
Nitric Oxide- and Sulfur-Mediated Reversal of Cadmium-Inhibited Photosynthetic Performance Involves Hydrogen Sulfide and Regulation of Nitrogen, Sulfur, and Antioxidant Metabolism in Mustard



Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Growth Conditions
2.2. Plant Growth and Relative Water Content (RWC)
2.3. Measurement of Photosynthesis and Related Variables
2.4. Assessment of Oxidative Damage
2.4.1. In Vitro Estimation of H2O2 and O2•−
2.4.2. In Vivo Localization of H2O2 and O2•− and Lipid Peroxidation
2.4.3. In Vitro Measurement of Leaf Lipid Peroxidation
2.5. Assay of Antioxidant Enzymes
2.5.1. SOD (EC 1.15.1.1)
2.5.2. GR (EC 1.6.4.2)
2.5.3. APX (EC 1.11.1.11)
2.6. Assay of Non-Protein Thiol (NPT) and Phytochelatin (PC) Content
2.7. Estimation of Leaf N Content, Nitrate Reductase (NR) Activity, Photosynthetic Sulfur Use Efficiency (p-SUE), and Photosynthetic Nitrogen Use Efficiency (-NUE)
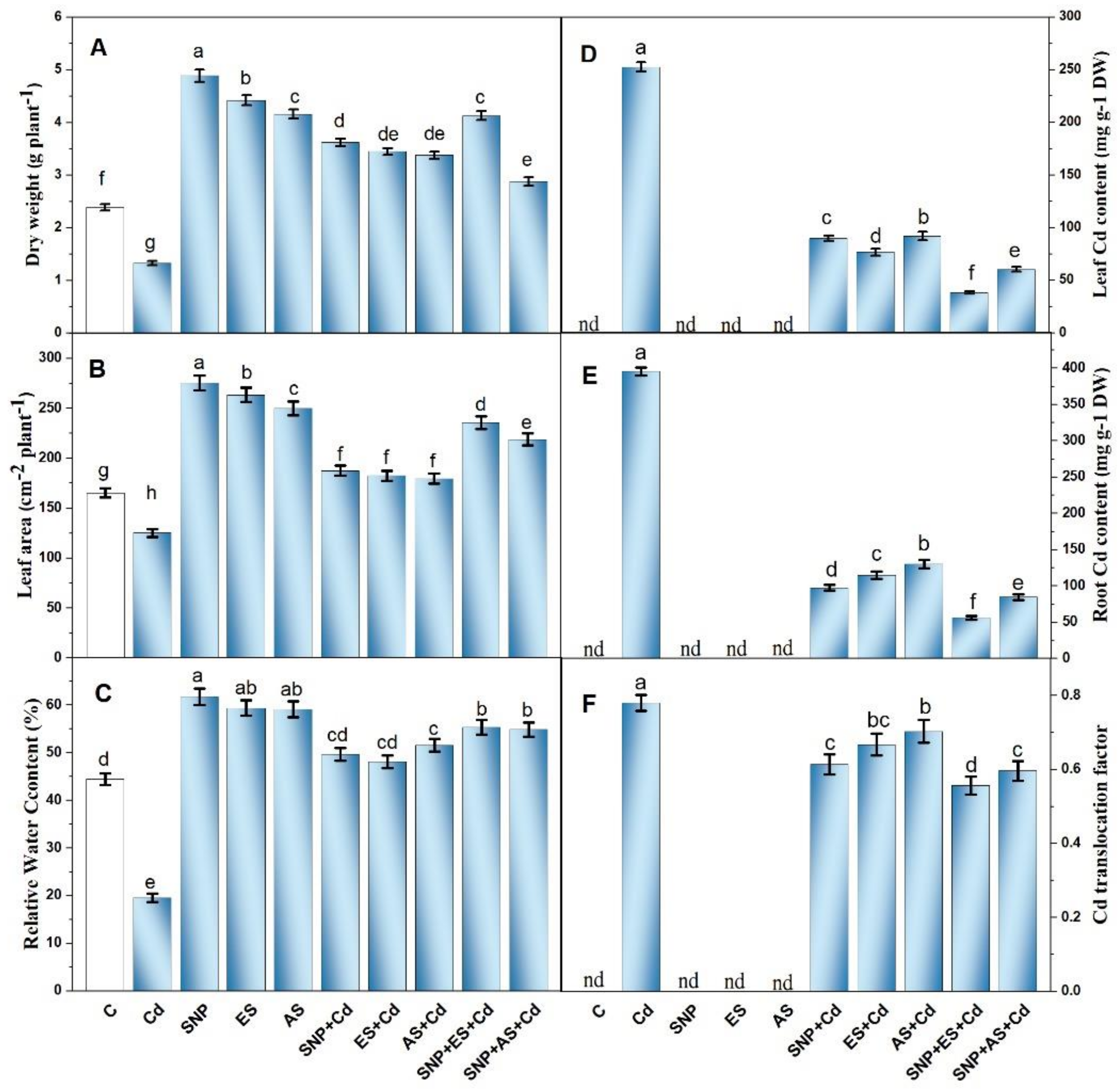
2.8. Quantification and Histochemical Localization of Cd
2.9. Assay for S-Assimilating Enzymes, S, Cys, GSH, and AsA Content
2.10. Determination of Endogenous NO and H2S Content
2.11. Determination of L-DES and OAS-TL Activity
2.12. Statistical Analysis
3. Results
3.1. Effect of SNP and/or S on Plant Growth, Water Status, and Cd Accumulation
3.2. Effect of SNP and/or S on Photosynthetic Traits under Cd Stress
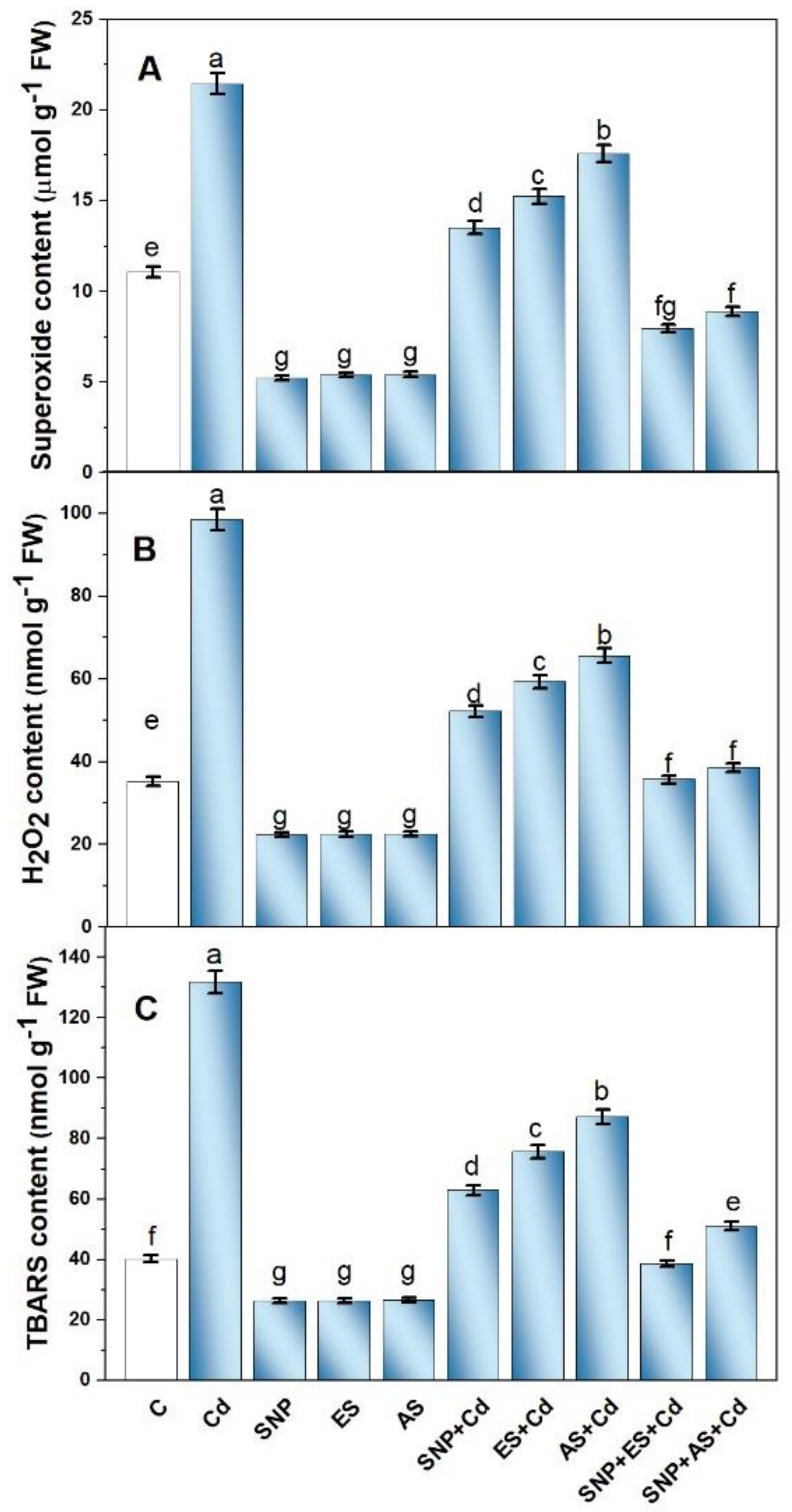
3.3. Effect of SNP and/or S on Cd-Induced Oxidative Stress
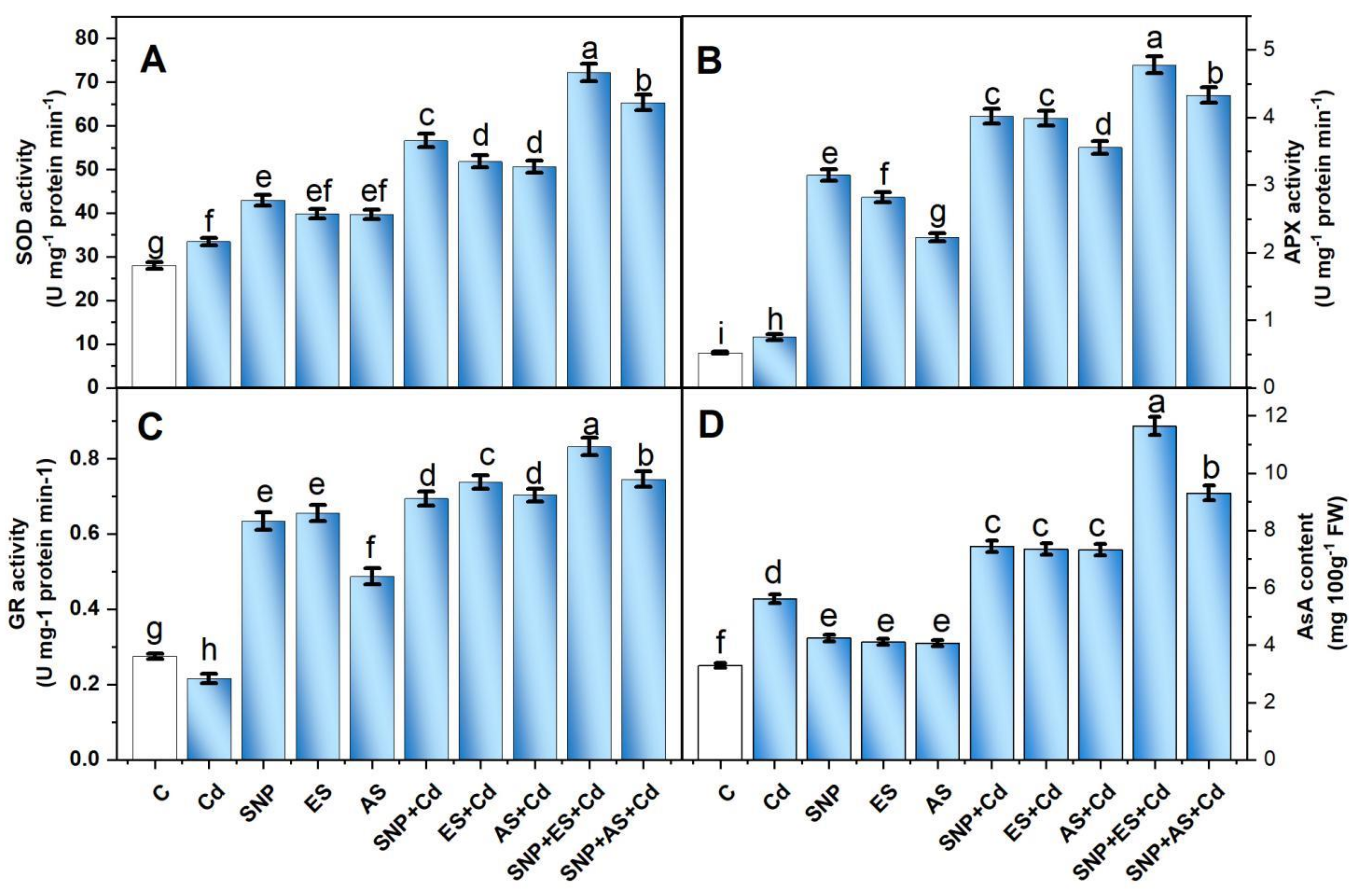
3.4. Effect of SNP and/or S on Antioxidants under Cd Stress
3.5. Effect of SNP and/or S on S-Assimilation under Cd Stress
3.6. Effect of SNP and/or S on N Assimilation, p-SUE, p-NUE, and NPT and PC Content
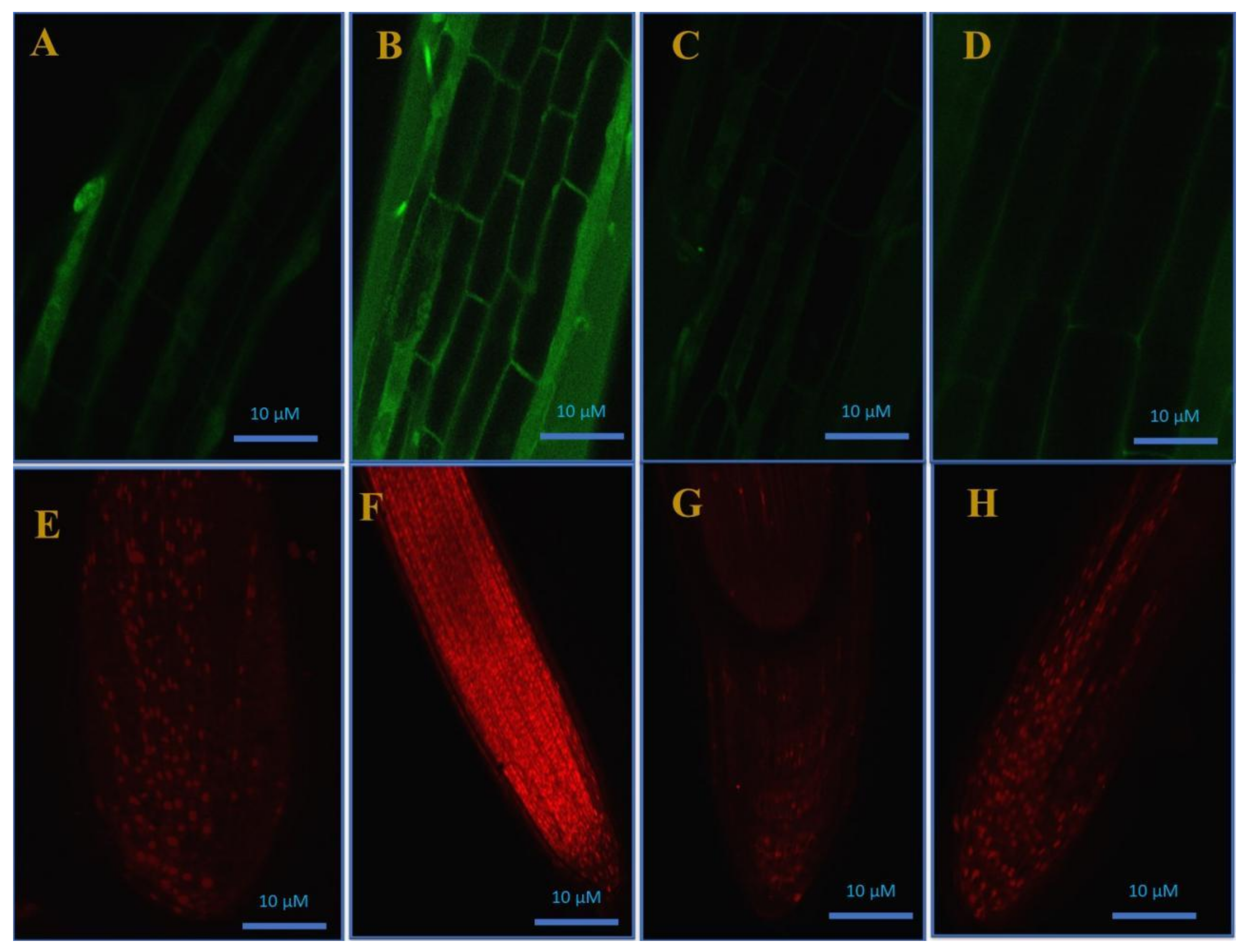
3.7. Confocal Laser Microscopy
3.8. How SNP and/or S Influence Endogenous NO and H2S Production
3.9. Validation of Role of H2S in SNP plus S-Mediated Response by Inhibiting NO and H2S under Cd Stress
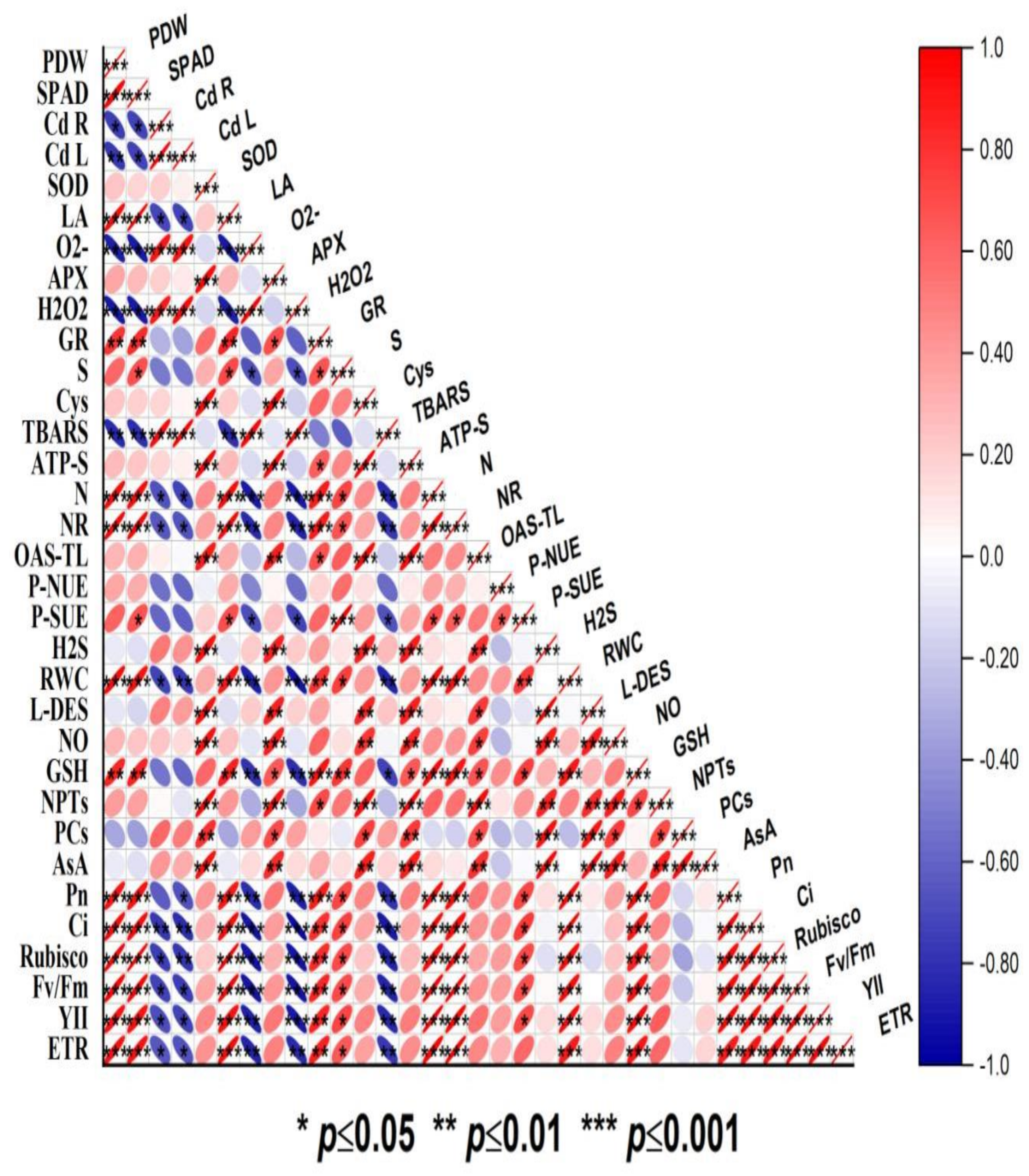
3.10. Relationship
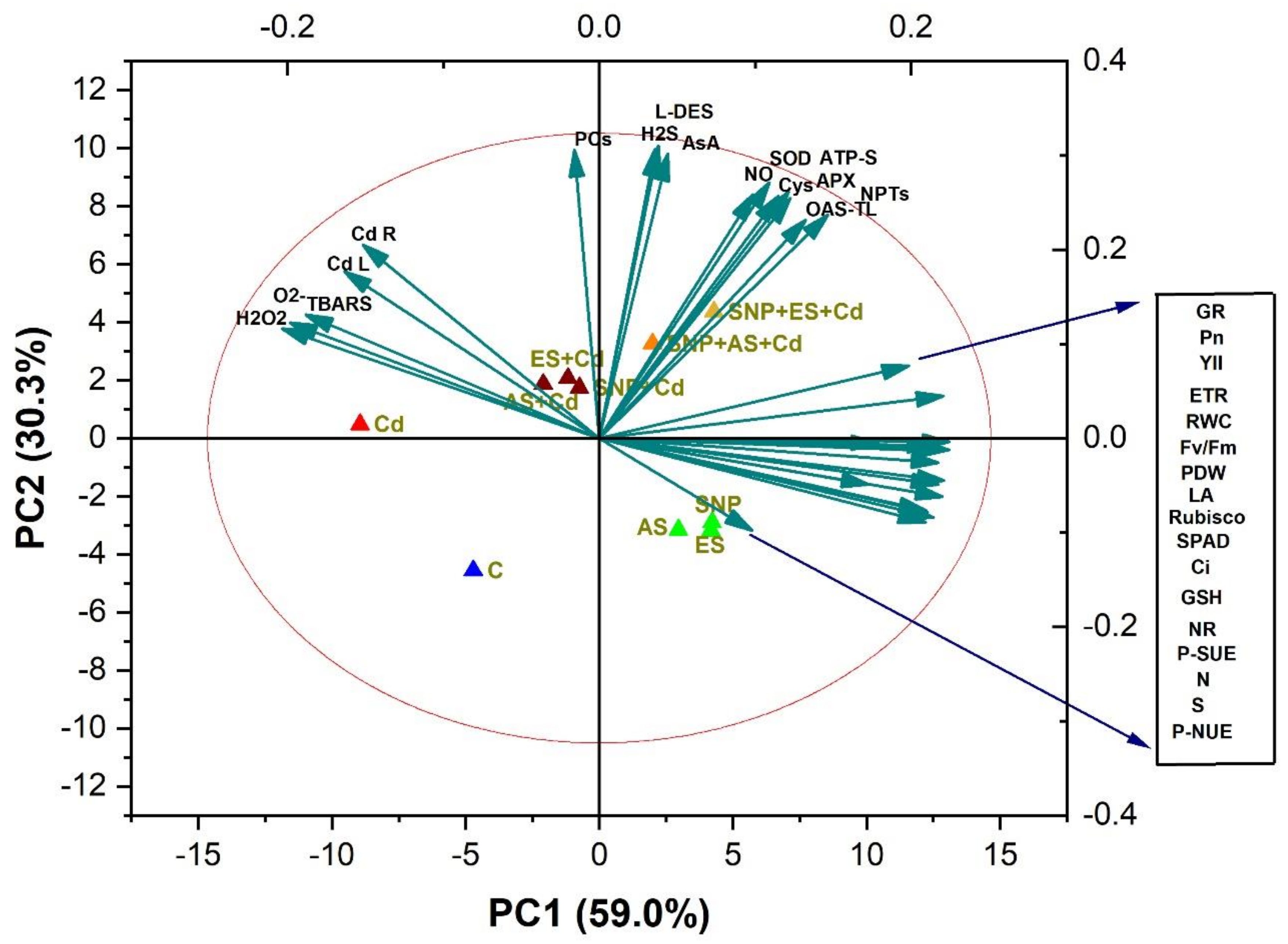
3.11. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubey, S.; Shri, M.; Gupta, A.; Rani, V.; Chakrabarty, D. Toxicity and detoxification of heavy metals during plant growth and metabolism. Environ. Chem. Lett. 2018, 16, 1169–1192. [Google Scholar] [CrossRef]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2020, 52, 675–726. [Google Scholar] [CrossRef]
- Shiyu, Q.I.N.; Hongen, L.I.U.; Zhaojun, N.I.E.; Rengel, Z.; Wei, G.A.O.; Chang, L.I.; Peng, Z.H.A.O. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere 2020, 30, 168–180. [Google Scholar]
- Mir, I.R.; Gautam, H.; Anjum, N.A.; Masood, A.; Khan, N.A. Calcium and nitric oxide signaling in plant cadmium stress tolerance: A cross talk. S. Afr. J. Bot. 2022, 150, 387–403. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Sehar, Z.; Anjum, N.A.; Masood, A.; Khan, N.A. The outcomes of the functional interplay of nitric oxide and hydrogen sulfide in metal stress tolerance in plants. Plant Physiol. Biochem. 2020, 155, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, S.M.; Vats, S.; Bhat, J.A.; Dhakte, P.; Goyal, V.; Khatri, P.; Deshmukh, R. Nitric oxide and hydrogen sulfide crosstalk during heavy metal stress in plants. Physiol. Plant. 2020, 168, 437–455. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Masood, A.; Anjum, N.A.; Khan, N.A. Ethylene-nitrogen synergism induces tolerance to copper stress by modulating antioxidant system and nitrogen metabolism and improves photosynthetic capacity in mustard. Environ. Sci. Pollut. Res. 2022, 25, 49029–49049. [Google Scholar] [CrossRef]
