
Stem Cell Scaffolds for the Treatment of Spinal Cord Injury—A Review
 , , and
, , and
Abstract
:1. Introduction
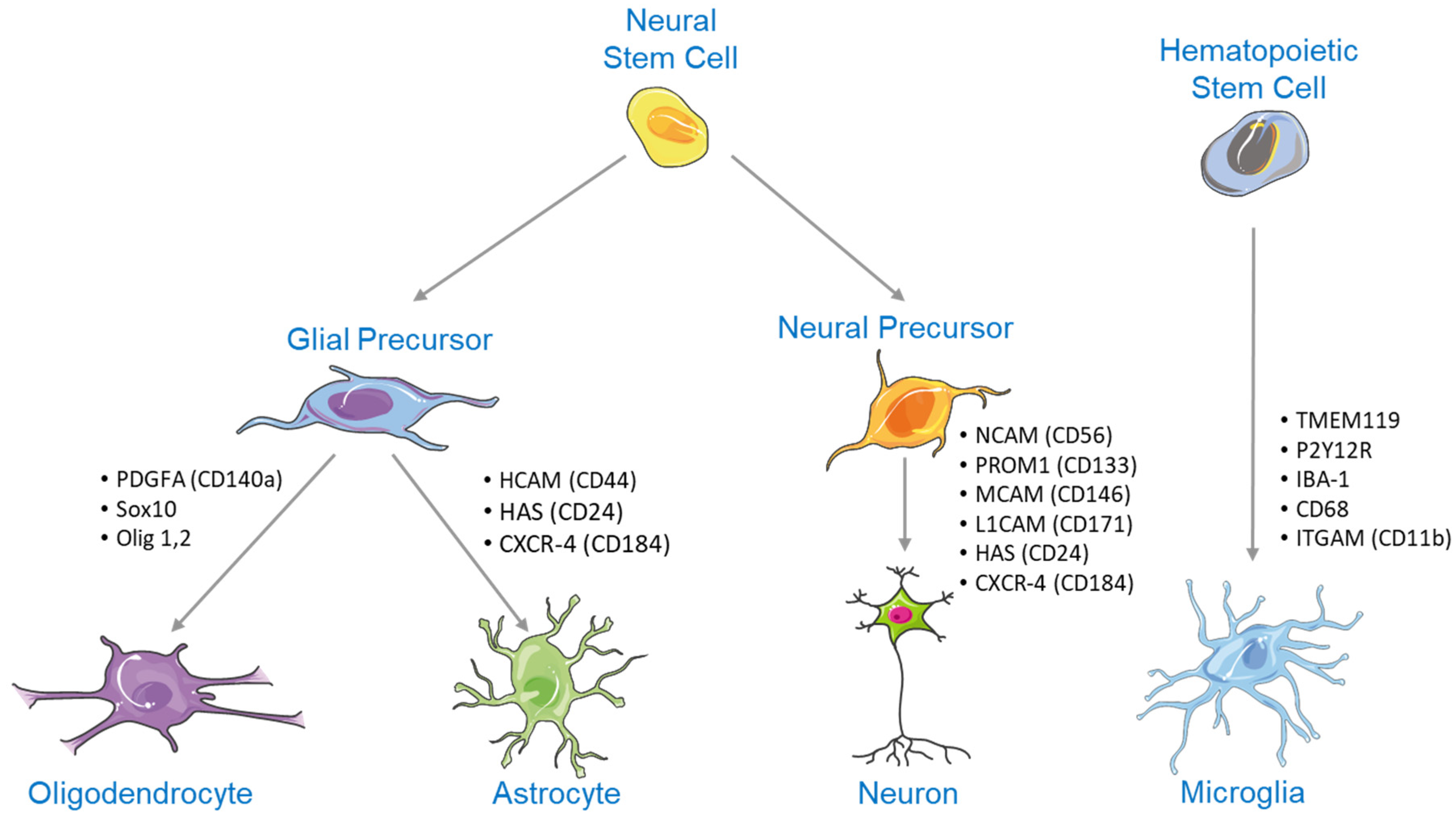
2. NSC Differentiation
2.1. Notch Signaling in NSC Differentiation
2.2. Wnt/β-Catenin Signaling in NSC Differentiation
2.3. Shh Signaling in NSC Differentiation
2.4. BMP Signaling in NSC Differentiation
3. Substrates Indicated for Axonal Regrowth Post-Injury
3.1. Plasma Membrane Sealants
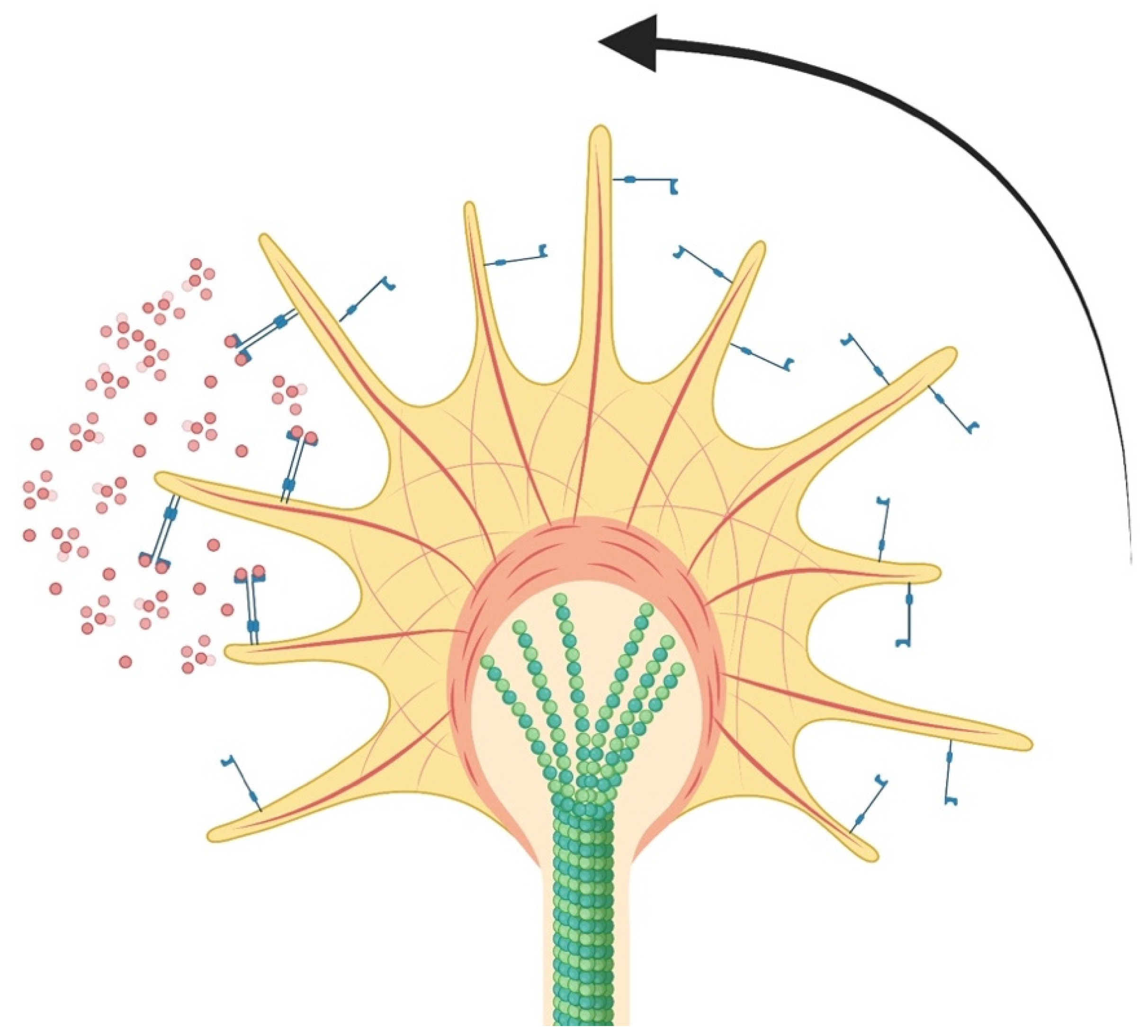
3.2. Growth Cone Formation and Stability
3.3. Neurotrophic Factors and Guidance
3.4. Matrix Vehicles for Axonal Regeneration
3.5. Electrical Stimulation for Axonal Growth
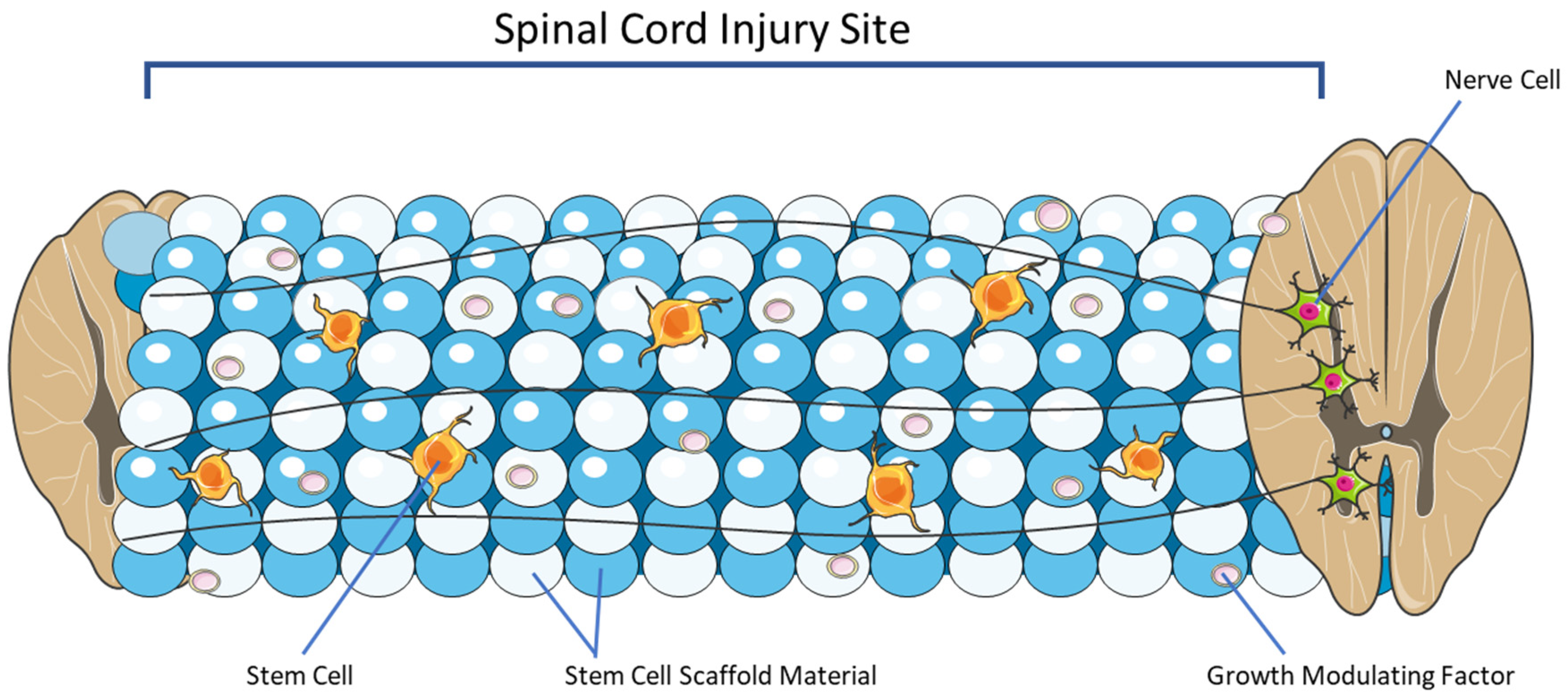
4. Overview of Stem Cell Scaffolding
4.1. Natural Polymer Scaffolds
4.2. Synthetic Polymer Scaffolds
4.3. Hydrogel Scaffolds
4.4. Hybrid or Composite Scaffolds
4.5. Growth Modulating Factors
5. Emerging Pre-Clinical Studies and Their Applications for Clinical Adoption
6. Conclusions and Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3D | Three dimensional |
| AIS | American Spinal Injury Association Impairment Scale |
| BDNF | Brain-derived neurotrophic factor |
| BMP | Bone morphogenetic protein |
| CNS | Central nervous system |
| CSF | Cerebrospinal fluid |
| ES | Electrical stimulation |
| GDNF | Glial cell line-derived neurotrophic factor |
| GFAP | Glial fibrillary acidic protein |
| GSK-3β | Glycogen-synthase-kinase-3β |
| GelMA | Gelatin methacryloyl |
| IMLL | Intramedullary lesion length |
| MAPK | Mitogenactivated protein kinase |
| MP | Methylprednisolone |
| MRI | Magnetic resonance imaging |
| MSC | Mesenchymal stem cell |
| MenSCs | Menstrual blood-derived mesenchymal stem cells |
| NCID | Notch intracellular signaling domain |
| NF-L | Neurofilament light |
| NGF | Nerve growth factor |
| NSC | Neural stem cell |
| NSPCs | Neural stem progenitor cells |
| NT3 | Neurotrophin-3 (NT3) |
| PI3K | Phosphatidylinositol 3-kinase |
| RAG | Recombination activating gene |
| SCI | Spinal cord injury |
| Shh | Sonic hedgehog |
| VEGF | Vascular endothelial growth factor |
References
- Barbiellini Amidei, C.; Salmaso, L.; Bellio, S.; Saia, M. Epidemiology of traumatic spinal cord injury: A large population-based study. Spinal Cord 2022, 60, 812–819. [Google Scholar] [CrossRef]
- Aarabi, B.; Albrecht, J.S.; Simard, J.M.; Chryssikos, T.; Schwartzbauer, G.; Sansur, C.A.; Crandall, K.; Gertner, M.; Howie, B.; Wessell, A.; et al. Trends in Demographics and Markers of Injury Severity in Traumatic Cervical Spinal Cord Injury. J. Neurotrauma 2021, 38, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, R.C.; Sterner, R.M. Immune response following traumatic spinal cord injury: Pathophysiology and therapies. Front. Immunol. 2022, 13, 1084101. [Google Scholar] [CrossRef]
- Gumy, L.F.; Tan, C.L.; Fawcett, J.W. The role of local protein synthesis and degradation in axon regeneration. Exp. Neurol. 2010, 223, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, B.K.; Shi, R. Mechanisms of neuronal membrane sealing following mechanical trauma. Neurosci. Bull. 2014, 30, 627–644. [Google Scholar] [CrossRef] [Green Version]
- Hur, E.M.; Saijilafu; Zhou, F.Q. Growing the growth cone: Remodeling the cytoskeleton to promote axon regeneration. Trends Neurosci. 2012, 35, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, S. Neurotrophic factors: Are they axon guidance molecules? Adv. Exp. Med. Biol. 2007, 621, 81–94. [Google Scholar] [CrossRef]
- Fiani, B.; Arshad, M.A.; Shaikh, E.S.; Baig, A.; Farooqui, M.; Ayub, M.A.; Zafar, A.; Quadri, S.A. Current updates on various treatment approaches in the early management of acute spinal cord injury. Rev. Neurosci. 2021, 32, 513–530. [Google Scholar] [CrossRef]
- Hsieh, Y.L.; Tay, J.; Hsu, S.H.; Chen, W.T.; Fang, Y.D.; Liew, C.Q.; Chou, E.H.; Wolfshohl, J.; d’Etienne, J.; Wang, C.H.; et al. Early versus Late Surgical Decompression for Traumatic Spinal Cord Injury on Neurological Recovery: A Systematic Review and Meta-Analysis. J. Neurotrauma 2021, 38, 2927–2936. [Google Scholar] [CrossRef]
- Badhiwala, J.H.; Wilson, J.R.; Witiw, C.D.; Harrop, J.S.; Vaccaro, A.R.; Aarabi, B.; Grossman, R.G.; Geisler, F.H.; Fehlings, M.G. The influence of timing of surgical decompression for acute spinal cord injury: A pooled analysis of individual patient data. Lancet Neurol. 2021, 20, 117–126. [Google Scholar] [CrossRef]
- Aarabi, B.; Akhtar-Danesh, N.; Chryssikos, T.; Shanmuganathan, K.; Schwartzbauer, G.T.; Simard, J.M.; Olexa, J.; Sansur, C.A.; Crandall, K.M.; Mushlin, H.; et al. Efficacy of Ultra-Early (<12 h), Early (12–24 h), and Late (>24–138.5 h) Surgery with Magnetic Resonance Imaging-Confirmed Decompression in American Spinal Injury Association Impairment Scale Grades A, B, and C Cervical Spinal Cord Injury. J. Neurotrauma 2020, 37, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Zarepour, A.; Hooshmand, S.; Gokmen, A.; Zarrabi, A.; Mostafavi, E. Spinal Cord Injury Management through the Combination of Stem Cells and Implantable 3D Bioprinted Platforms. Cells 2021, 10, 3189. [Google Scholar] [CrossRef] [PubMed]
- Blando, S.; Anchesi, I.; Mazzon, E.; Gugliandolo, A. Can a Scaffold Enriched with Mesenchymal Stem Cells Be a Good Treatment for Spinal Cord Injury? Int. J. Mol. Sci. 2022, 23, 7545. [Google Scholar] [CrossRef] [PubMed]
