
The Large American Liver Fluke (Fascioloides magna): A Survivor’s Journey through a Constantly Changing World

{kind=link}
{kind=link}
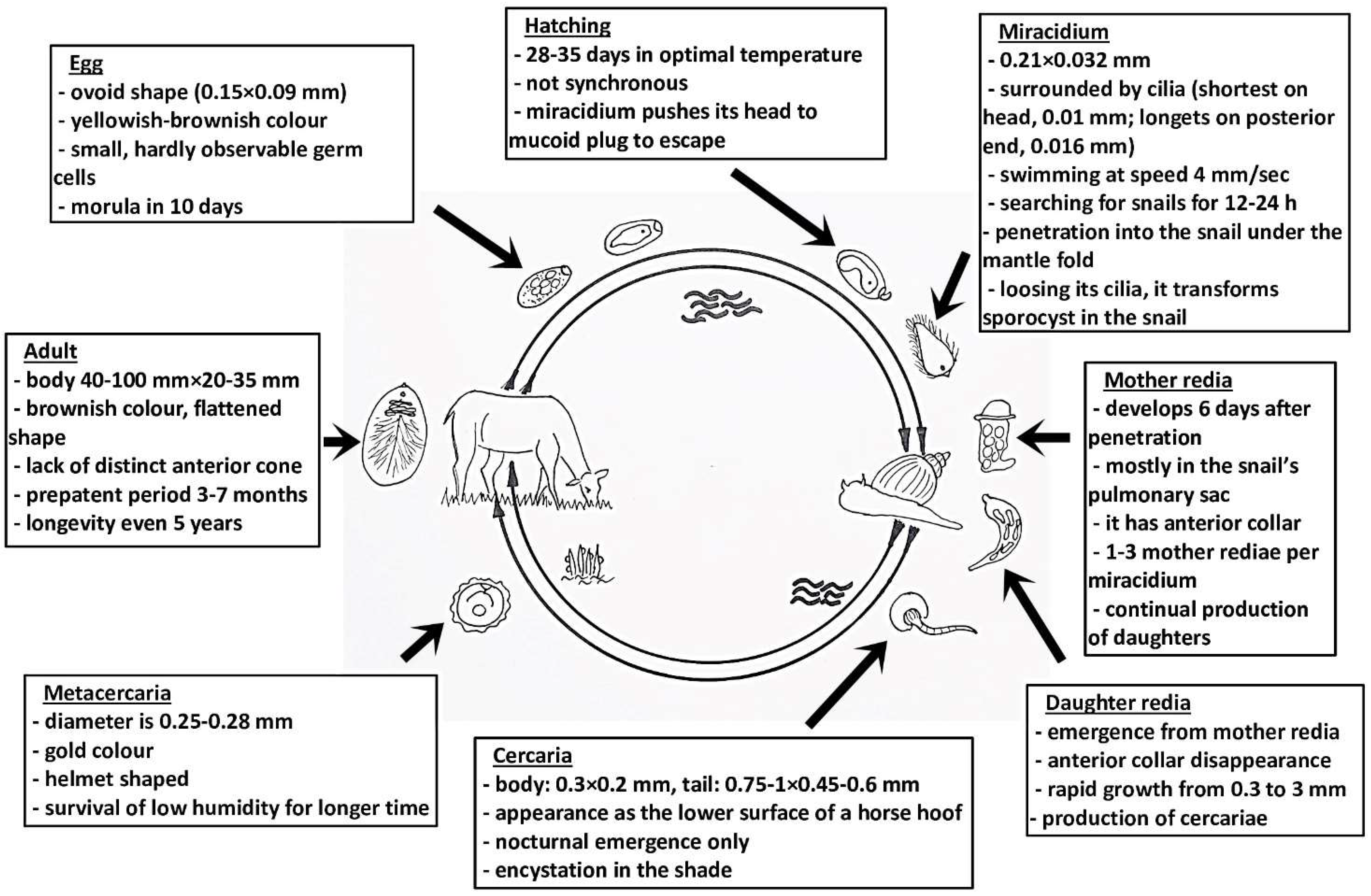{kind=link}
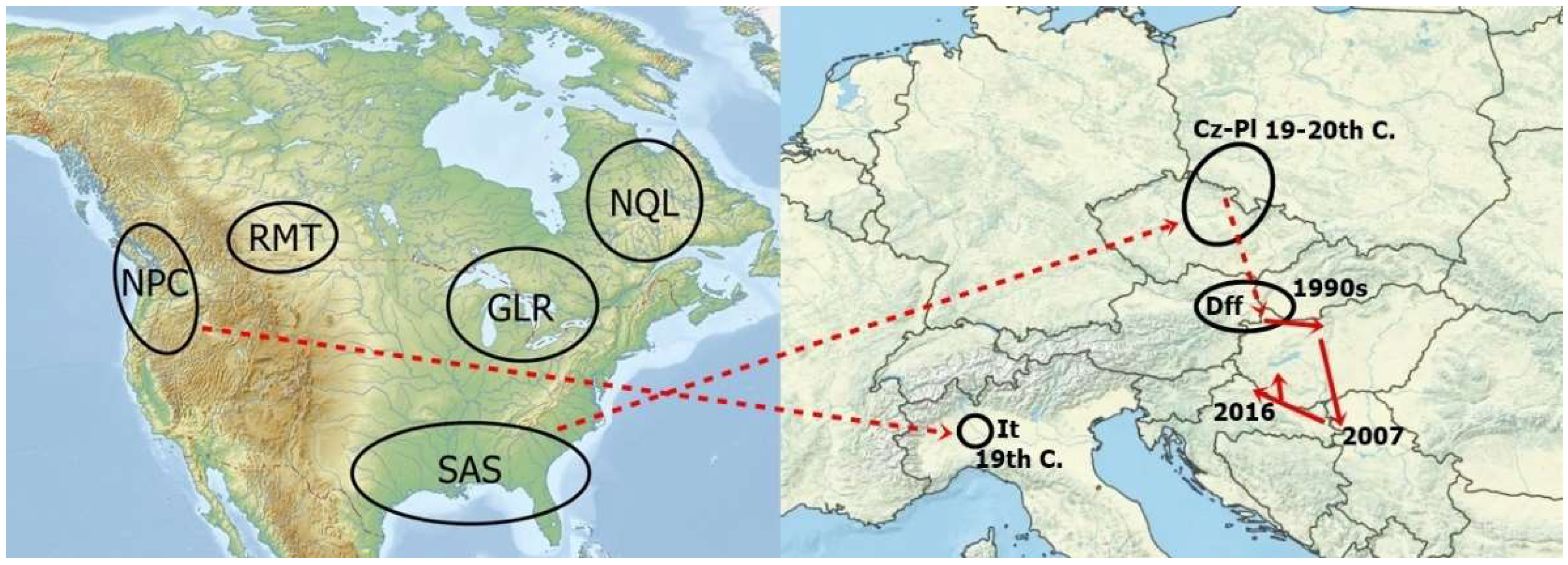{kind=link}
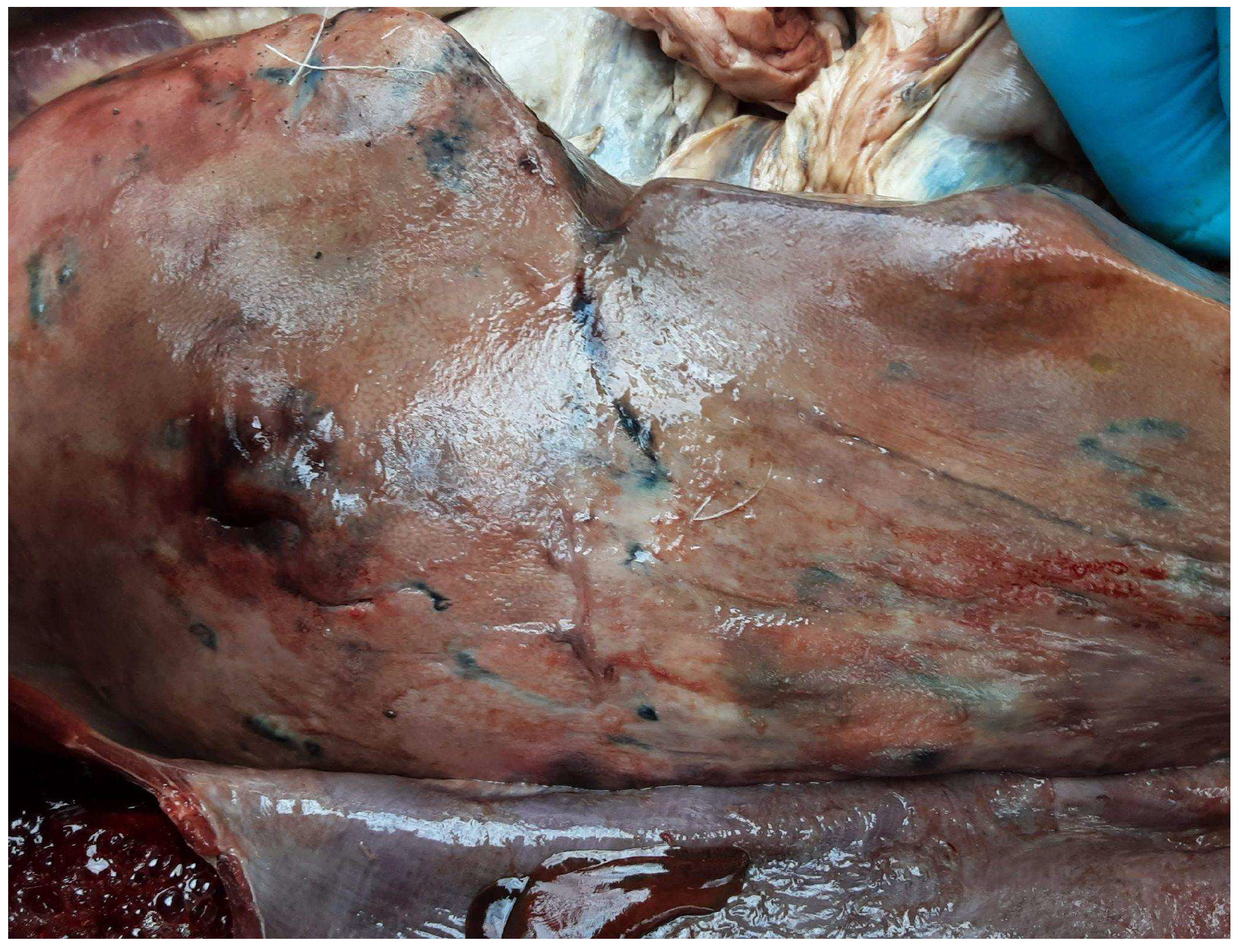{kind=link}
{kind=link}
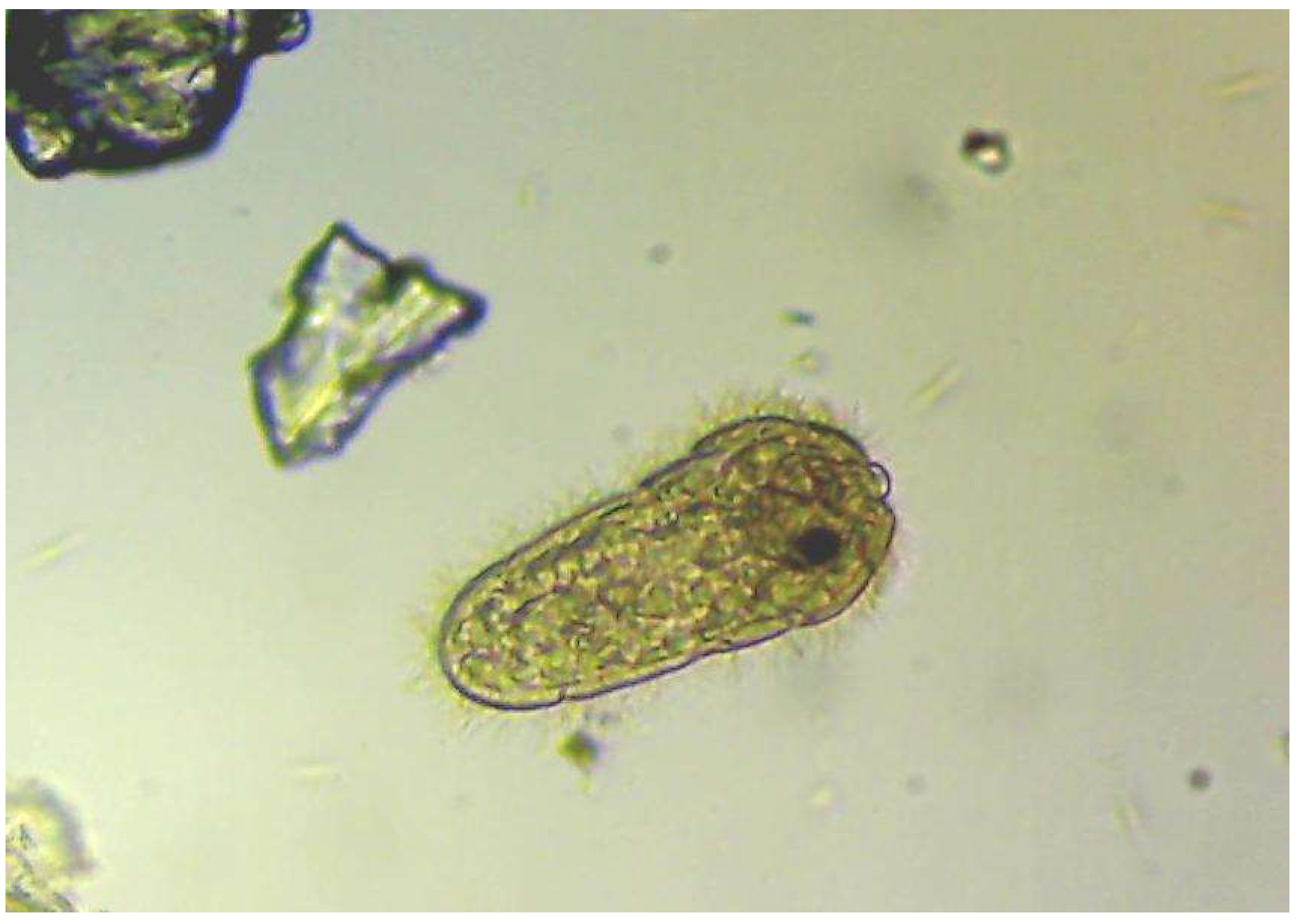{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Evolutionary History of the Large American Liver Fluke and Its Relatives
2.1. The Cretaceous-Paleogene Boundary
2.2. Distribution by the Proboscidea Order
2.3. The Fall of the Proboscideans and the Dawn of the Ruminants
2.4. Switch to the Cervid Host
3. Recent European Distribution
4. Final Hosts of F. magna
4.1. The Special Role of Roe Deer
4.2. Efforts to Control F. magna in Cervid Hosts
5. Intermediate Hosts of F. magna
5.1. Susceptibility of the Intermediate Hosts
5.2. The Course of Infection in Intermediate Hosts
5.3. Self-Fertilisation: A Trade-off between Population Persistence and Population Health
5.4. Environmental Demands of Snail Hosts
5.5. Human-Mediated Distribution of Epidemiological Risk of Snails
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lotfy, W.M.; Brant, S.V.; DeJong, R.J.; Le, T.H.; Demiaszkiewicz, A.; Rajapakse, R.J.; Perera, V.B.; Laursen, J.R.; Loker, E.S. Evolutionary origins, diversification, and biogeography of liver flukes (Digenea, Fasciolidae). Am. J. Trop. Med. Hyg. 2008, 79, 248. [Google Scholar] [CrossRef] [PubMed]
- Kassai, T. Digenea. In Veterinary Helminthology; Butterworth-Heinemann: Oxford, UK, 1999. [Google Scholar]
- Malcicka, M. Life history and biology of Fascioloides magna (Trematoda) and its native and exotic hosts. Ecol. Evol. 2015, 5, 1381–1397. [Google Scholar] [CrossRef]
- Bassi, R. Sulla cachessia ittero-verminosa, o marciaia, causata dal Distomum magnum. J. Med. Vet. Torino 1875, 4, 497–515. [Google Scholar]
- Juhász, A.; Stothard, J.R. The giant liver fluke in Europe: A review of Fascioloides magna within cervids and livestock with considerations on an expanding snail-fluke transmission risk. Adv. Parasit. 2022, 119, 223–257. [Google Scholar]
- Swales, W.E. The life cycle of Fascioloides magna (Bassi, 1875), the large liver fluke of ruminants, in Canada: With observations on the bionomics of the larval stages and the intermediate hosts, pathology of Fascioloidiasis magna, and control measures. Can. J. Res. 1935, 12, 177–215. [Google Scholar] [CrossRef]
- Králová-Hromadová, I.; Juhásová, L.; Bazsalovicsová, E. The Giant Liver Fluke, Fascioloides magna: Past, Present and Future Research; Springer International Publishing: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Marinković, D.; Kukolj, V.; Aleksić-Kovačević, S.; Jovanović, M.; Knežević, M. The role of hepatic myofibroblasts in liver cirrhosis in fallow deer (Dama dama) naturally infected with giant liver fluke (Fascioloides magna). BMC Vet. Res. 2013, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Šimonji, K.; Konjević, D.; Bujanić, M.; Rubić, I.; Farkaš, V.; Beletić, A.; Grbavac, L.; Kuleš, J. Liver proteome alterations in red deer (Cervus elaphus) infected by the giant liver fluke Fascioloides magna. Pathogens 2022, 11, 1503. [Google Scholar] [CrossRef]