- Rasheed, F.; Mir, I.R.; Sehar, Z.; Fatma, M.; Gautam, H.; Khan, S.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation. Plants 2022, 11, 3131. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, Q.F.; Li, J.; Xiong, J.; Zhou, L.N.; He, S.L.; Liu, H. Effects of exogenous sulfur on alleviating cadmium stress in tartary buckwheat. Sci. Rep. 2019, 9, 7397. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Freschi, L.; Corpas, F.J.; Khan, N.A. Nitric oxide signaling and its crosstalk with other plant growth regulators in plant responses to abiotic stress. Environ. Sci. Pollut. Res. 2017, 24, 2273–2285. [Google Scholar] [CrossRef]
- Corpas, F.J.; González-Gordo, S.; Cañas, A.; Palma, J.M. Nitric oxide and hydrogen sulfide in plants: Which comes first? J.Exp. Bot. 2019, 70, 4391–4404. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.; Ali, H.M.; Khan, M.I.R.; Singh, V.P.; Siddiqui, M.H. Exogenous nitric oxide requires endogenous hydrogen sulfide to induce the resilience through sulfur assimilation in tomato seedlings under hexavalent chromium toxicity. Plant Physiol. Biochem. 2020, 155, 20–34. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Masood, A.; Anjum, N.A.; Khan, N.A. Nitric oxide pre-treatment advances seed germination and alleviates copper-induced photosynthetic inhibition in Indian mustard. Plants 2020, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ruiz, M.; Aparicio-Chacón, M.V.; Palma, J.M.; Corpas, F.J. Arsenate disrupts ion balance, sulfur and nitric oxide metabolisms in roots and leaves of pea (Pisum sativum L.) plants. Environ. Exp. Bot. 2019, 161, 143–156. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Nitro-oxidative stress vs oxidative or nitrosative stress in higher plants. New Phytol. 2013, 199, 633–635. [Google Scholar] [CrossRef]
- Begara-Morales, J.C.; Sánchez-Calvo, B.; Chaki, M.; Valderrama, R.; Mata-Pérez, C.; Padilla, M.N.; Barroso, J.B. Antioxidant systems are regulated by nitric oxide-mediated post-translational modifications (NO-PTMs). Front. Plant Sci. 2016, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Mobin, M.; Anjum, N.A.; Khan, N.A. Modulation and significance of nitrogen and sulfur metabolism in cadmium challenged plants. Plant Growth Regul. 2016, 78, 1–11. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Gautam, H.; Majid, A.; Anjum, N.A.; Masood, A.; Khan, N.A. Appraisal of functional significance of sulfur assimilatory products in plants under elevated metal accumulation. Crop Pasture Sci. 2022, 73, 573–584. [Google Scholar] [CrossRef]
- Rego, T.J.; Sahrawat, K.L.; Wani, S.P.; Pardhasaradhi, G. Widespread deficiencies of sulfur, boron, and zinc in Indian semi-arid tropical soils: On-farm crop responses. J. Plant Nutr. 2007, 30, 1569–1583. [Google Scholar] [CrossRef] [Green Version]