- Dorazco-Valdes, J. Estudio comparativo entre los aspectos clinicos, electroencefalograficos y la prueba del dibujo de la figura humana en el nino epileptico [Comparative study between the clinical aspects, the encephalographic and the draw-a-person test in epileptic children]. J. Neurol. Sci. 1968, 6, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Valdoz, J.C.; Johnson, B.C.; Jacobs, D.J.; Franks, N.A.; Dodson, E.L.; Sanders, C.; Cribbs, C.G.; Van Ry, P.M. The ECM: To Scaffold, or Not to Scaffold, That Is the Question. Int. J. Mol. Sci. 2021, 22, 12690. [Google Scholar] [CrossRef]
- Tang, Y.; Yu, P.; Cheng, L. Current progress in the derivation and therapeutic application of neural stem cells. Cell Death Dis. 2017, 8, e3108. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, A.; Radoszkiewicz, K.; Rybkowska, P.; Wedzinska, A.; Sarnowska, A. Interaction of Neural Stem Cells (NSCs) and Mesenchymal Stem Cells (MSCs) as a Promising Approach in Brain Study and Nerve Regeneration. Cells 2022, 11, 1464. [Google Scholar] [CrossRef]
- de Vasconcelos, P.; Lacerda, J.F. Hematopoietic Stem Cell Transplantation for Neurological Disorders: A Focus on Inborn Errors of Metabolism. Front. Cell. Neurosci. 2022, 16, 895511. [Google Scholar] [CrossRef]
- De Gioia, R.; Biella, F.; Citterio, G.; Rizzo, F.; Abati, E.; Nizzardo, M.; Bresolin, N.; Comi, G.P.; Corti, S. Neural Stem Cell Transplantation for Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 3103. [Google Scholar] [CrossRef]
- Navarro Quiroz, E.; Navarro Quiroz, R.; Ahmad, M.; Gomez Escorcia, L.; Villarreal, J.L.; Fernandez Ponce, C.; Aroca Martinez, G. Cell Signaling in Neuronal Stem Cells. Cells 2018, 7, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penton, A.L.; Leonard, L.D.; Spinner, N.B. Notch signaling in human development and disease. Semin. Cell Dev. Biol. 2012, 23, 450–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzari-Tavakoli, A.; Babajani, A.; Farjoo, M.H.; Hajinasrollah, M.; Bahrami, S.; Niknejad, H. The Cross-Talks Among Bone Morphogenetic Protein (BMP) Signaling and Other Prominent Pathways Involved in Neural Differentiation. Front. Mol. Neurosci. 2022, 15, 827275. [Google Scholar] [CrossRef] [PubMed]
- Teodorczyk, M.; Schmidt, M.H.H. Notching on Cancer’s Door: Notch Signaling in Brain Tumors. Front. Oncol. 2014, 4, 341. [Google Scholar] [CrossRef] [Green Version]
- Imayoshi, I.; Kageyama, R. The role of Notch signaling in adult neurogenesis. Mol. Neurobiol. 2011, 44, 7–12. [Google Scholar] [CrossRef]
- Nian, F.S.; Hou, P.S. Evolving Roles of Notch Signaling in Cortical Development. Front. Neurosci. 2022, 16, 844410. [Google Scholar] [CrossRef]
- Yamamoto, S.; Nagao, M.; Sugimori, M.; Kosako, H.; Nakatomi, H.; Yamamoto, N.; Takebayashi, H.; Nabeshima, Y.; Kitamura, T.; Weinmaster, G.; et al. Transcription factor expression and Notch-dependent regulation of neural progenitors in the adult rat spinal cord. J. Neurosci. 2001, 21, 9814–9823. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, Q.; Tian, H.; Lv, W.; Song, L.; Li, Z.; Yao, H.; Shi, S. Electroacupuncture in promoting neural repair after spinal cord injury: Inhibiting the Notch signaling pathway and regulating downstream proteins expression. Anat. Rec. 2021, 304, 2494–2505. [Google Scholar] [CrossRef]
- Yuan, H.; Fang, C.L.; Deng, Y.P.; Huang, J.; Niu, R.Z.; Chen, J.L.; Chen, T.B.; Zhu, Z.Q.; Chen, L.; Xiong, L.L.; et al. A2B5-positive oligodendrocyte precursor cell transplantation improves neurological deficits in rats following spinal cord contusion associated with changes in expression of factors involved in the Notch signaling pathway. Neurochirurgie 2022, 68, 188–195. [Google Scholar] [CrossRef]
- Chen, Y.; Lian, X.H.; Liao, L.Y.; Liu, Y.T.; Liu, S.L.; Gao, Q. Transplantation of bone marrow mesenchymal stem cells alleviates spinal cord injury via inhibiting Notch signaling. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 31–38. [Google Scholar] [CrossRef]
- Qian, D.; Li, L.; Rong, Y.; Liu, W.; Wang, Q.; Zhou, Z.; Gu, C.; Huang, Y.; Zhao, X.; Chen, J.; et al. Blocking Notch signal pathway suppresses the activation of neurotoxic A1 astrocytes after spinal cord injury. Cell Cycle 2019, 18, 3010–3029. [Google Scholar] [CrossRef] [PubMed]
- Toledo, E.M.; Colombres, M.; Inestrosa, N.C. Wnt signaling in neuroprotection and stem cell differentiation. Prog. Neurobiol. 2008, 86, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.Y.; Nusse, R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [Green Version]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, K.; Semenov, M.; Kato, Y.; Spokony, R.; Liu, C.; Katsuyama, Y.; Hess, F.; Saint-Jeannet, J.P.; He, X. LDL-receptor-related proteins in Wnt signal transduction. Nature 2000, 407, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.P.; Coulter, M.; Miotke, J.; Meyer, R.L.; Takemaru, K.; Levine, J.M. Abrogation of beta-catenin signaling in oligodendrocyte precursor cells reduces glial scarring and promotes axon regeneration after CNS injury. J. Neurosci. 2014, 34, 10285–10297. [Google Scholar] [CrossRef] [Green Version]