- Sommer, M.F.; Drdlicek, J.; Müller, M.; Thelemann, A.; Just, F.T. Fascioloides magna and other liver parasites in cloven-hoofed game from northeastern Bavaria, Germany: Occurrence and pathological findings with special emphasis on red deer (Cervus elaphus). Eur. J. Wildl. Res. 2022, 68, 73. [Google Scholar] [CrossRef]
- Pankrác, J.; Novobilský, A.; Rondelaud, D.; Leontovyč, R.; Syrovátka, V.; Rajský, D.; Horák, P.; Kašný, M. Effect of Fascioloides magna (Digenea) on fecundity, shell height, and survival rate of Pseudosuccinea columella (Lymnaeidae). Parasit. Res. 2016, 115, 3119–3125. [Google Scholar] [CrossRef]
- Rondelaud, D.; Novobilský, A.; Höglund, J.; Kašný, M.; Pankrác, J.; Vignoles, P.; Dreyfuss, G. Growth rate of the intermediate snail host Galba truncatula influences redial development of the trematode Fascioloides magna. J. Helmint. 2014, 88, 427–433. [Google Scholar] [CrossRef]
- Lockyer, A.E.; Jones, C.S.; Noble, L.R.; Rollinson, D. Trematodes and snails: An intimate association. Can. J. Zool. 2004, 82, 251–269. [Google Scholar] [CrossRef]
- Campbell, W.C. Notes on the Egg and Miracidium of Fascioloides magna, (Trematoda). Transac. Am. Microsc. Soc. 1961, 80, 308–319. [Google Scholar] [CrossRef]
- Sattmann, H.; Hörweg, C.; Gaub, L.; Feix, A.S.; Haider, M.; Walochnik, J.; Rabitsch, W.; Prosl, H. Wherefrom and whereabouts of an alien: The American liver fluke Fascioloides magna in Austria: An overview. Wien. Klin. Wochenschr. 2014, 126, 23. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.J.; Park, A.W.; Cressler, C.E.; Dallas, T.; Huang, S.; Mideo, N.; Morales-Castilla, I.; Davies, T.J.; Stephens, P. The ghost of hosts past: Impacts of host extinction on parasite specificity. Philos. Trans. R. Soc. B 2021, 376, 20200351. [Google Scholar] [CrossRef]
- Rogers, R.R.; Rogers, K.A.C.; Bagley, B.C.; Goodin, J.J.; Hartman, J.H.; Thole, J.T.; Zatoń, M. Pushing the record of trematode parasitism of bivalves upstream and back to the Cretaceous. Geology 2018, 46, 431–434. [Google Scholar] [CrossRef]
- Barrios-de Pedro, S.; Osuna, A.; Buscalioni, Á.D. Helminth eggs from early cretaceous faeces. Sci. Rep. UK 2020, 10, 18747. [Google Scholar] [CrossRef]
- Poinar, G.; Boucot, A.J. Evidence of intestinal parasites of dinosaurs. Parasitology 2006, 133, 245–249. [Google Scholar] [CrossRef]
- Kaiho, K.; Oshima, N.; Adachi, K.; Adachi, Y.; Mizukami, T.; Fujibayashi, M.; Saito, R. Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction. Sci. Rep. UK 2016, 6, 28427. [Google Scholar] [CrossRef]
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17084–17093. [Google Scholar] [CrossRef] [PubMed]
- de Moya, R.S.; Allen, J.M.; Sweet, A.D.; Walden, K.K.; Palma, R.L.; Smith, V.S.; Cameron, S.L.; Valim, M.P.; Galloway, T.D.; Weckstein, J.D.; et al. Extensive host-switching of avian feather lice following the Cretaceous-Paleogene mass extinction event. Commun. Biol. 2019, 2, 445. [Google Scholar] [CrossRef]
- Renne, P.R.; Sprain, C.J.; Richards, M.A.; Self, S.; Vanderkluysen, L.; Pande, K. State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact. Science 2015, 350, 76–78. [Google Scholar] [CrossRef]