- Bashir, H.; Ahmad, J.; Bagheri, R.; Nauman, M.; Qureshi, M.I. Limited sulfur resource forces Arabidopsis thaliana to shift towards non-sulfur tolerance under cadmium stress. Environ. Exp. Bot. 2013, 94, 19–32. [Google Scholar] [CrossRef]
- Mir, I.R.; Rather, B.A.; Masood, A.; Majid, A.; Sehar, Z.; Anjum, N.A.; Sofo, A.; D’Ippolito, I.; Khan, N.A. Soil sulfur sources differentially enhance cadmium tolerance in Indian mustard (Brassica juncea L.). Soil Syst. 2021, 5, 29. [Google Scholar] [CrossRef]
- Bortoloti, G.A.; Baron, D. Phytoremediation of toxic heavy metals by brassica plants: A biochemical and physiological approach. Environ. Adv. 2022, 8, 100204. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gill, S.S.; Umar, S.; Ahmad, I.; Duarte, A.C.; Pereira, E. Improving growth and productivity of oleiferous Brassicas under changing environment: Significance of nitrogen and sulphur nutrition, and underlying mechanisms. Sci. World J. 2012, 2012, 657808. [Google Scholar] [CrossRef] [Green Version]
- Mir, I.R.; Rather, B.A.; Masood, A.; Anjum, N.A.; Khan, N.A. Nitrogen sources mitigate cadmium phytotoxicity differentially by modulating cellular buffers, N-assimilation, non-protein thiols, and phytochelatins in mustard (Brassica juncea L.). J. Soil Sci. Plant Nutr. 2022, 18, 1–21. [Google Scholar] [CrossRef]
- Corpas, F.J. Hydrogen sulfide: A new warrior against abiotic stress. Trends Plant Sci. 2019, 24, 983–988. [Google Scholar] [CrossRef]
- Kushwaha, B.K.; Singh, V.P. Mitigation of chromium (VI) toxicity by additional sulfur in some vegetable crops involves glutathione and hydrogen sulfide. Plant Physiol. Biochem. 2020, 155, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Gautam, H.; Fatma, M.; Sehar, Z.; Mir, I.R.; Khan, N.A. Hydrogen sulfide, ethylene, and nitric oxide regulate redox homeostasis and protect photosynthetic metabolism under high temperature stress in rice plants. Antioxidants 2022, 11, 1478. [Google Scholar] [CrossRef]
- Bano, K.; Kumar, B.; Alyemeni, M.N.; Ahmad, P. Protective mechanisms of sulfur against arsenic phytotoxicity in Brassica napus by regulating thiol biosynthesis, sulfur-assimilation, photosynthesis, and antioxidant response. Plant Physiol. Biochem. 2022, 188, 1–11. [Google Scholar] [CrossRef]
- Alsahli, A.A.; Bhat, J.A.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Hydrogen sulfide (H2S) mitigates arsenic (As)-induced toxicity in pea (Pisum sativum L.) plants by regulating osmoregulation, antioxidant defense system, ascorbate glutathione cycle and glyoxalase system. J. Plant Growth Regul. 2020, 40, 2515–2531. [Google Scholar] [CrossRef]
