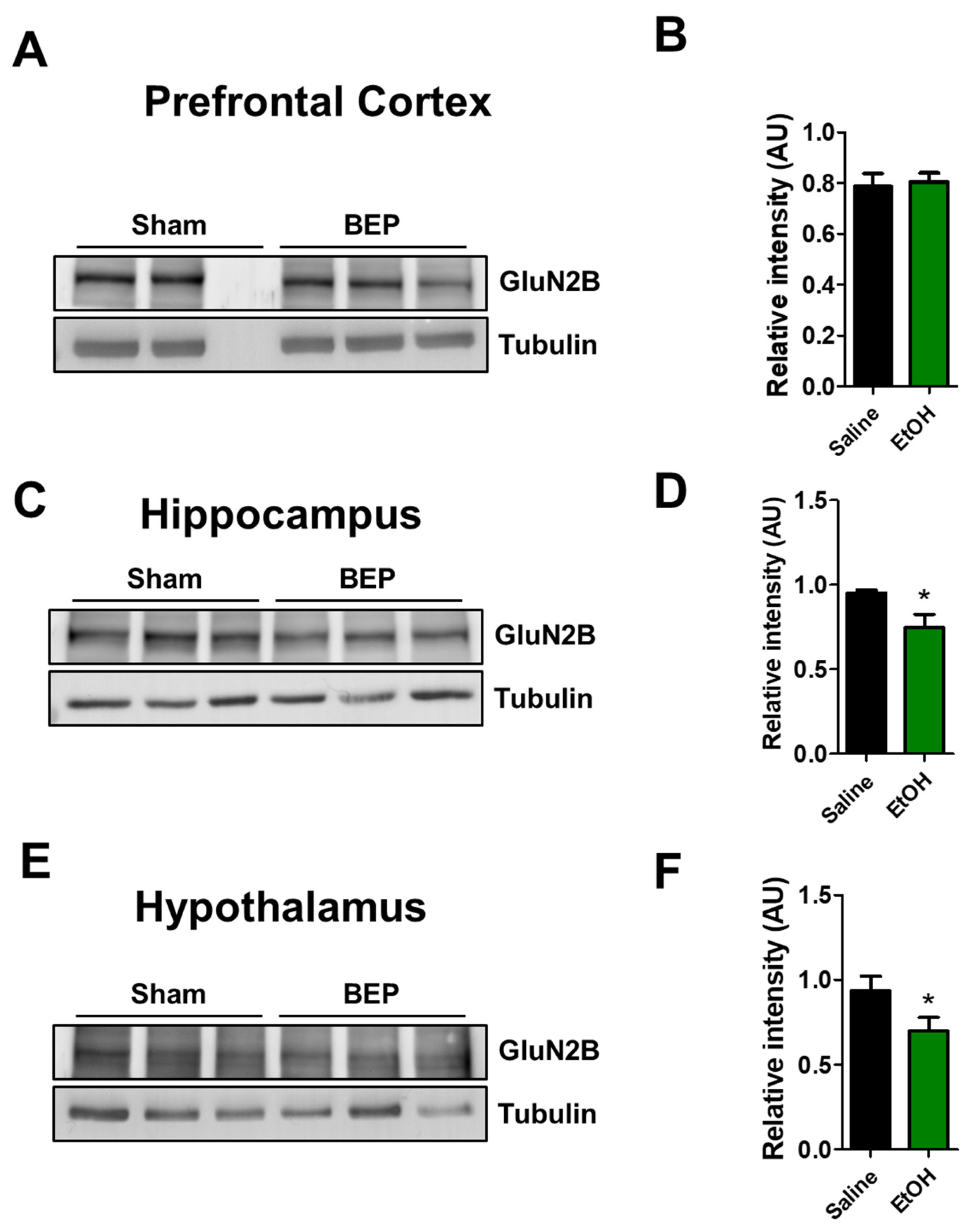
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents and Antibodies
4.3. Binge-like Ethanol Protocol in Rats
4.4. Attentional Set-Shifting Task (ASST)
4.5. Western Blot
4.6. PSD Isolation
4.7. Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koob, G.F.; Le Moal, M. Plasticity of reward neurocircuitry and the ‘dark side’ of drug addiction. Nat. Neurosci. 2005, 8, 1442–1444. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Status Report on Alcohol and Health 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Hendriks, H.F.J. Alcohol and Human Health: What Is the Evidence? Ann. Rev. Food Sci. Technol. 2020, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- SAMHSA. Substance Abuse and Mental Health Services Administration. National Survey on Drug Use and Health: National Findings; U.S. Department of Health and Human Services: Rockville, MA, USA, 2007.