- Strand, N.S.; Hoi, K.K.; Phan, T.M.T.; Ray, C.A.; Berndt, J.D.; Moon, R.T. Wnt/beta-catenin signaling promotes regeneration after adult zebrafish spinal cord injury. Biochem. Biophys. Res. Commun. 2016, 477, 952–956. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Liu, Y.; Sun, L.; Fu, M.; Zhao, Y. Salvianolic acid B activates Wnt/beta-catenin signaling following spinal cord injury. Exp. Ther. Med. 2020, 19, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Zhang, T.; Wang, F.; Lv, C. Therapeutic Potential of Wnt-3a in Neurological Recovery after Spinal Cord Injury. Eur. Neurol. 2019, 81, 197–204. [Google Scholar] [CrossRef]
- Cui, Y.; Yin, Y.; Xiao, Z.; Zhao, Y.; Chen, B.; Yang, B.; Xu, B.; Song, H.; Zou, Y.; Ma, X.; et al. LncRNA Neat1 mediates miR-124-induced activation of Wnt/beta-catenin signaling in spinal cord neural progenitor cells. Stem Cell Res. Ther. 2019, 10, 400. [Google Scholar] [CrossRef]
- Zhu, K.; Xu, W.; Han, D.; Mei, X. Effect of Sirtuin-1 and Wnt/beta-Catenin Signaling Pathway in Rat Model of Spinal Cord Injury. Comput. Math. Methods Med. 2022, 2022, 1799607. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Wang, Y.S.; Yuan, Y.J.; Wan, Z.H.; Yao, T.C.; Li, H.H.; Tang, P.F.; Mei, X.F. Neuroprotective effect of rapamycin on spinal cord injury via activation of the Wnt/beta-catenin signaling pathway. Neural Regen. Res. 2015, 10, 951–957. [Google Scholar] [CrossRef]
- Komada, M. Sonic hedgehog signaling coordinates the proliferation and differentiation of neural stem/progenitor cells by regulating cell cycle kinetics during development of the neocortex. Congenit. Anom. 2012, 52, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Ihrie, R.A.; Shah, J.K.; Harwell, C.C.; Levine, J.H.; Guinto, C.D.; Lezameta, M.; Kriegstein, A.R.; Alvarez-Buylla, A. Persistent sonic hedgehog signaling in adult brain determines neural stem cell positional identity. Neuron 2011, 71, 250–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charytoniuk, D.; Porcel, B.; Rodriguez Gomez, J.; Faure, H.; Ruat, M.; Traiffort, E. Sonic Hedgehog signalling in the developing and adult brain. J. Physiol. 2002, 96, 9–16. [Google Scholar] [CrossRef]
- Petralia, R.S.; Schwartz, C.M.; Wang, Y.X.; Mattson, M.P.; Yao, P.J. Subcellular localization of Patched and Smoothened, the receptors for Sonic hedgehog signaling, in the hippocampal neuron. J. Comp. Neurol. 2011, 519, 3684–3699. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Wang, L.; Tang, F.; Guo, S.; Liao, H.; Fan, C.; Yang, Q. Resveratrol-mediated neurorestoration after cerebral ischemic injury—Sonic Hedgehog signaling pathway. Life Sci. 2021, 280, 119715. [Google Scholar] [CrossRef]
- Al-Sammarraie, N.; Ray, S.K. Bone morphogenic protein signaling in spinal cord injury. Neuroimmunol. Neuroinflamm. 2021, 8, 53–63. [Google Scholar] [CrossRef]
- Cole, A.E.; Murray, S.S.; Xiao, J. Bone Morphogenetic Protein 4 Signalling in Neural Stem and Progenitor Cells during Development and after Injury. Stem Cells Int. 2016, 2016, 9260592. [Google Scholar] [CrossRef] [Green Version]
- Hart, C.G.; Karimi-Abdolrezaee, S. Bone morphogenetic proteins: New insights into their roles and mechanisms in CNS development, pathology and repair. Exp. Neurol. 2020, 334, 113455. [Google Scholar] [CrossRef]
- Mira, H.; Andreu, Z.; Suh, H.; Lie, D.C.; Jessberger, S.; Consiglio, A.; San Emeterio, J.; Hortiguela, R.; Marques-Torrejon, M.A.; Nakashima, K.; et al. Signaling through BMPR-IA regulates quiescence and long-term activity of neural stem cells in the adult hippocampus. Cell Stem Cell 2010, 7, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.; Zou, H. BMP signaling in axon regeneration. Curr. Opin. Neurobiol. 2014, 27, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Hart, C.G.; Dyck, S.M.; Kataria, H.; Alizadeh, A.; Nagakannan, P.; Thliveris, J.A.; Eftekharpour, E.; Karimi-Abdolrezaee, S. Acute upregulation of bone morphogenetic protein-4 regulates endogenous cell response and promotes cell death in spinal cord injury. Exp. Neurol. 2020, 325, 113163. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, I.; Taniguchi, J.; Hata, K.; Saeki, N.; Yamashita, T. BMP inhibition enhances axonal growth and functional recovery after spinal cord injury. J. Neurochem. 2008, 105, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Huebner, E.A.; Strittmatter, S.M. Axon regeneration in the peripheral and central nervous systems. In Results and Problems in Cell Differentiation; Springer: Berlin/Heidelberg, Germany, 2009; Volume 48, pp. 339–351. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Borgens, R.B. Polymer and nano-technology applications for repair and reconstruction of the central nervous system. Exp. Neurol. 2012, 233, 126–144. [Google Scholar] [CrossRef]
- Jin, X. Membrane resealing as a promising strategy for early treatment of neurotrauma. Neural Regen. Res. 2014, 9, 1876–1877. [Google Scholar] [CrossRef] [Green Version]
- Vargas, S.A.; Bittner, G.D. Natural mechanisms and artificial PEG-induced mechanism that repair traumatic damage to the plasmalemma in eukaryotes. Curr. Top. Membr. 2019, 84, 129–167. [Google Scholar] [CrossRef]