- Fatma, M.; Masood, A.; Per, T.S.; Rasheed, F.; Khan, N.A. Interplay between nitric oxide and sulfur assimilation in salt tolerance in plants. Crop J. 2016, 4, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric oxide alleviates salt stress inhibited photosynthetic performance by interacting with sulfur assimilation in mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Prasad, S.M.; Singh, V.P. Additional calcium and sulfur manages hexavalent chromium toxicity in Solanum lycopersicum L. and Solanum melongena L. seedlings by involving nitric oxide. J. Hazard. Mat. 2020, 39, 122607. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; AlZuaibr, F.M.; Al-Huqail, A.A.; Siddiqui, M.H.; Ali, H.M.; Al-Muwayhi, M.A.; Al-Haque, H.N. Hydrogen sulfide-mediated activation of O-Acetylserine (thiol) Lyase and L/D-Cysteine desulfhydrase enhance dehydration tolerance in Eruca sativa mill. Int. J. Mol. Sci. 2018, 19, 3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usuda, H. The activation state of ribulose 1,5-bisphosphate carboxylase in maize leaves in dark and light. Plant Cell Physiol. 1985, 26, 1455–1463. [Google Scholar]
- Okuda, T.; Masuda, Y.; Yamanka, A.; Sagisaka, S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991, 97, 1265–1267. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.L.; Cui, J.; Tao, L.; Yang, H. Fluroxypyr triggers oxidative damage by producing superoxide and hydrogen peroxide in rice (Oryza sativa). Ecotoxicology 2010, 19, 124–132. [Google Scholar] [CrossRef]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B. Histochemical detection of superoxide 780 and H2O2 accumulation in Brassica juncea seedlings. Biol. Protoc. 2014, 4, e1108. [Google Scholar]
- Awasthi, J.P.; Saha, B.; Chowardhara, B.; Devi, S.S.; Borgohain, P.; Panda, S.K. Qualitative analysis of lipid peroxidation in plants under multiple stress through schiff’s reagent: A histochemical approach. Bio-protocol 2018, 8, e2807. [Google Scholar] [CrossRef]
- Dhindsa, R.H.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence correlated with increased level of membrane permeability, lipid peroxidation and decreased level of SOD and CAT. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- De Vos, C.R.; Vonk, M.J.; Vooijs, R.; Schat, H. Glutathione depletion due to copper-induced phytochelatin synthesis causes oxidative stress in Silene cucubalus. Plant Physiol. 1992, 98, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Lindner, R.C. Rapid analytical method for some of the more common organic substances of plant and soil. Plant Physiol. 1944, 19, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.M.; Warner, R.L.; Kleinhofs, A. In vitro stability of nitrate reductase from barley leaves. Phytochemistry 1982, 21, 531–533. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Histochemical methods for detection of heavy metals and strontium in the tissues of higher plants. Russian J. Plant Physiol. 2011, 58, 721–727. [Google Scholar] [CrossRef]
- Lappartient, A.G.; Touraine, B. Demand-driven control of root ATP sulfurylase activity and SO42-uptake in intact canola (the role of phloem-translocated glutathione). Plant Physiol. 1996, 111, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Chesnin, L.; Yien, C.H. 1950. Turbidimetric determination of available sulphates. Soil Sci. Soc. Am. Proc. 1950, 15, 149–151. [Google Scholar] [CrossRef]