- Sprain, C.J.; Renne, P.R.; Vanderkluysen, L.; Pande, K.; Self, S.; Mittal, T. The eruptive tempo of Deccan volcanism in relation to the Cretaceous-Paleogene boundary. Science 2019, 363, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Hull, P.M.; Bornemann, A.; Penman, D.E.; Henehan, M.J.; Norris, R.D.; Wilson, P.A.; Blum, P.; Alegret, L.; Batenburg, S.J.; Zachos, J.C.; et al. On impact and volcanism across the Cretaceous-Paleogene boundary. Science 2020, 367, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.K. Constraints on the onset duration of the Paleocene–Eocene Thermal Maximum. Philos. Trans. R. Soc. A 2018, 376, 20170082. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. Adv. Parasit. 2009, 69, 41–146. [Google Scholar]
- Choi, Y.J.; Fontenla, S.; Fischer, P.U.; Le, T.H.; Costábile, A.; Blair, D.; Brindley, P.J.; Tort, J.F.; Cabada, M.M.; Mitreva, M. Adaptive radiation of the flukes of the family Fasciolidae inferred from genome-wide comparisons of key species. Mol. Biol. Evol. 2020, 37, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Correa, A.C.; Escobar, J.S.; Durand, P.; Renaud, F.; David, P.; Jarne, P.; Pointier, J.-P.; Hurtrez-Boussès, S. Bridging gaps in the molecular phylogeny of the Lymnaeidae (Gastropoda: Pulmonata), vectors of fascioliasis. BMC Evol. Biol. 2010, 10, 381. [Google Scholar] [CrossRef]
- Liu, A.G.; Seiffert, E.R.; Simons, E.L. Stable isotope evidence for an amphibious phase in early proboscidean evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 5786–5791. [Google Scholar] [CrossRef] [PubMed]
- Los Huertos, M. Coevolution of biota, geology, and climate. In Ecology and Management of Inland Waters: A Californian Perspective with Global Applications; Elsevier: Amsterdam, Netherlands, 2020. [Google Scholar]
- Lounnas, M.; Correa, A.C.; Vázquez, A.A.; Dia, A.; Escobar, J.S.; Nicot, A.; Arenas, J.; Ayaqui, R.; Dubois, M.P.; Gimenez, T.; et al. Self-fertilization, long-distance flash invasion and biogeography shape the population structure of Pseudosuccinea columella at the worldwide scale. Molec. Ecol. 2017, 26, 887–903. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, T.A.; Georgopoulou, E. Extinction risk is linked to lifestyle in freshwater gastropods. Divers. Distrib. 2021, 27, 2357–2368. [Google Scholar] [CrossRef]
- Palasio, R.G.S.; Chiaravalloti-Neto, F.; Tuan, R. Distribution of genetic diversity of neotropical Biomphalaria (Preston 1910) (Basommatophora: Planorbidae) intermediate hosts for schistosomiasis in Southeast Brazil. Front. Trop. Dis. 2023, 4, 1143186. [Google Scholar] [CrossRef]
- Yu, T.; Neubauer, T.A.; Jochum, A. First freshwater gastropod preserved in amber suggests long-distance dispersal during the Cretaceous Period. Geol. Mag. 2021, 158, 1327–1334. [Google Scholar] [CrossRef]
- Juhász, A.; Majoros, G. A proposed ectochory of Galba truncatula snails between wallow sites enhances transmission of Fascioloides magna at Gemenc, in Hungary. Int. J. Parasitol. Parasites Wildl. 2023, 21, 2019–2223. [Google Scholar] [CrossRef] [PubMed]
- Hurtrez-Boussès, S.; Hurtrez, J.E.; Turpin, H.; Durand, C.; Durand, P.; De Meeüs, T.; Meunier, C.; Renaud, F. Hydrographic network structure and population genetic differentiation in a vector of fasciolosis, Galba truncatula. Infect. Genet. Evol. 2010, 10, 178–183. [Google Scholar] [CrossRef] [PubMed]