- Ghergherehchi, C.L.; Mikesh, M.; Sengelaub, D.R.; Jackson, D.M.; Smith, T.; Nguyen, J.; Shores, J.T.; Bittner, G.D. Polyethylene glycol (PEG) and other bioactive solutions with neurorrhaphy for rapid and dramatic repair of peripheral nerve lesions by PEG-fusion. J. Neurosci. Methods 2019, 314, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ghergherehchi, C.L.; Shores, J.T.; Alderete, J.; Weitzel, E.K.; Bittner, G.D. Methylene blue enhances polyethylene glycol-fusion repair of completely severed rat sciatic nerves. Neural Regen. Res. 2021, 16, 2056–2063. [Google Scholar] [CrossRef]
- Mikesh, M.; Ghergherehchi, C.L.; Hastings, R.L.; Ali, A.; Rahesh, S.; Jagannath, K.; Sengelaub, D.R.; Trevino, R.C.; Jackson, D.M.; Bittner, G.D. Polyethylene glycol solutions rapidly restore and maintain axonal continuity, neuromuscular structures, and behaviors lost after sciatic nerve transections in female rats. J. Neurosci. Res. 2018, 96, 1223–1242. [Google Scholar] [CrossRef] [Green Version]
- Kahn, O.I.; Baas, P.W. Microtubules and Growth Cones: Motors Drive the Turn. Trends Neurosci. 2016, 39, 433–440. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erturk, A.; Hellal, F.; Enes, J.; Bradke, F. Disorganized microtubules underlie the formation of retraction bulbs and the failure of axonal regeneration. J. Neurosci. 2007, 27, 9169–9180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Li, X.; Zhao, Y.; Xiao, Z.; Xue, W.; Sun, J.; Li, X.; Zhuang, Y.; Chen, Y.; Dai, J. Cetuximab and Taxol co-modified collagen scaffolds show combination effects for the repair of acute spinal cord injury. Biomater. Sci. 2018, 6, 1723–1734. [Google Scholar] [CrossRef]
- Pastuhov, S.I.; Hisamoto, N.; Matsumoto, K. MAP kinase cascades regulating axon regeneration in C. elegans. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Auerbach, J.M.; Rodríguez-Gómez, J.A.; Velasco, I.; Gavin, D.; Lumelsky, N.; Lee, S.-H.; Nguyen, J.; Sánchez-Pernaute, R.; Bankiewicz, K.; et al. Dopamine neurons derived from embryonic stem cells function in an animal model of Parkinson’s disease. Nature 2002, 418, 50–56. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Marlin, M.C.; Li, G. Biogenesis and function of the NGF/TrkA signaling endosome. Int. Rev. Cell Mol. Biol. 2015, 314, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Sasaki, Y.; Wen, Z.; Bassell, G.J.; Zheng, J.Q. An essential role for beta-actin mRNA localization and translation in Ca2+-dependent growth cone guidance. Nat. Neurosci. 2006, 9, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Difato, F.; Tsushima, H.; Pesce, M.; Benfenati, F.; Blau, A.; Chieregatti, E. The formation of actin waves during regeneration after axonal lesion is enhanced by BDNF. Sci. Rep. 2011, 1, 183. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, N.; Katsuno, H. Actin Waves: Origin of Cell Polarization and Migration? Trends Cell Biol. 2017, 27, 515–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, G.; Letourneau, P.C. Localized sources of neurotrophins initiate axon collateral sprouting. J. Neurosci. 1998, 18, 5403–5414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bove, M.; Monto, F.; Guillem-Llobat, P.; Ivorra, M.D.; Noguera, M.A.; Zambrano, A.; Sirerol-Piquer, M.S.; Requena, A.C.; Garcia-Alonso, M.; Tejerina, T.; et al. NT3/TrkC Pathway Modulates the Expression of UCP-1 and Adipocyte Size in Human and Rodent Adipose Tissue. Front. Endocrinol. 2021, 12, 630097. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Tuszynski, M.H. Growth factors and combinatorial therapies for CNS regeneration. Exp. Neurol. 2008, 209, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.R.; Kraus, K.; Batter, D.K.; Blunt, D.G.; Weremowitz, J.; Lynch, S.E.; Antoniades, H.N.; Hansson, H.A. Gel matrix vehicles for growth factor application in nerve gap injuries repaired with tubes: A comparison of biomatrix, collagen, and methylcellulose. Exp. Neurol. 1997, 146, 395–402. [Google Scholar] [CrossRef]
- Yoshii, S.; Oka, M.; Shima, M.; Taniguchi, A.; Akagi, M. Bridging a 30-mm nerve defect using collagen filaments. J. Biomed. Mater. Res. A 2003, 67, 467–474. [Google Scholar] [CrossRef]
- de la Torre, J.C. Catecholamine fiber regeneration across a collagen bioimplant after spinal cord transection. Brain Res. Bull. 1982, 9, 545–552. [Google Scholar] [CrossRef]
- Joosten, E.A. Biodegradable biomatrices and bridging the injured spinal cord: The corticospinal tract as a proof of principle. Cell Tissue Res. 2012, 349, 375–395. [Google Scholar] [CrossRef]
- Lu, X.; Perera, T.H.; Aria, A.B.; Callahan, L.A.S. Polyethylene glycol in spinal cord injury repair: A critical review. J. Exp. Pharmacol. 2018, 10, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.; Bento, A.R.; Barros, D.; Laundos, T.L.; Sousa, S.R.; Quelhas, P.; Sousa, M.M.; Pego, A.P.; Amaral, I.F. Fibrin functionalization with synthetic adhesive ligands interacting with alpha6beta1 integrin receptor enhance neurite outgrowth of embryonic stem cell-derived neural stem/progenitors. Acta. Biomater. 2017, 59, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, T.M.; Baron-Heeris, D.; Houwers, I.G.J.; Keenan, R.; Williams, R.J.; Nisbet, D.R.; Harvey, A.R.; Hodgetts, S.I. Peptide Hydrogel Scaffold for Mesenchymal Precursor Cells Implanted to Injured Adult Rat Spinal Cord. Tissue Eng. Part A 2021, 27, 993–1007. [Google Scholar] [CrossRef]