- Gaitonde, M.K. A spectrophotometric method for the direct determination of cysteine in the presence of other naturally occurring amino acids. Biochem. J. 1967, 104, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Guo, Z.; Xing, J.; Huang, B. Nitric oxide is involved in abscisic acid-induced antioxidant activities in Stylosanthes guianensis. Environ. Exp. Bot. 2005, 56, 3223–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nashef, A.S.; Osuga, D.T.; Feeney, R.E. Determination of hydrogen sulfide with 5,5′-dithiobis-(2-nitrobenzoic acid), N-ethylmaleimide, and parachloromercuribenzoate. Anal. Biochem. 1977, 79, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Bloem, E.; Riemenschneider, A.; Volker, J.; Papenbrock, J.; Schmidt, A.; Salac, I.; Schnug, E. Sulphur supply and infection with Pyrenopeziza brassicae influence L-cysteine desulphydrase activity in Brassica napus L. J. Exp. Bot. 2004, 55, 2305–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Sehar, Z.; Khan, N.A. Gibberellic acid and sulfur-mediated reversal of cadmium-inhibited photosynthetic performance in mungbean (Vigna radiata L.) involves nitric oxide. J. Plant Growth Regul. 2020, 39, 1605–1615. [Google Scholar] [CrossRef]
- Per, T.S.; Masood, A.; Khan, N.A. Nitric oxide improves S-assimilation and GSH production to prevent inhibitory effects of cadmium stress on photosynthesis in mustard (Brassica juncea L.). Nitric Oxide 2017, 68, 111–124. [Google Scholar] [CrossRef]
- Fuentes-Lara, L.O.; Medrano-Macías, J.; Pérez-Labrada, F.; Rivas-Martínez, E.N.; García-Enciso, E.L.; González-Morales, S.; Benavides-Mendoza, A. From elemental sulfur to hydrogen sulfide in agricultural soils and plants. Molecules 2019, 24, 2282. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Song, K.; Shi, L.; Duan, D.; Zhang, H.; Sun, Y.; Qin, Q.; Xue, Y. Influence of elemental sulfur on cadmium bioavailability, microbial community in paddy soil and Cd accumulation in rice plants. Sci. Rep. 2021, 11, 11468. [Google Scholar] [CrossRef]
- Dong, Y.; Silbermann, M.; Speiser, A.; Forieri, I.; Linster, E.; Poschet, G.; Wirtz, M. Sulfur availability regulates plant growth via glucose-TOR signaling. Nat. Commun. 2017, 8, 1174. [Google Scholar] [CrossRef] [Green Version]
- Lehotai, N.; Feigl, G.; Koós, Á.; Molnár, Á.; Ördög, A.; Pető, A.; Erdei, L.; Kolbert, Z. Nitric oxide–cytokinin interplay influences selenite sensitivity in Arabidopsis. Plant Cell Rep. 2016, 35, 2181–2195. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Zeliha, K.; Erdal, I. Effects of elemental sulfur and sulfur containing waste on nutrient concentrations and growth of bean and corn plants grown on a calcareous soil. Afr. J. Biotechnol. 2009, 8, 4481–4489. [Google Scholar]
- Liu, F.; Guo, F.Q. Nitric oxide deficiency accelerates chlorophyll breakdown and stability loss of thylakoid membranes during dark-induced leaf senescence in Arabidopsis. PLoS ONE 2013, 8, e56345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, I.; Lindermayr, C. Nitric oxide-based protein modification: Formation and site-specificity of protein S-nitrosylation. Front. Plant Sci. 2013, 4, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.N.; Siddiqui, M.H.; AlSolami, M.A.; Alamri, S.; Hu, Y.; Ali, H.M.; Al-Ghamdi, A. Crosstalk of hydrogen sulfide and nitric oxide requires calcium to mitigate impaired photosynthesis under cadmium stress by activating defense mechanisms in Vigna radiata. Plant Physiol. Biochem. 2020, 156, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Martí, M.C.; Jiménez, A.; Sevilla, F. Thioredoxin network in plant mitochondria: Cysteine S-posttranslational modifications and stress conditions. Front. Plant Sci. 2020, 11, 1476. [Google Scholar] [CrossRef] [PubMed]