- NIAAA. National Institute on Alcohol Abuse and Alcoholism. Strategic Plan 2017–2021; U.S. Department of Health and Human Services: Rockville, MA, USA, 2017.
- Chung, T.; Creswell, K.G.; Bachrach, R.; Clark, D.B.; Martin, C.S. Adolescent Binge Drinking. Alcohol Res. Curr. Rev. 2018, 39, 5–15. [Google Scholar]
- Merrill, J.E.; Carey, K.B. Drinking Over the Lifespan: Focus on College Ages. Alcohol Res. Curr. Rev. 2016, 38, 103–114. [Google Scholar]
- Briones, T.L.; Woods, J. Chronic binge-like alcohol consumption in adolescence causes depression-like symptoms possibly mediated by the effects of BDNF on neurogenesis. Neuroscience 2013, 254, 324–334. [Google Scholar] [CrossRef] [PubMed]
- McBride, O.; Cheng, H.G. Exploring the emergence of alcohol use disorder symptoms in the two years after onset of drinking: Findings from the National Surveys on Drug Use and Health. Addiction 2011, 106, 555–563. [Google Scholar] [CrossRef]
- Spear, L.P. Effects of adolescent alcohol consumption on the brain and behaviour. Nat. Rev. Neurosci. 2018, 19, 197–214. [Google Scholar] [CrossRef]
- Fuhrmann, D.; Knoll, L.J.; Blakemore, S.J. Adolescence as a Sensitive Period of Brain Development. Trends Cogn. Sci. 2015, 19, 558–566. [Google Scholar] [CrossRef]
- Dahl, R.E. Adolescent brain development: A period of vulnerabilities and opportunities. Keynote address. Ann. N. Y. Acad. Sci. 2004, 1021, 1–22. [Google Scholar] [CrossRef]
- Crone, E.A.; Dahl, R.E. Understanding adolescence as a period of social-affective engagement and goal flexibility. Nat. Rev. Neurosci. 2012, 13, 636–650. [Google Scholar] [CrossRef]
- Lauharatanahirun, N.; Maciejewski, D.F.; Kim-Spoon, J.; King-Casas, B. Risk-related brain activation is linked to longitudinal changes in adolescent health risk behaviors. Dev. Cogn. Neurosci. 2023, 63, 101291. [Google Scholar] [CrossRef] [PubMed]
- Squeglia, L.M.; Jacobus, J.; Tapert, S.F. The effect of alcohol use on human adolescent brain structures and systems. Handb. Clin. Neurol. 2014, 125, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Dees, W.L.; Hiney, J.K.; Srivastava, V.K. Alcohol and Puberty. Alcohol Res. Curr. Rev. 2017, 38, 277–282. [Google Scholar]
- Seemiller, L.R.; Gould, T.J. The effects of adolescent alcohol exposure on learning and related neurobiology in humans and rodents. Neurobiol. Learn. Mem. 2020, 172, 107234. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Dorsky, R.I. Development of the hypothalamus: Conservation, modification and innovation. Development 2017, 144, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Bear, M.H.; Bhimji, S.S. Neuroanatomy, Hypothalamus; StatPearls: Treasure Island, FL, USA, 2018. [Google Scholar]
- Flament-Durand, J. The hypothalamus: Anatomy and functions. Acta Psychiatr. Belg. 1980, 80, 364–375. [Google Scholar] [PubMed]
- Blaine, S.K.; Sinha, R. Alcohol, stress, and glucocorticoids: From risk to dependence and relapse in alcohol use disorders. Neuropharmacology 2017, 122, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.M.; Burgess, N. The hippocampus and memory: Insights from spatial processing. Nat. Rev. Neurosci. 2008, 9, 182–194. [Google Scholar] [CrossRef]
- Eichenbaum, H. A cortical-hippocampal system for declarative memory. Nat. Rev. Neurosci. 2000, 1, 41–50. [Google Scholar] [CrossRef]