- McGregor, C.E.; English, A.W. The Role of BDNF in Peripheral Nerve Regeneration: Activity-Dependent Treatments and Val66Met. Front. Cell. Neurosci. 2018, 12, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Majed, A.A.; Brushart, T.M.; Gordon, T. Electrical stimulation accelerates and increases expression of BDNF and trkB mRNA in regenerating rat femoral motoneurons. Eur. J. Neurosci. 2000, 12, 4381–4390. [Google Scholar] [PubMed]
- Kawamura, K.; Kano, Y. Electrical stimulation induces neurite outgrowth in PC12m3 cells via the p38 mitogen-activated protein kinase pathway. Neurosci. Lett. 2019, 698, 81–84. [Google Scholar] [CrossRef]
- Geremia, N.M.; Gordon, T.; Brushart, T.M.; Al-Majed, A.A.; Verge, V.M. Electrical stimulation promotes sensory neuron regeneration and growth-associated gene expression. Exp. Neurol. 2007, 205, 347–359. [Google Scholar] [CrossRef]
- Keane, G.C.; Pan, D.; Roh, J.; Larson, E.L.; Schellhardt, L.; Hunter, D.A.; Snyder-Warwick, A.K.; Moore, A.M.; Mackinnon, S.E.; Wood, M.D. The Effects of Intraoperative Electrical Stimulation on Regeneration and Recovery After Nerve Isograft Repair in a Rat Model. Hand 2022, 17, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.; Schellhardt, L.; Keane, G.C.; Hunter, D.A.; Moore, A.M.; Snyder-Warwick, A.K.; Mackinnon, S.E.; Wood, M.D. Short-Duration, Pulsatile, Electrical Stimulation Therapy Accelerates Axon Regeneration and Recovery following Tibial Nerve Injury and Repair in Rats. Plast. Reconstr. Surg. 2022, 149, 681e–690e. [Google Scholar] [CrossRef]
- Koh, G.P.; Fouad, C.; Lanzinger, W.; Willits, R.K. Effect of Intraoperative Electrical Stimulation on Recovery after Rat Sciatic Nerve Isograft Repair. Neurotrauma. Rep. 2020, 1, 181–191. [Google Scholar] [CrossRef]
- Juckett, L.; Saffari, T.M.; Ormseth, B.; Senger, J.L.; Moore, A.M. The Effect of Electrical Stimulation on Nerve Regeneration Following Peripheral Nerve Injury. Biomolecules 2022, 12, 1856. [Google Scholar] [CrossRef]
- Abdollahiyan, P.; Oroojalian, F.; Mokhtarzadeh, A. The triad of nanotechnology, cell signalling, and scaffold implantation for the successful repair of damaged organs: An overview on soft-tissue engineering. J. Control. Release 2021, 332, 460–492. [Google Scholar] [CrossRef]
- Golland, B.; Tipper, J.L.; Hall, R.M.; Tronci, G.; Russell, S.J. A Biomimetic Nonwoven-Reinforced Hydrogel for Spinal Cord Injury Repair. Polymers 2022, 14, 4376. [Google Scholar] [CrossRef]
- Gao, C.; Li, Y.; Liu, X.; Huang, J.; Zhang, Z. 3D bioprinted conductive spinal cord biomimetic scaffolds for promoting neuronal differentiation of neural stem cells and repairing of spinal cord injury. Chem. Eng. J. 2023, 451, 138788. [Google Scholar] [CrossRef]
- Zeng, X.; Zeng, Y.S.; Ma, Y.H.; Lu, L.Y.; Du, B.L.; Zhang, W.; Li, Y.; Chan, W.Y. Bone marrow mesenchymal stem cells in a three-dimensional gelatin sponge scaffold attenuate inflammation, promote angiogenesis, and reduce cavity formation in experimental spinal cord injury. Cell Transpl. 2011, 20, 1881–1899. [Google Scholar] [CrossRef] [PubMed]
- Bruzauskaite, I.; Bironaite, D.; Bagdonas, E.; Bernotiene, E. Scaffolds and cells for tissue regeneration: Different scaffold pore sizes-different cell effects. Cytotechnology 2016, 68, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karageorgiou, V.; Kaplan, D. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 2005, 26, 5474–5491. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Huang, H.; Niu, J.; Chen, C.; Weng, J.; Yu, F.; Wang, D.; Kang, B.; Wang, T.; Yuan, G.; et al. Exploring the interconnectivity of biomimetic hierarchical porous Mg scaffolds for bone tissue engineering: Effects of pore size distribution on mechanical properties, degradation behavior and cell migration ability. J. Magnes. Alloy. 2021, 9, 1954–1966. [Google Scholar] [CrossRef]
- Zhao, H.; Li, L.; Ding, S.; Liu, C.; Ai, J. Effect of porous structure and pore size on mechanical strength of 3D-printed comby scaffolds. Mater. Lett. 2018, 223, 21–24. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, H.N.; Lee, S.J.; Song, J.E.; Kwon, S.Y.; Chung, J.W.; Lee, D.; Khang, G. Effect of pore sizes of PLGA scaffolds on mechanical properties and cell behaviour for nucleus pulposus regeneration in vivo. J. Tissue Eng. Regen. Med. 2017, 11, 44–57. [Google Scholar] [CrossRef]
- Liu, H.; Feng, Y.; Che, S.; Guan, L.; Yang, X.; Zhao, Y.; Fang, L.; Zvyagin, A.V.; Lin, Q. An Electroconductive Hydrogel Scaffold with Injectability and Biodegradability to Manipulate Neural Stem Cells for Enhancing Spinal Cord Injury Repair. Biomacromolecules 2023, 24, 86–97. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, Y.; Liu, W. Motor functional recovery efficacy of scaffolds with bone marrow stem cells in rat spinal cord injury: A Bayesian network meta-analysis. Spinal Cord 2023, 61, 93–98. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, B.; Ding, J.; Yan, L.; Thawani, J.P.; Fu, C.; Chen, X. Polymer scaffolds facilitate spinal cord injury repair. Acta Biomater. 2019, 88, 57–77. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liang, Z.; Yang, L.; Du, W.; Yu, T.; Tang, H.; Li, C.; Qiu, H. The role of natural polymers in bone tissue engineering. J. Control. Release 2021, 338, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Mecham, R.P. Overview of extracellular matrix. Curr. Protoc. Cell Biol. 2012, 57, 10–11. [Google Scholar] [CrossRef]