- Wangeline, A.L.; Burkhead, J.L.; Hale, K.L.; Lindblom, S.D.; Terry, N.; Pilon, M.; Pilon-Smits, E.A. Overexpression of ATP sulfurylase in Indian mustard—Effects on tolerance and accumulation of twelve metals. J. Environ. Qual. 2004, 33, 54–60. [Google Scholar]
- Veliz, C.G.; Roberts, I.N.; Criado, M.V.; Caputo, C. Sulphur deficiency inhibits nitrogen assimilation and recycling in barley plants. Biol. Plant. 2017, 61, 675–684. [Google Scholar] [CrossRef]
- Liang, Y.; Zheng, P.; Li, S.; Li, K.Z.; Xu, H.N. Nitrate reductase-dependent NO production is involved in H2S-induced nitrate stress tolerance in tomato via activation of antioxidant enzymes. Sci. Hort. 2018, 229, 207–214. [Google Scholar] [CrossRef]
- Lea, U.S.; Leydecker, M.T.; Quillere’, I.; Meyer, C.; Lillo, C. Posttranslational regulation of nitrate reductase strongly affects the levels of free amino acids and nitrate, whereas transcriptional regulation has only minor influence. Plant Physiol. 2006, 140, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol. Plant. 2020, 168, 345–360. [Google Scholar] [CrossRef]
- Wang, B.L.; Shi, L.; Li, Y.X.; Zhang, W.H. Boron toxicity is alleviated by hydrogen sulfide in cucumber (Cucumis sativus L.) seedlings. Planta 2010, 231, 1301–1309. [Google Scholar] [CrossRef]
- Wang, H.R.; Che, Y.H.; Wang, Z.H.; Zhang, B.N.; Huang, D.; Feng, F.; Ao, H. The multiple effects of hydrogen sulfide on cadmium toxicity in tobacco may be interacted with CaM signal transduction. J. Hazard. Mat. 2021, 403, 123651. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, C.; Wang, X.; Liu, C.; Ding, X.; Ma, P.; Jia, H. Hydrogen sulfide regulates the activity of antioxidant enzymes through persulfidation and improves the resistance of tomato seedling to Copper Oxide nanoparticles (CuO NPs)-induced oxidative stress. Plant Physiol. Biochem. 2020, 156, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Siddiqui, M.H. Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide 2017, 68, 91–102. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Concentration | Experiment 1 | Experiment 2 |
|---|---|---|---|
| Control | 0 | Yes | Yes |
| Cadmium (Cd) | 200 mg Cd kg−1 soil | Yes | Yes |
| SNP | 100 µM SNP | Yes | No |
| ES | 200 mg S kg−1 soil | Yes | No |
| AS | 200 mg S kg−1 soil | Yes | No |
| SNP + Cd | 100 µM SNP + 200 mg Cd kg−1 soil | Yes | No |
| ES + Cd | 200 mg S kg−1 soil + 200 mg Cd kg−1 soil | Yes | No |
| AS + Cd | 200 mg S kg−1 soil + 200 mg Cd kg−1 soil | Yes | No |
| SNP + ES + Cd | 100 µM SNP +200 mg S kg−1 soil + 200 mg Cd kg−1 soil | Yes | Yes |
| SNP + AS + Cd | 100 µM SNP +200 mg S kg−1 soil + 200 mg Cd kg−1 soil | Yes | No |
| SNP + ES + Cd + HT | 100 µM + 200 mg S kg−1 soil + 200 mg Cd kg−1 soil + 100 µM HT | No | Yes |
| SNP + ES + Cd + cPTIO | 100 µM + 200 mg S kg−1 soil + 200 mg Cd kg−1 soil + 100 µM cPTIO | No | Yes |