- Eichenbaum, H. Hippocampus: Cognitive processes and neural representations that underlie declarative memory. Neuron 2004, 44, 109–120. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Carvajal, F.J.; Mira, R.G.; Arce, C.; Lerma-Cabrera, J.M.; Orellana, J.A.; Cerpa, W.; Quintanilla, R.A. Adolescent Binge Alcohol Exposure Affects the Brain Function Through Mitochondrial Impairment. Mol. Neurobiol. 2018, 55, 4473–4491. [Google Scholar] [CrossRef] [PubMed]
- Mira, R.G.; Lira, M.; Tapia-Rojas, C.; Rebolledo, D.L.; Quintanilla, R.A.; Cerpa, W. Effect of Alcohol on Hippocampal-Dependent Plasticity and Behavior: Role of Glutamatergic Synaptic Transmission. Front. Behav. Neurosci. 2019, 13, 288. [Google Scholar] [CrossRef]
- Vertes, R.P. Interactions among the medial prefrontal cortex, hippocampus and midline thalamus in emotional and cognitive processing in the rat. Neuroscience 2006, 142, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Klenowski, P.M. Emerging role for the medial prefrontal cortex in alcohol-seeking behaviors. Addict. Behav. 2018, 77, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Arnsten, A.F. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Itoga, C.A.; Chen, Y.; Fateri, C.; Echeverry, P.A.; Lai, J.M.; Delgado, J.; Badhon, S.; Short, A.; Baram, T.Z.; Xu, X. New viral-genetic mapping uncovers an enrichment of corticotropin-releasing hormone-expressing neuronal inputs to the nucleus accumbens from stress-related brain regions. J. Comp. Neurol. 2019, 527, 2474–2487. [Google Scholar] [CrossRef] [PubMed]
- Girotti, M.; Adler, S.M.; Bulin, S.E.; Fucich, E.A.; Paredes, D.; Morilak, D.A. Prefrontal cortex executive processes affected by stress in health and disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 85, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Logue, S.F.; Gould, T.J. The neural and genetic basis of executive function: Attention, cognitive flexibility, and response inhibition. Pharmacol. Biochem. Behav. 2014, 123, 45–54. [Google Scholar] [CrossRef]
- Orsini, C.A.; Heshmati, S.C.; Garman, T.S.; Wall, S.C.; Bizon, J.L.; Setlow, B. Contributions of medial prefrontal cortex to decision making involving risk of punishment. Neuropharmacology 2018, 139, 205–216. [Google Scholar] [CrossRef]
- Galandra, C.; Basso, G.; Cappa, S.; Canessa, N. The alcoholic brain: Neural bases of impaired reward-based decision-making in alcohol use disorders. Neurol. Sci. 2018, 39, 423–435. [Google Scholar] [CrossRef]
- Kumar, A. NMDA Receptor Function During Senescence: Implication on Cognitive Performance. Front. Neurosci. 2015, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, J.; Colechio, E.M.; Stanton, P.; Panksepp, J. Positive Emotional Learning Induces Resilience to Depression: A Role for NMDA Receptor-mediated Synaptic Plasticity. Curr. Neuropharmacol. 2017, 15, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Goebel-Goody, S.M.; Davies, K.D.; Alvestad Linger, R.M.; Freund, R.K.; Browning, M.D. Phospho-regulation of synaptic and extrasynaptic N-methyl-d-aspartate receptors in adult hippocampal slices. Neuroscience 2009, 158, 1446–1459. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.G.; Zukin, R.S. NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat. Rev. Neurosci. 2007, 8, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Paul, S.; Xu, Y.; Gurd, J.W.; Lombroso, P.J. Calcium-dependent cleavage of striatal enriched tyrosine phosphatase (STEP). J. Neurochem. 1999, 73, 1995–2001. [Google Scholar] [PubMed]