- Zhao, P.; Gu, H.; Mi, H.; Rao, C.; Fu, J.; Turng, L.-S. Fabrication of scaffolds in tissue engineering: A review. Front. Mech. Eng. 2017, 13, 107–119. [Google Scholar] [CrossRef]
- Mneimneh, A.T.; Mehanna, M.M. Collagen-based scaffolds: An auspicious tool to support repair, recovery, and regeneration post spinal cord injury. Int. J. Pharm. 2021, 601, 120559. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Yang, H.; Zhao, Y.; Li, T.; Xin, D.; Gai, C.; Jiang, Z.; Wang, Z. 3D Gelatin Microsphere Scaffolds Promote Functional Recovery after Spinal Cord Hemisection in Rats. Adv. Sci. 2023, 10, e2204528. [Google Scholar] [CrossRef] [PubMed]
- Gardin, C.; Vindigni, V.; Bressan, E.; Ferroni, L.; Nalesso, E.; Puppa, A.D.; D’Avella, D.; Lops, D.; Pinton, P.; Zavan, B. Hyaluronan and fibrin biomaterial as scaffolds for neuronal differentiation of adult stem cells derived from adipose tissue and skin. Int. J. Mol. Sci. 2011, 12, 6749–6764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, A.C.; Fideles, S.O.M.; Pomini, K.T.; Bellini, M.Z.; Pereira, E.; Reis, C.H.B.; Pilon, J.P.G.; de Marchi, M.A.; Trazzi, B.F.M.; da Silva, W.S.; et al. Potential of Fibrin Glue and Mesenchymal Stem Cells (MSCs) to Regenerate Nerve Injuries: A Systematic Review. Cells 2022, 11, 221. [Google Scholar] [CrossRef]
- Rose, J.C.; Camara-Torres, M.; Rahimi, K.; Kohler, J.; Moller, M.; De Laporte, L. Nerve Cells Decide to Orient inside an Injectable Hydrogel with Minimal Structural Guidance. Nano Lett. 2017, 17, 3782–3791. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, H.Q.; Chao, X.; Xu, W.X.; Liu, Y.; Ling, G.X.; Zhang, P. Multimodal therapy strategies based on hydrogels for the repair of spinal cord injury. Mil. Med. Res. 2022, 9, 16. [Google Scholar] [CrossRef]
- Macaya, D.; Spector, M. Injectable hydrogel materials for spinal cord regeneration: A review. Biomed. Mater. 2012, 7, 012001. [Google Scholar] [CrossRef] [PubMed]
- El-Sherbiny, I.M.; Yacoub, M.H. Hydrogel scaffolds for tissue engineering: Progress and challenges. Glob. Cardiol. Sci. Pract. 2013, 2013, 316–342. [Google Scholar] [CrossRef] [Green Version]
- Zamanifard, M.; Khorasani, M.T.; Daliri, M.; Parvazinia, M. Preparation and modeling of electrospun polyhydroxybutyrate/polyaniline composite scaffold modified by plasma and printed by an inkjet method and its cellular study. J. Biomater. Sci. Polym. Ed. 2020, 31, 1515–1537. [Google Scholar] [CrossRef]
- Mungenast, L.; Zuger, F.; Selvi, J.; Faia-Torres, A.B.; Ruhe, J.; Suter-Dick, L.; Gullo, M.R. Directional Submicrofiber Hydrogel Composite Scaffolds Supporting Neuron Differentiation and Enabling Neurite Alignment. Int. J. Mol. Sci. 2022, 23, 11525. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Li, Y.; Huang, J.; Cheng, S.; Zhang, Z. Inhibited astrocytic differentiation in neural stem cell-laden 3D bioprinted conductive composite hydrogel scaffolds for repair of spinal cord injury. Biomater. Adv. 2023, 148, 213385. [Google Scholar] [CrossRef]
- Kirschenbaum, B.; Goldman, S.A. Brain-derived neurotrophic factor promotes the survival of neurons arising from the adult rat forebrain subependymal zone. Proc. Natl. Acad. Sci. USA 1995, 92, 210–214. [Google Scholar] [CrossRef]
- Pan, S.; Qi, Z.; Li, Q.; Ma, Y.; Fu, C.; Zheng, S.; Kong, W.; Liu, Q.; Yang, X. Graphene oxide-PLGA hybrid nanofibres for the local delivery of IGF-1 and BDNF in spinal cord repair. Artif. Cells Nanomed. Biotechnol. 2019, 47, 651–664. [Google Scholar] [CrossRef]
- Walker, M.J.; Xu, X.M. History of Glial Cell Line-Derived Neurotrophic Factor (GDNF) and Its Use for Spinal Cord Injury Repair. Brain Sci. 2018, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veneruso, V.; Rossi, F.; Villella, A.; Bena, A.; Forloni, G.; Veglianese, P. Stem cell paracrine effect and delivery strategies for spinal cord injury regeneration. J. Control. Release 2019, 300, 141–153. [Google Scholar] [CrossRef]
- Hassannejad, Z.; Zadegan, S.A.; Vaccaro, A.R.; Rahimi-Movaghar, V.; Sabzevari, O. Biofunctionalized peptide-based hydrogel as an injectable scaffold for BDNF delivery can improve regeneration after spinal cord injury. Injury 2019, 50, 278–285. [Google Scholar] [CrossRef]
- Tollemar, V.; Collier, Z.J.; Mohammed, M.K.; Lee, M.J.; Ameer, G.A.; Reid, R.R. Stem cells, growth factors and scaffolds in craniofacial regenerative medicine. Genes Dis. 2016, 3, 56–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, J.D.; Luoma, J.E.; Anseth, K.S. Covalently tethered transforming growth factor beta in PEG hydrogels promotes chondrogenic differentiation of encapsulated human mesenchymal stem cells. Drug Deliv. Transl. Res. 2012, 2, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Fu, C.; Xiong, F.; He, C.; Wei, Q. Stem Cell Therapy for Spinal Cord Injury. Cell Transplant. 2021, 30, 963689721989266. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Moradi, L.; Katebi, M.; Ai, J.; Azami, M.; Moradveisi, B.; Ostad, S.N. Delivery of injectable thermo-sensitive hydrogel releasing nerve growth factor for spinal cord regeneration in rat animal model. J. Tissue Viability 2020, 29, 359–366. [Google Scholar] [CrossRef]