- McCabe, D.J. Rivers and streams: Life in flowing water. Nat. Educ. Knowl. 2010, 3, 19. [Google Scholar]
- Sturm, R. Aquatic molluscs in high mountain lakes of the Eastern Alps (Austria): Species-environment relationships and specific colonization behaviour. Chin. J. Oceanol. Limnol. 2012, 30, 59. [Google Scholar] [CrossRef]
- Bargues, M.D.; Artigas, P.; Khoubbane, M.; Ortiz, P.; Naquira, C.; Mas-Coma, S. Molecular characterisation of Galba truncatula, Lymnaea neotropica and L. schirazensis from Cajamarca, Peru and their potential role in transmission of human and animal fascioliasis. Parasites Vectors 2012, 5, 174. [Google Scholar] [CrossRef]
- Alba, A.; Vázquez, A.A.; Sánchez, J.; Lounnas, M.; Pointier, J.P.; Hurtrez-Boussès, S.; Gourbal, B. Patterns of distribution, population genetics and ecological requirements of field-occurring resistant and susceptible Pseudosuccinea columella snails to Fasciola hepatica in Cuba. Sci. Rep. 2019, 9, 14359. [Google Scholar] [CrossRef]
- Ngcamphalala, P.I.; Malatji, M.P.; Mukaratirwa, S. Geography and ecology of invasive Pseudosuccinea columella (Gastropoda: Lymnaeidae) and implications in the transmission of Fasciola species (Digenea: Fasciolidae)—A review. J. Helminthol. 2022, 96, e1. [Google Scholar] [CrossRef]
- Grabner, D.S.; Mohamed, F.A.; Nachev, M.; Meabed, E.M.; Sabry, A.H.A.; Sures, B. Invasion biology meets parasitology: A case study of parasite spill-back with Egyptian Fasciola gigantica in the invasive snail Pseudosuccinea columella. PLoS ONE 2014, 9, e88537. [Google Scholar] [CrossRef]
- Saarinen, J. The palaeontology of browsing and grazing. In The Ecology of Browsing and Grazing II, 1st ed.; Gordon, I.J., Prins, H.H.T., Eds.; Springer Nature: Cham, Switzerland, 2019. [Google Scholar]
- Gheerbrant, E. Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. Proc. Natl. Acad. Sci. USA 2009, 106, 10717–10721. [Google Scholar] [CrossRef] [PubMed]
- Baleka, S.; Varela, L.; Tambusso, P.S.; Paijmans, J.L.; Mothé, D.; Stafford, T.W.; Farina, R.A.; Hofreiter, M. Revisiting proboscidean phylogeny and evolution through total evidence and palaeogenetic analyses including Notiomastodon ancient DNA. iScience 2022, 25, 103559. [Google Scholar] [CrossRef]
- Tabuce, R.; Sarr, R.; Adnet, S.; Lebrun, R.; Lihoreau, F.; Martin, J.E.; Sambou, B.; Thiam, M.; Hautier, L. Filling a gap in the proboscidean fossil record: A new genus from the Lutetian of Senegal. J. Paleontol. 2020, 94, 580–588. [Google Scholar] [CrossRef]
- Cantalapiedra, J.L.; Sanisidro, O.; Zhang, H.; Alberdi, M.T.; Prado, J.L.; Blanco, F.; Saarinen, J. The rise and fall of proboscidean ecological diversity. Nat. Ecol. Evol. 2021, 5, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Pham, K.L.T.; Doan, H.T.T.; Le, T.X.; Nguyen, K.T.; Lawton, S.P. Description and phylogenetic analyses of ribosomal transcription units from species of Fasciolidae (Platyhelminthes: Digenea). J. Helminthol. 2020, 94, e136. [Google Scholar] [CrossRef] [PubMed]