- Carvajal, F.J.; Mattison, H.A.; Cerpa, W. Role of NMDA Receptor-Mediated Glutamatergic Signaling in Chronic and Acute Neuropathologies. Neural Plast. 2016, 2016, 2701526. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, G.E.; Bading, H. Synaptic versus extrasynaptic NMDA receptor signalling: Implications for neurodegenerative disorders. Nat. Rev. Neurosci. 2010, 11, 682–696. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, Y.T. GluN2B-containing NMDARs in the mammalian brain: Pharmacology, physiology, and pathology. Front. Mol. Neurosci. 2023, 16, 1190324. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Cai, W.; Yu, J.; Liu, S.S.; Zhuo, M.; Li, B.M.; Zhang, X.H. Surface expression of hippocampal NMDA GluN2B receptors regulated by fear conditioning determines its contribution to memory consolidation in adult rats. Sci. Rep. 2016, 6, 30743. [Google Scholar] [CrossRef]
- Warburton, E.C.; Brown, M.W. Neural circuitry for rat recognition memory. Behav. Brain Res. 2015, 285, 131–139. [Google Scholar] [CrossRef]
- Clay, J.M.; Parker, M.O. The role of stress-reactivity, stress-recovery and risky decision-making in psychosocial stress-induced alcohol consumption in social drinkers. Psychopharmacology 2018, 235, 3243–3257. [Google Scholar] [CrossRef] [PubMed]
- Dias-Ferreira, E.; Sousa, J.C.; Melo, I.; Morgado, P.; Mesquita, A.R.; Cerqueira, J.J.; Costa, R.M.; Sousa, N. Chronic stress causes frontostriatal reorganization and affects decision-making. Science 2009, 325, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Popik, P.; Nikiforuk, A. Attentional Set-Shifting Paradigm in the Rat. Curr. Protoc. Neurosci. 2015, 72, 9.51.1–9.51.13. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, T.E.; Wang, Y.T. The intersections of NMDAR-dependent synaptic plasticity and cell survival. Neuropharmacology 2013, 74, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Lovinger, D.M.; White, G.; Weight, F.F. Ethanol inhibits NMDA-activated ion current in hippocampal neurons. Science 1989, 243, 1721–1724. [Google Scholar] [CrossRef] [PubMed]
- Lovinger, D.M.; White, G.; Weight, F.F. NMDA receptor-mediated synaptic excitation selectively inhibited by ethanol in hippocampal slice from adult rat. J. Neurosci. 1990, 10, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Mira, R.G.; Tapia-Rojas, C.; Perez, M.J.; Jara, C.; Vergara, E.H.; Quintanilla, R.A.; Cerpa, W. Alcohol impairs hippocampal function: From NMDA receptor synaptic transmission to mitochondrial function. Drug Alcohol Depend. 2019, 205, 107628. [Google Scholar] [CrossRef]
- Tait, D.S.; Bowman, E.M.; Neuwirth, L.S.; Brown, V.J. Assessment of intradimensional/extradimensional attentional set-shifting in rats. Neurosci. Biobehav. Rev. 2018, 89, 72–84. [Google Scholar] [CrossRef]
- Wang, R.; Reddy, P.H. Role of Glutamate and NMDA Receptors in Alzheimer’s Disease. J. Alzheimer’s Dis. JAD 2017, 57, 1041–1048. [Google Scholar] [CrossRef]
- Lu, S.M.; Yeh, H.H. Ethanol modulates AMPA-induced current responses of primary somatosensory cortical neurons. Neurochem. Int. 1999, 35, 175–183. [Google Scholar] [CrossRef]