| Treatment | SPAD Value | Net Photosynthesis (µmol CO2 m−2 s−1) | Intercellular CO2 Concentration (µmol CO2 mol−1) | Stomatal Conductance (mmol m−2 s−1) | Rubisco Activity (µmol CO2 mg−1 Protein min−1) |
|---|---|---|---|---|---|
| Control | 26.7 ± 1.3f | 22.2 ± 0.9f | 278.4 ± 13d | 231.4 ± 11.3i | 32.6 ± 1.6f |
| Cd | 16.8 ± 0.8g | 14.5 ± 0.6g | 197.2 ± 9.6e | 180.6 ± 8.8j | 21.3 ± 1.0g |
| SNP | 44.6 ± 2.23a | 29.6 ± 1.3a | 404.6 ± 19.8a | 568.5 ± 27.8a | 67.1 ± 3.4a |
| ES | 41.5 ± 2.0b | 28.4 ± 1.2b | 392.3 ± 18ab | 521.6 ± 25.8b | 61.4 ± 3.1b |
| AS | 40.9 ± 2b | 27.9 ± 1.3bc | 390.3 ± 19ab | 496.7 ± 24.6c | 59.3 ± 2.9b |
| SNP + Cd | 30.4 ± 1.5e | 24.4 ± 1.1e | 318.9 ± 15c | 342.6 ± 16.8f | 40.4 ± 2.0e |
| ES + Cd | 28.6 ± 1.4ef | 23.5 ± 1.1ef | 306.2 ± 15.3cd | 304.4 ± 14.8g | 39.7 ± 1.9e |
| AS + Cd | 28.14 ± 1.4ef | 23.2 ± 1.0ef | 303.4 ± 14.9cd | 298.8 ± 13.8h | 38.4 ± 1.9e |
| SNP + ES + Cd | 36.5 ± 1.8c | 26.3 ± 1.3c | 356.8 ± 17.2b | 438.7 ± 21.4d | 52.9 ± 2.6c |
| SNP + AS + Cd | 33.4 ± 1.7d | 25.6 ± 1.2d | 354.2 ± 16.8cd | 394.6 ± 19.3e | 46.3 ± 2.3f |
| Treatment | Fv/Fm | YII | ETR |
|---|---|---|---|
| Control | 0.78 ± 0.01d | 0.306 ± 0.01f | 83.3 ± 1.0e |
| Cd | 0.58 ± 0.01g | 0.188 ± 0.01g | 27.8 ± 1.0f |
| SNP | 0.86 ± 0.03a | 0.528 ± 0.03a | 188.3 ± 3.0a |
| ES | 0.82 ± 0.03b | 0.491 ± 0.02b | 172.8 ± 3.0b |
| AS | 0.81 ± 0.02c | 0.479 ± 0.01bc | 158.4 ± 3.0c |
| SNP + Cd | 0.67 ± 0.02ef | 0.405 ± 0.02d | 102.1 ± 3.0d |
| ES + Cd | 0.63 ± 0.02f | 0.372 ± 0.01e | 96.9 ± 2.1de |
| AS + Cd | 0.63 ± 0.02f | 0.368 ± 0.01e | 94.7 ± 2.0de |
| SNP + ES + Cd | 0.81 ± 0.03bc | 0.525 ± 0.03a | 155.4 ± 3.0c |
| SNP + AS + Cd | 0.71 ± 0.02d | 0.433 ± 0.01c | 119.8 ± 3.0bc |
| Treatment | ATP-S Activity | OAS-TL Activity | Sulfur Content | Cysteine Content | GSH Content |
|---|---|---|---|---|---|
| (U mg−1 Protein min−1) | (mg g−1 DW) | (nmol g−1 FW) | |||
| Control | 1.42 ± 0.07i | 0.53 ± 0.02h | 4.33 ± 0.19g | 18.33 ± 0.9i | 250.1 ± 12.5h |
| Cd | 1.86 ± 0.09h | 0.70 ± 0.03g | 3.09 ± 0.15h | 22.09 ± 1.1h | 189.6 ± 9.5i |
| SNP | 2.51 ± 0.12fg | 0.88 ± 0.04f | 5.47 ± 0.27f | 31.47 ± 1.5g | 511.4 ± 25.5c |
| ES | 2.81 ± 0.14f | 1.04 ± 0.05de | 9.64 ± 0.48a | 39.64 ± 1.9f | 570.6 ± 28.5a |
| AS | 2.39 ± 0.12fg | 1.19 ± 0.05d | 8.55 ± 0.42b | 38.55 ± 1.9f | 506.4 ± 25.3cd |
| SNP + Cd | 3.15 ± 0.15e | 1.02 ± 0.05de | 4.29 ± 0.21g | 49.29 ± 2.46de | 460.4 ± 23.1e |
| ES + Cd | 3.52 ± 0.17bc | 1.35 ± 0.07bc | 6.67 ± 0.33e | 55.89 ± 2.8c | 417.1 ± 20.8f |
| AS + Cd | 3.29 ± 0.16bcd | 1.21 ± 0.06d | 6.18 ± 0.3e | 51.67 ± 2.6d | 368.5 ± 18.4g |
| SNP + ES + Cd | 4.36 ± 0.21a | 1.67 ± 0.09a | 7.75 ± 0.38c | 65.81 ± 3.29a | 559.2 ± 27.9ab |
| SNP + AS + Cd | 3.88 ± 0.19b | 1.49 ± 0.08b | 7.38 ± 0.36cd | 60.38 ± 3.0b | 511.2 ± 25.5c |
| Treatment | Nitrogen Content | NR Activity | p-NUE | p-SUE | NPT Content | PCs Content |
|---|---|---|---|---|---|---|
| (mg g−1 DW) | (µmol NO2 mg−1 Protein h−1) | (g m−2) | (nmol g−1 FW) | |||
| Control | 40.2 ± 2.0f | 198.1 ± 9.9g | 45.3 ± 2.3g | 17.7 ± 0.8h | 344.3 ± 17.2g | 94.1 ± 4.7f |
| Cd | 27.5 ± 1.3g | 130.8 ± 6.5g | 32.6 ± 1.6h | 3.09 ± 0.15i | 407.5 ± 20.3f | 160.6 ± 8e |