- Heneberg, P. Phylogenetic data suggest the reclassification of Fasciola jacksoni (Digenea: Fasciolidae) as Fascioloides jacksoni comb. nov. Parasitol. Res. 2013, 112, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, R.P.V.J.; Pham, K.L.T.; Karunathilake, K.K.; Lawton, S.P.; Le, T.H. Characterization and phylogenetic properties of the complete mitochondrial genome of Fascioloides jacksoni (syn. Fasciola jacksoni) support the suggested intergeneric change from Fasciola to Fascioloides (Platyhelminthes: Trematoda: Plagiorchiida). Infect. Genet. Evol. 2020, 82, 104281. [Google Scholar]
- Reblánová, M.; Špakulová, M.; Orosová, M.; Bazsalovicsová, E.; Rajský, D. A description of karyotype of the giant liver fluke Fascioloides magna (Trematoda, Platyhelminthes) and a review of Fasciolidae cytogenetics. Helminthologia 2010, 47, 69–75. [Google Scholar] [CrossRef]
- Bargues, M.D.; Halajian, A.; Artigas, P.; Luus-Powell, W.J.; Valero, M.A.; Mas-Coma, S. Paleobiogeographical origins of Fasciola hepatica and F. gigantica in light of new DNA sequence characteristics of F. nyanzae from hippopotamus. Front. Vet. Sci. 2022, 9, 990872. [Google Scholar] [CrossRef]
- Mennecart, B.; Dziomber, L.; Aiglstorfer, M.; Bibi, F.; DeMiguel, D.; Fujita, M.; Kubo, M.O.; Laurens, F.; Meng, J.; Métais, G.; et al. Ruminant inner ear shape records 35 million years of neutral evolution. Nat. Commun. 2022, 13, 7222. [Google Scholar] [CrossRef]
- Codron, D. Evolution of large mammal herbivores in savannas. In Savanna Woody Plants and Large Herbivores, 1st ed.; Scogings, P.F., Sankaran, M., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2019. [Google Scholar]
- Sheil, D. Dangerous giants?—Large herbivores, forest feedbacks and climate tipping points. Russ. J. Ecosyst. Ecol. 2020, 5, 1–33. [Google Scholar] [CrossRef]
- Ao, H.; Rohling, E.J.; Zhang, R.; Roberts, A.P.; Holbourn, A.E.; Ladant, J.B.; Dupont-Nivet, G.; Kuhnt, W.; Zhang, P.; Wu, F.; et al. Global warming-induced Asian hydrological climate transition across the Miocene–Pliocene boundary. Nat. Commun. 2021, 12, 6935. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Crespo, V.A.; Prado, J.L.; Alberdi, M.T.; Arroyo-Cabrales, J.; Johnson, E. Feeding ecology of the gomphotheres (Proboscidea, Gomphotheriidae) of America. Quat. Sci. Rev. 2020, 229, 106126. [Google Scholar] [CrossRef]
- Ameen, M.; Khan, A.M.; Ahmad, R.M.; Ijaz, M.U.; Imran, M. Tooth marker of ecological abnormality: The interpretation of stress in extinct mega herbivores (proboscideans) of the Siwaliks of Pakistan. Ecol. Evol. 2022, 12, e9432. [Google Scholar] [CrossRef]
- Gilbert, C.; Ropiquet, A.; Hassanin, A. Mitochondrial and nuclear phylogenies of Cervidae (Mammalia, Ruminantia): Systematics, morphology, and biogeography. Mol. Phylogenet. Evol. 2006, 40, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Polziehn, R.O.; Strobeck, C. Phylogeny of wapiti, red deer, sika deer, and other North American cervids as determined from mitochondrial DNA. Mol. Phylogenet. Evol. 1998, 10, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Pybus, M.J. Liver flukes. In Parasitic Diseases of Wild Mammals; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa State Press: Ames, IA, USA, 2021; pp. 121–149. [Google Scholar]