- Yao, X.; Sun, C.; Fan, B.; Zhao, C.; Zhang, Y.; Duan, H.; Pang, Y.; Shen, W.; Li, B.; Wang, X.; et al. Neurotropin exerts neuroprotective effects after spinal cord injury by inhibiting apoptosis and modulating cytokines. J. Orthop. Translat. 2021, 26, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Kataria, H.; Alizadeh, A.; Karimi-Abdolrezaee, S. Neuregulin-1/ErbB network: An emerging modulator of nervous system injury and repair. Prog. Neurobiol. 2019, 180, 101643. [Google Scholar] [CrossRef]
- Liu, W.; Xu, B.; Xue, W.; Yang, B.; Fan, Y.; Chen, B.; Xiao, Z.; Xue, X.; Sun, Z.; Shu, M.; et al. A functional scaffold to promote the migration and neuronal differentiation of neural stem/progenitor cells for spinal cord injury repair. Biomaterials 2020, 243, 119941. [Google Scholar] [CrossRef]
- Kourgiantaki, A.; Tzeranis, D.S.; Karali, K.; Georgelou, K.; Bampoula, E.; Psilodimitrakopoulos, S.; Yannas, I.V.; Stratakis, E.; Sidiropoulou, K.; Charalampopoulos, I.; et al. Neural stem cell delivery via porous collagen scaffolds promotes neuronal differentiation and locomotion recovery in spinal cord injury. NPJ Regen. Med. 2020, 5, 12. [Google Scholar] [CrossRef]
- Deng, W.S.; Ma, K.; Liang, B.; Liu, X.Y.; Xu, H.Y.; Zhang, J.; Shi, H.Y.; Sun, H.T.; Chen, X.Y.; Zhang, S. Collagen scaffold combined with human umbilical cord-mesenchymal stem cells transplantation for acute complete spinal cord injury. Neural Regen. Res. 2020, 15, 1686–1700. [Google Scholar] [CrossRef]
- Liu, S.; Xie, Y.Y.; Wang, L.D.; Tai, C.X.; Chen, D.; Mu, D.; Cui, Y.Y.; Wang, B. A multi-channel collagen scaffold loaded with neural stem cells for the repair of spinal cord injury. Neural Regen. Res. 2021, 16, 2284–2292. [Google Scholar] [CrossRef]
- Deng, W.S.; Yang, K.; Liang, B.; Liu, Y.F.; Chen, X.Y.; Zhang, S. Collagen/heparin sulfate scaffold combined with mesenchymal stem cells treatment for canines with spinal cord injury: A pilot feasibility study. J. Orthop. Surg. 2021, 29, 23094990211012293. [Google Scholar] [CrossRef]
- Deng, W.S.; Liu, X.Y.; Ma, K.; Liang, B.; Liu, Y.F.; Wang, R.J.; Chen, X.Y.; Zhang, S. Recovery of motor function in rats with complete spinal cord injury following implantation of collagen/silk fibroin scaffold combined with human umbilical cord-mesenchymal stem cells. Rev. Assoc. Med. Bras. 2021, 67, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Tang, J.; Zhao, Y.; Zhang, J.; Xiao, Z.; Chen, B.; Han, G.; Yin, N.; Jiang, X.; Zhao, C.; et al. Long-term clinical observation of patients with acute and chronic complete spinal cord injury after transplantation of NeuroRegen scaffold. Sci. China Life Sci. 2022, 65, 909–926. [Google Scholar] [CrossRef]
- Chen, C.; Xu, H.H.; Liu, X.Y.; Zhang, Y.S.; Zhong, L.; Wang, Y.W.; Xu, L.; Wei, P.; Chen, Y.X.; Liu, P.; et al. 3D printed collagen/silk fibroin scaffolds carrying the secretome of human umbilical mesenchymal stem cells ameliorated neurological dysfunction after spinal cord injury in rats. Regen. Biomater. 2022, 9, rbac014. [Google Scholar] [CrossRef] [PubMed]
- DeBari, M.K.; King, C.I.; Altgold, T.A.; Abbott, R.D. Silk Fibroin as a Green Material. ACS Biomater. Sci. Eng. 2021, 7, 3530–3544. [Google Scholar] [CrossRef] [PubMed]
- Shriver, Z.; Capila, I.; Venkataraman, G.; Sasisekharan, R. Heparin and heparan sulfate: Analyzing structure and microheterogeneity. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 207, pp. 159–176. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhang, X.; Li, X.; Ju, D.; Mao, T.; Lu, Y.; Gu, Y.; Qi, L.; Wang, Q.; Wu, Q.; et al. A decellularized spinal cord extracellular matrix-gel/GelMA hydrogel three-dimensional composite scaffold promotes recovery from spinal cord injury via synergism with human menstrual blood-derived stem cells. J. Mater. Chem. B 2022, 10, 5753–5764. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, H.; Hu, Y.; Huang, Z.; Wang, Y.; Xia, Y.; Chen, X.; Guo, J.; Cheng, H.; Xia, L.; et al. GelMA-MXene hydrogel nerve conduits with microgrooves for spinal cord injury repair. J. Nanobiotechnol. 2022, 20, 460. [Google Scholar] [CrossRef]
- Abdolahi, S.; Aligholi, H.; Khodakaram-Tafti, A.; Khaleghi Ghadiri, M.; Stummer, W.; Gorji, A. Improvement of Rat Spinal Cord Injury Following Lentiviral Vector-Transduced Neural Stem/Progenitor Cells Derived from Human Epileptic Brain Tissue Transplantation with a Self-assembling Peptide Scaffold. Mol. Neurobiol. 2021, 58, 2481–2493. [Google Scholar] [CrossRef]
- Shen, H.; Xu, B.; Yang, C.; Xue, W.; You, Z.; Wu, X.; Ma, D.; Shao, D.; Leong, K.; Dai, J. A DAMP-scavenging, IL-10-releasing hydrogel promotes neural regeneration and motor function recovery after spinal cord injury. Biomaterials 2022, 280, 121279. [Google Scholar] [CrossRef]
- Li, L.; Xiao, B.; Mu, J.; Zhang, Y.; Zhang, C.; Cao, H.; Chen, R.; Patra, H.K.; Yang, B.; Feng, S.; et al. A MnO(2) Nanoparticle-Dotted Hydrogel Promotes Spinal Cord Repair via Regulating Reactive Oxygen Species Microenvironment and Synergizing with Mesenchymal Stem Cells. ACS Nano 2019, 13, 14283–14293. [Google Scholar] [CrossRef]