| SNP | 75.6 ± 3.8a | 683.1 ± 34.1a | 39.2 ± 1.9b | 19.9 ± 0.95c | 588.7 ± 29.4e | 60.4 ± 3.0g |
| ES | 71.4 ± 3.6b | 581.2 ± 29.0b | 53.7 ± 2.7a | 35.5 ± 1.7a | 625.7 ± 31.2de | 55.6 ± 2.60g |
| AS | 66.9 ± 3.4c | 515.3 ± 25.7c | 41.7 ± 2.3b | 31.9 ± 1.6b | 567.8 ± 28.3e | 61.4 ± 3.1g |
| SNP + Cd | 56.6 ± 2.8d | 382.4 ± 19.1e | 43.5 ± 2.2f | 14.2 ± 0.6g | 676.4 ± 33.8d | 250.4 ± 12.5d |
| ES + Cd | 49.5 ± 2.5e | 338.6 ± 16.9ef | 47.5 ± 2.4e | 25.5 ± 1.23f | 711.6 ± 35.5c | 294.1 ± 14.7c |
| AS + Cd | 48.2 ± 2.4e | 332.1 ± 16.6ef | 38.3 ± 1.8f | 23.0 ± 1.1g | 621.8 ± 31.0de | 253.5 ± 12.8d |
| SNP + ES + Cd | 70.3 ± 3.5b | 577.0 ± 28.8b | 41.6 ± 2.3c | 25.9 ± 1.2d | 974.7 ± 48.7a | 415.2 ± 20.7a |
| SNP + AS + Cd | 64.8 ± 3.2cd | 421.3 ±21.1d | 39.8 ± 1.9d | 24.9 ± 1.2e | 833.1 ± 41.6b | 321.2 ± 16b |
| Treatment | NO Content (µg g−1 FW) | H2S Content (µg g−1 FW) | L-DES Activity (nmol mg−1 Protein) |
|---|---|---|---|
| Control | 3.08 ± 0.15g | 2.33 ± 0.11f | 10.71 ± 0.53i |
| Cd | 6.67 ± 0.33e | 5.83 ± 0.29c | 19.82 ± 0.99e |
| SNP | 9.56 ± 0.47c | 4.52 ± 0.22d | 15.52 ± 0.77f |
| ES | 5.62 ± 0.28f | 4.12 ± 0.21d | 14.83 ± 0.74fg |
| AS | 5.55 ± 0.27f | 4.08 ± 0.19de | 13.65 ± 0.68h |
| SNP + Cd | 9.81 ± 0.49c | 7.61 ± 0.38b | 28.61 ± 1.43c |
| ES + Cd | 8.19 ± 0.4d | 7.48 ± 0.34b | 24.50 ± 1.22d |
| AS + Cd | 8.15 ± 0.38d | 7.44 ± 0.33b | 24.01 ± 1.2d |
| SNP + ES + Cd | 13.7 ± 0.7a | 9.83 ± 0.48a | 34.29 ± 1.7a |
| SNP + AS + Cd | 11.6 ± 0.68b | 9.59 ± 0.45a | 31.520 + 1.57b |
| Treatment | Plant Dry Weight (g plant−1) | Net Photosynthesis (µmol CO2 m−2 s−1) | p-SUE (g m−2) | H2S Content (µg g−1 FW) | NO Content (µg g−1 FW) |
|---|---|---|---|---|---|
| Control | 2.16 ± 0.10b | 18.01 ± 0.90b | 16.85 ± 0.84b | 3.92 ± 0.19f | 4.33 ± 0.21f |
| Cd | 1.28 ± 0.06c | 12.78 ± 0.63c | 8.37 ± 0.41e | 5.37 ± 0.26g | 6.51 ± 0.32g |
| SNP + ES + Cd | 3.05 ± 0.15a | 26.57 ± 1.32a | 23.57 ± 1.17a | 10.56 ± 0.52a | 14.63 ± 0.73e |
| SNP + ES + Cd + HT | 1.08 ± 0.05d | 9.53 ± 0.47e | 9.08 ± 0.45d | 1.08 ± 0.05f | 2.69 ± 0.13d |
| SNP + ES + Cd + cPTIO | 1.25 ± 0.06c | 11.94 ± 0.6d | 12.65 ± 0.63c | 5.12 ± 0.23b | 1.65 ± 0.08a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mir, I.R.; Rather, B.A.; Masood, A.; Khan, N.A. Nitric Oxide- and Sulfur-Mediated Reversal of Cadmium-Inhibited Photosynthetic Performance Involves Hydrogen Sulfide and Regulation of Nitrogen, Sulfur, and Antioxidant Metabolism in Mustard. Stresses 2022, 2, 550-577. https://doi.org/10.3390/stresses2040037
Mir IR, Rather BA, Masood A, Khan NA. Nitric Oxide- and Sulfur-Mediated Reversal of Cadmium-Inhibited Photosynthetic Performance Involves Hydrogen Sulfide and Regulation of Nitrogen, Sulfur, and Antioxidant Metabolism in Mustard. Stresses. 2022; 2(4):550-577. https://doi.org/10.3390/stresses2040037
Chicago/Turabian StyleMir, Iqbal R., Bilal A. Rather, Asim Masood, and Nafees A. Khan. 2022. "Nitric Oxide- and Sulfur-Mediated Reversal of Cadmium-Inhibited Photosynthetic Performance Involves Hydrogen Sulfide and Regulation of Nitrogen, Sulfur, and Antioxidant Metabolism in Mustard" Stresses 2, no. 4: 550-577. https://doi.org/10.3390/stresses2040037