- Parker, A.; David, A.A. Genetic characterization of the giant liver fluke, Fascioloides magna (Platyhelminthes: Fasciolidae) from the Adirondack Region of Northern New York. Acta Parasitol. 2021, 66, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Králová-Hromadová, I.; Bazsalovicsová, E.; Štefka, J.; Špakulová, M.; Vávrová, S.; Szemes, T.; Tkach, V.; Trudgett, A.; Pybus, M. Multiple origins of European populations of the giant liver fluke Fascioloides magna (Trematoda: Fasciolidae), a liver parasite of ruminants. Int. J. Parasitol. 2011, 41, 373–383. [Google Scholar] [CrossRef]
- Engelstädter, J.; Fortuna, N.Z. The dynamics of preferential host switching: Host phylogeny as a key predictor of parasite distribution. Evolution 2019, 73, 1330–1340. [Google Scholar] [CrossRef]
- Demiaszkiewicz, A.W.; Kuligowska, I.; Pyziel, A.M.; Lachowicz, J.; Kowalczyk, R. Extension of occurrence area of the American fluke Fascioloides magna in south-western Poland. Ann. Parasitol. 2015, 61, 93–96. [Google Scholar]
- Erhardová, B. Fascioloides magna in Europe. Helminthologia 1961, 3, 91–106. [Google Scholar]
- Erhardová-Kotrlová, B. The Occurence of Fascioloides magna (Bassi, 1875) in Czechoslovakia; Czechoslovak Academy of Science: Prague, Czech Republic, 1971; p. 155. [Google Scholar]
- Rajský, D.; Patus, A.; Bukovjan, K. Prvýnález The first finding of Fascioloides magna Bassi, 1875 (in Slovakian: Fascioloides magna Bassi, 1875 na Slovensku. Slov. Vet. Čas. 1994, 19, 29–30. [Google Scholar]
- Majoros, G.; Sztojkov, V. Appearance of the large American liver fluke Fascioloides magna (Bassi, 1875) (Trematoda: Fasciolata) in Hungary. Parasitol. Hung. 1994, 27, 27–38. [Google Scholar]
- Sztojkov, V.; Majoros, G.; Kámán, K. Szarvasokban élő nagy amerikai májmétely (Fascioloides magna) megjelenése Magyarországon (in Hungarian). Mag. Allat. Lap 1995, 50, 157–159. [Google Scholar]
- Houszka, M.; Piekarska, J.; Podkowik, M.; Gorczykowski, M.; Bania, J. Morphology and molecular study of Fascioloides magna–a growing threat to cervids (Cervidae) in Poland. J. Vet. Res. 2016, 60, 435–439. [Google Scholar] [CrossRef]
- Juhásová, L.; Králová-Hromadová, I.; Zeleňáková, M.; Blišťan, P.; Bazsalovicsová, E. Transmission risk assessment of invasive fluke Fascioloides magna using GIS-modelling and multicriteria analysis methods. Helminthologia 2017, 54, 119–131. [Google Scholar] [CrossRef]
- Marinculić, A.; Džakula, N.; Janicki, Z.; Hardy, Z.; Lučinger, S.; Živičnjak, T. Appearance of American liver fluke (Fascioloides magna, Bassi, 1875) in Croatia. Vet. Arh. 2002, 72, 319–325. [Google Scholar]
- Slavica, A.; Florijančić, T.; Janicki, Z.; Konjević, D.; Severin, K.; Marinculić, A.; Pintur, K. Treatment of fascioloidosis (Fascioloides magna, Bassi, 1875) in free ranging and captive red deer (Cervus elaphus L.) at eastern Croatia. Vet. Arh. 2006, 76, S9–S18. [Google Scholar]
- Konjević, D.; Janicki, Z.; Calmels, P.; Jan, D.S.; Marinculić, A.; Šimunović, M.; Pavlak, M.; Krapinec, K.; Poljak, Z. Evaluation of factors affecting the efficacy of treatment against Fascioloides magna in wild red deer population. Vet. Ital. 2018, 54, 33–39. [Google Scholar]
- Trailović, S.M.; Marinković, D.; Kulišić, Z. Diagnosis and therapy of liver fluke (Fascioloides magna) infection in fallow deer (Dama dama) in Serbia. J. Wildl. Dis. 2016, 52, 319–326. [Google Scholar] [CrossRef]