- Roberto, M.; Varodayan, F.P. Synaptic targets: Chronic alcohol actions. Neuropharmacology 2017, 122, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Clapp, P.; Gibson, E.S.; Dell’acqua, M.L.; Hoffman, P.L. Phosphorylation regulates removal of synaptic N-methyl-D-aspartate receptors after withdrawal from chronic ethanol exposure. J. Pharmacol. Exp. Ther. 2010, 332, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Euston, D.R.; Gruber, A.J.; McNaughton, B.L. The role of medial prefrontal cortex in memory and decision making. Neuron 2012, 76, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T. Collateral projection from the amygdalo--hippocampal transition area and CA1 to the hypothalamus and medial prefrontal cortex in the rat. Neurosci. Res. 2000, 38, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.; Hinman, J.R.; Jacobson, T.K.; Szkudlarek, E.; Argraves, M.; Escabi, M.A.; Markus, E.J. Dissociation between dorsal and ventral hippocampal theta oscillations during decision-making. J. Neurosci. 2013, 33, 6212–6224. [Google Scholar] [CrossRef] [PubMed]
- Le Merre, P.; Esmaeili, V.; Charriere, E.; Galan, K.; Salin, P.A.; Petersen, C.C.H.; Crochet, S. Reward-Based Learning Drives Rapid Sensory Signals in Medial Prefrontal Cortex and Dorsal Hippocampus Necessary for Goal-Directed Behavior. Neuron 2018, 97, 83–91.E5. [Google Scholar] [CrossRef] [PubMed]
- Strange, B.A.; Witter, M.P.; Lein, E.S.; Moser, E.I. Functional organization of the hippocampal longitudinal axis. Nat. Rev. Neurosci. 2014, 15, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Kjelstrup, K.B.; Solstad, T.; Brun, V.H.; Hafting, T.; Leutgeb, S.; Witter, M.P.; Moser, E.I.; Moser, M.B. Finite scale of spatial representation in the hippocampus. Science 2008, 321, 140–143. [Google Scholar] [CrossRef]
- Moser, M.B.; Moser, E.I.; Forrest, E.; Andersen, P.; Morris, R.G. Spatial learning with a minislab in the dorsal hippocampus. Proc. Natl. Acad. Sci. USA 1995, 92, 9697–9701. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 1999, 22, 105–122. [Google Scholar] [CrossRef]
- Cook, S.C.; Wellman, C.L. Chronic stress alters dendritic morphology in rat medial prefrontal cortex. J. Neurobiol. 2004, 60, 236–248. [Google Scholar] [CrossRef]
- Liston, C.; McEwen, B.S.; Casey, B.J. Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proc. Natl. Acad. Sci. USA 2009, 106, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Townshend, J.M.; Kambouropoulos, N.; Griffin, A.; Hunt, F.J.; Milani, R.M. Binge drinking, reflection impulsivity, and unplanned sexual behavior: Impaired decision-making in young social drinkers. Alcohol. Clin. Exp. Res. 2014, 38, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Bechara, A.; Gong, Q.; Huang, X.; Li, X.; Xue, G.; Wong, S.; Lu, Z.L.; Palmer, P.; Wei, Y.; et al. Abnormal affective decision making revealed in adolescent binge drinkers using a functional magnetic resonance imaging study. Psychol. Addict. Behav. J. Soc. Psychol. Addict. Behav. 2013, 27, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Salling, M.C.; Skelly, M.J.; Avegno, E.; Regan, S.; Zeric, T.; Nichols, E.; Harrison, N.L. Alcohol Consumption during Adolescence in a Mouse Model of Binge Drinking Alters the Intrinsic Excitability and Function of the Prefrontal Cortex through a Reduction in the Hyperpolarization-Activated Cation Current. J. Neurosci. 2018, 38, 6207–6222. [Google Scholar] [CrossRef]