- Yang, B.; Liang, C.; Chen, D.; Cheng, F.; Zhang, Y.; Wang, S.; Shu, J.; Huang, X.; Wang, J.; Xia, K.; et al. A conductive supramolecular hydrogel creates ideal endogenous niches to promote spinal cord injury repair. Bioact. Mater. 2022, 15, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chu, R.; Ni, N.; Nan, G. The effect of Matrigel as scaffold material for neural stem cell transplantation for treating spinal cord injury. Sci. Rep. 2020, 10, 2576. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yang, H.; Chen, D.; Xie, Y.; Tai, C.; Wang, L.; Wang, P.; Wang, B. Three-dimensional bioprinting sodium alginate/gelatin scaffold combined with neural stem cells and oligodendrocytes markedly promoting nerve regeneration after spinal cord injury. Regen. Biomater. 2022, 9, rbac038. [Google Scholar] [CrossRef] [PubMed]
- Koffler, J.; Zhu, W.; Qu, X.; Platoshyn, O.; Dulin, J.N.; Brock, J.; Graham, L.; Lu, P.; Sakamoto, J.; Marsala, M.; et al. Biomimetic 3D-printed scaffolds for spinal cord injury repair. Nat. Med. 2019, 25, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Li, C.; Chen, Y.; Zhang, L.; Yang, H.; Zhang, Z.; Yue, J.; Gao, W.; Zhu, H.; Pan, H.; et al. LncRNA-GAS5 promotes spinal cord repair and the inhibition of neuronal apoptosis via the transplantation of 3D printed scaffold loaded with induced pluripotent stem cell-derived neural stem cells. Ann. Transl. Med. 2021, 9, 931. [Google Scholar] [CrossRef]
- Liu, X.; Song, S.; Chen, Z.; Gao, C.; Li, Y.; Luo, Y.; Huang, J.; Zhang, Z. Release of O-GlcNAc transferase inhibitor promotes neuronal differentiation of neural stem cells in 3D bioprinted supramolecular hydrogel scaffold for spinal cord injury repair. Acta Biomater. 2022, 151, 148–162. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Subject | Stem Cell Type | Scaffold Material | Outcome |
|---|---|---|---|---|
| Kourgiantaki et al. [129] | C57/BL6 mice | NSPCs | Collagen | Improved axonal elongation, neural regeneration at SCI lesions, enhanced NSPC differentiation, and functional integration of the regenerated cells into the preexisting neural network |
| Liu et al. [131] | Sprague-Dawley rats | NSCs | Collagen | Improved hindlimb motor function, nerve regeneration, and neural cell extension |
| Deng et al. [130] | Sprague-Dawley rats and beagle canines | MSCs | Collagen | Increased motor scores, reduced SCI lesions |
| Deng et al. [130] | Humans | MSCs | Collagen | Emergence of novel nerve fiber growth, improved electrophysiological activity of neurons adjacent to the SCI lesion, increased daily life scores, increased American Spinal Injury Association scores, improved bladder and bowel functioning |
| Tang et al. [109] | Humans | Bone marrow mononuclear cells and MSCs | Collagen | Improved bowel and bladder sensation, improved voluntary walking activity, enhanced finger mobility |
| Liu et al. [128] | Sprague-Dawley rats | NSPCs | Collagen modified with N-cadherin | Increased NSPC recruitment to SCI lesion, improved locomotor activity |
| Chen et al. [135] | Sprague-Dawley rats | MSCs | Collagen modified with silk | Improved nerve fiber regeneration, enhanced remyelination, establishment of novel synaptic connections at the SCI lesion |
| Deng et al. [132] | Beagle canines | MSCs | Collagen modified with heparan sulfate | Improved locomotor activity, improved urodynamic parameters, modulation of cytokines |
| Source | Subject | Stem Cell Type | Scaffold Material | Outcome |
|---|---|---|---|---|
| Wang et al. [144] | Sprague-Dawley rats | NSCs | Matrigel | Slight neural recovery and improved motor function |
| Li et al. [142] | Sprague-Dawley rats | MSCs | Hyaluronic acid hydrogel with manganese dioxide nanoparticles | Enhanced MSC growth and differentiation, restoration of locomotor function |
| Abdolahi et al. [140] | Sprague-Dawley rats | NSCs | PuraMatrix peptide hydrogel | Enhance NSC survival and differentiation, reduced SCI lesion volume, improved neurologic functioning |
| Yang et al. [143] | C57/BL6 mice | NSPCs | Hydrogel enhanced with agarose, gelatin, and polypyrrole | Enhanced NSPC differentiation, reduced SCI lesion volume |
| He et al. [138] | Sprague-Dawley rats | MenSCs | DSCG/GelMA hydrogel | Improved motor function, reduced inflammation, enhanced MenSC differentiation |
| Cai et al. [139] | Sprague-Dawley rats | NSCs | GelMA-MXene hydrogel | Improved motor function, reduced inflammation, enhanced NSC differentiation |
| Shen et al. [141] | C57/BL6 mice | NSCs | IL-10-enhanced hydrogel | Enhanced NSC differentiation, neural regeneration, and axonal regrowth |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hey, G.; Willman, M.; Patel, A.; Goutnik, M.; Willman, J.; Lucke-Wold, B. Stem Cell Scaffolds for the Treatment of Spinal Cord Injury—A Review. Biomechanics 2023, 3, 322-342. https://doi.org/10.3390/biomechanics3030028
Hey G, Willman M, Patel A, Goutnik M, Willman J, Lucke-Wold B. Stem Cell Scaffolds for the Treatment of Spinal Cord Injury—A Review. Biomechanics. 2023; 3(3):322-342. https://doi.org/10.3390/biomechanics3030028
Chicago/Turabian StyleHey, Grace, Matthew Willman, Aashay Patel, Michael Goutnik, Jonathan Willman, and Brandon Lucke-Wold. 2023. "Stem Cell Scaffolds for the Treatment of Spinal Cord Injury—A Review" Biomechanics 3, no. 3: 322-342. https://doi.org/10.3390/biomechanics3030028