- Mirčeta, J.; Pelić, M.; Božić, B.; Petrović, J.; Urošević, M.; Stankov, B.; Bugarski, D. Prevalence of the giant liver fluke (Fascioloides magna, Bassi, 1875) in red deer (Cervus elaphus) in the region of floodplain forests of northern Serbia. Arch. Vet. Med. 2018, 11, 17–26. [Google Scholar] [CrossRef]
- Salomon, S. Feucioloides magna Lei deutschem Rotwild (in German). Berl. Tierarztl. Wschr. 1932, 48, 627–628. [Google Scholar]
- Plötz, C.; Rehbein, S.; Bamler, H.; Reindl, H.; Pfister, K.; Scheuerle, M.C. Fascioloides magna–epizootiology in a deer farm in Germany. Berl. Munch. Tierärzt. Wochensch. 2015, 128, 111–182. [Google Scholar]
- Pfeiffer, H. Fascioloides magna; first report in Austria (in German). Wiener Tierarztl. Monatsschr. 1983, 70, 168–170. [Google Scholar]
- Konjević, D.; Bujanić, M.; Beck, A.; Beck, R.; Martinković, F.; Janicki, Z. First record of chronic Fascioloides magna infection in roe deer (Capreolus capreolus). Int. J. Parasitol. Parasites Wildl. 2021, 15, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Halász, T.; Tari, T.; Nagy, E.; Nagy, G.; Csivincsik, Á. Hatchability of Fascioloides magna eggs in cervids. Pathogens 2023, 12, 741. [Google Scholar] [CrossRef] [PubMed]
- Králová-Hromadová, I.; Špakulová, M.; Horáčková, E.; Turčeková, L.; Novobilský, A.; Beck, R.; Koudela, B.; Marinculic, A.; Rajský, D.; Pybus, M. Sequence analysis of ribosomal and mitochondrial genes of the giant liver fluke Fascioloides magna (Trematoda: Fasciolidae): Intraspecific variation and differentiation from Fasciola hepatica. J. Parasitol. 2008, 94, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Bazsalovicsová, E.; Králová-Hromadová, I.; Štefka, J.; Minárik, G.; Bokorová, S.; Pybus, M. Genetic interrelationships of North American populations of giant liver fluke Fascioloides magna. Parasites Vectors 2015, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, S.L.; Stromberg, B.E.; Hayden, D.W.; Averbeck, G.A.; Wilson, J.H. Natural infection of a horse with Fascioloides magna. J. Vet. Diagn. Investig. 2005, 17, 382–385. [Google Scholar] [CrossRef]
- Migaki, G.; Zinter, D.E.; Garner, F.M. Fascioloides magna in the pig; 3 cases. Am. J. Vet. Res. 1971, 32, 1417–1421. [Google Scholar]
- Erhardová-Kotrlá, B.; Blazer, K. Artificial infestation caused by the fluke Fascioloides magna. Acta Vet. Brno 1970, 39, 287–295. [Google Scholar]
- Konjević, D.; Bujanić, M.; Erman, V.; Kurilj, A.G.; Živičnjak, T.; Severin, K.; Tomić, S.; Martinković, F. New data on wild boar (Sus scrofa L.) a dead-end host for large American liver fluke (Fascioloides magna). Helminthologia 2017, 54, 77–80. [Google Scholar] [CrossRef]
- Leontovyč, R.; Košťáková, M.; Siegelová, V.; Melounová, K.; Pankrác, J.; Vrbová, K.; Horák, P.; Kašný, M. Highland cattle and Radix labiata, the hosts of Fascioloides magna. BMC Vet. Res. 2014, 10, 41. [Google Scholar] [CrossRef]
- Demiaszkiewicz, A.W.; Kowalczyk, R.; Filip, K.J.; Pyziel, A.M. Fascioloides magna: A parasite of roe deer in Bory Zielonogórskie (in Polish). Med. Weter. 2018, 74, 257–260. [Google Scholar] [CrossRef]