- Ketchesin, K.D.; Stinnett, G.S.; Seasholtz, A.F. Binge Drinking Decreases Corticotropin-Releasing Factor-Binding Protein Expression in the Medial Prefrontal Cortex of Mice. Alcohol. Clin. Exp. Res. 2016, 40, 1641–1650. [Google Scholar] [CrossRef]
- Robinson, S.L.; Marrero, I.M.; Perez-Heydrich, C.A.; Sepulveda-Orengo, M.T.; Reissner, K.J.; Thiele, T.E. Medial prefrontal cortex neuropeptide Y modulates binge-like ethanol consumption in C57BL/6J mice. Neuropsychopharmacology 2019, 44, 1132–1140. [Google Scholar] [CrossRef]
- McClintick, J.N.; McBride, W.J.; Bell, R.L.; Ding, Z.M.; Liu, Y.; Xuei, X.; Edenberg, H.J. Gene expression changes in the ventral hippocampus and medial prefrontal cortex of adolescent alcohol-preferring (P) rats following binge-like alcohol drinking. Alcohol 2018, 68, 37–47. [Google Scholar] [CrossRef]
- Tapia-Rojas, C.; Torres, A.K.; Quintanilla, R.A. Adolescence Binge Alcohol Consumption Induces Hippocampal Mitochondrial Impairment that Persists during the Adulthood. Neuroscience 2019, 406, 356–368. [Google Scholar] [CrossRef]
- Mira, R.G.; Lira, M.; Quintanilla, R.A.; Cerpa, W. Alcohol consumption during adolescence alters the hippocampal response to traumatic brain injury. Biochem. Biophys. Res. Commun. 2020, 528, 514–519. [Google Scholar] [CrossRef]
- Linley, S.B.; Gallo, M.M.; Vertes, R.P. Lesions of the ventral midline thalamus produce deficits in reversal learning and attention on an odor texture set shifting task. Brain Res. 2016, 1649, 110–122. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| STAGE | STIMULI | COMBINATION | ||
|---|---|---|---|---|
| TRAINING | ||||
| Digging training | Corn cob bedding | |||
| TASK | ODOR | MEDIUM | (+) | (−) |
| SD | M1 corn cob bedding | M2 | M1 | |
| M2 paper bedding | ||||
| CD | O1 ginger | M1 corn cob bedding | O1/M1 | O2/M2 |
| O2 cinnamon | M2 paper bedding | O2/M1 | O1/M2 | |
| CD rev | O1 ginger | M1 corn cob bedding | O2/M1 | O1/M2 |
| O2 cinnamon | M2 paper bedding | O2/M2 | O1/M1 | |
| ID | O3 lemon | M3 cellulose bedding | O3/M3 | O4/M4 |
| O4 fennel | M4 rubber bedding | O4/M3 | O3/M4 | |
| ID rev | O3 lemon | M3 cellulose bedding | O3/M4 | O4/M3 |
| O4 fennel | M4 rubber bedding | O4/M4 | O3/M3 | |
| ED | O5 citronella | M5 gravel bedding | M5/O5 | M6/O6 |
| O6 clove | M6 plush bedding | M6/O5 | M5/O6 | |
| ED rev | O5 citronella | M5 gravel bedding | M5/O6 | M6/O5 |
| O6 clove | M6 plush bedding | M6/O6 | M5/O5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arce, C.; Mira, R.G.; Lira, M.; Cerpa, W. Binge-like Alcohol Administration Alters Decision Making in an Adolescent Rat Model: Role of N-Methyl-D-Aspartate Receptor Signaling. Stresses 2024, 4, 1-13. https://doi.org/10.3390/stresses4010001
Arce C, Mira RG, Lira M, Cerpa W. Binge-like Alcohol Administration Alters Decision Making in an Adolescent Rat Model: Role of N-Methyl-D-Aspartate Receptor Signaling. Stresses. 2024; 4(1):1-13. https://doi.org/10.3390/stresses4010001
Chicago/Turabian StyleArce, Camila, Rodrigo G. Mira, Matías Lira, and Waldo Cerpa. 2024. "Binge-like Alcohol Administration Alters Decision Making in an Adolescent Rat Model: Role of N-Methyl-D-Aspartate Receptor Signaling" Stresses 4, no. 1: 1-13. https://doi.org/10.3390/stresses4010001