- Filip-Hutsch, K.; Pyziel-Serafin, A.M.; Hutsch, T.; Bulak, K.; Czopowicz, M.; Merta, D.; Kobielski, J.; Demiaszkiewicz, A.W. The occurrence of Fascioloides magna (Bassi, 1875) in the wild cervid population in the Lower Silesian Wilderness–epidemiological and pathological aspects. J. Vet. Res. 2022, 66, 381–387. [Google Scholar] [CrossRef]
- Shury, T.K.; Pybus, M.J.; Nation, N.; Cool, N.L.; Rettie, W.J. Fascioloides magna in moose (Alces alces) from Elk Island National Park, Alberta. Vet. Pathol. 2019, 56, 476–485. [Google Scholar] [PubMed]
- Živković, D.; John, S.; Verin, M.; Stephan, W.; Tellier, A. Neutral genomic signatures of host-parasite coevolution. BMC Evol. Biol. 2019, 19, 230. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, L.J.; Ashby, B. Coevolutionary theory of hosts and parasites. J. Evol. Biol. 2022, 35, 205–224. [Google Scholar] [CrossRef]
- Mestorino, N.; Formentini, E.A.; Lucas, M.F.; Fernandez, C.; Modamio, P.; Hernández, E.M.; Errecalde, J.O. Pharmacokinetic disposition of triclabendazole in cattle and sheep; discrimination of the order and the rate of the absorption process of its active metabolite triclabendazole sulfoxide. Vet. Res. Commun. 2008, 32, 21–33. [Google Scholar] [CrossRef]
- Haider, M.; Hörweg, C.; Liesinger, K.; Sattmann, H.; Walochnik, J. Recovery of Fascioloides magna (Digenea) population in spite of treatment programme? Screening of Galba truncatula (Gastropoda, Lymnaeidae) from Lower Austria. Vet. Parasitol. 2012, 187, 445–451. [Google Scholar] [CrossRef]
- Saadi, A.J.; Davison, A.; Wade, C.M. Molecular phylogeny of freshwater snails and limpets (Panpulmonata: Hygrophila). Zool. J. Linn. Soc. 2020, 190, 518–531. [Google Scholar] [CrossRef]
- Hörweg, C.; Prosl, H.; Wille-Piazzai, W.; Joachim, A.; Sattmann, H. Prevalence of Fascioloides magna in Galba truncatula in the Danube backwater area east of Vienna, Austria. Wien. Tierarztl. Monat. 2011, 98, 261–267. [Google Scholar]
- Schniebs, K.; Peter, G.; Vinarski, M.V.; Hundsdoerfer, A.K. Intraspecific morphological and genetic variability in the European freshwater snail Radix labiata (Rossmaessler, 1835)(Gastropoda: Basommatophora: Lymnaeidae). Contrib. Zool. 2013, 82, 55–68. [Google Scholar] [CrossRef]
- Salgado, S.Q.; Soriano, J.L.; Glöer, P. First report of Radix labiata (Rossmässler, 1835) (Gastropoda: Lymnaeidae) in Aragon (NE Spain). J. Conchol 2016, 40, 657–678. [Google Scholar]
- Garbar, А.; Komіushіn, А.V. Karyotypes of European species of Radix (Gastropoda: Pulmonata: Lymnaeidae) and their relevance to species distinction in the genus. Malacologia 2003, 45, 141–148. [Google Scholar]
- Alda, P.; Lounnas, M.; Vázquez, A.A.; Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon, R.T., Jr.; Jarne, P.; Loker, E.S.; Muñiz Pareja, F.C.; et al. A new multiplex PCR assay to distinguish among three cryptic Galba species, intermediate hosts of Fasciola hepatica. Vet. Parasitol. 2018, 251, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Bargues, M.D.; Mas-Coma, S. Phylogenetic analysis of lymnaeid snails based on 18S rDNA sequences. Mol. Biol. Evol. 1997, 14, 569–577. [Google Scholar] [CrossRef]
- Davis, C.N.; Tyson, F.; Cutress, D.; Davies, E.; Jones, D.L.; Brophy, P.M.; Prescott, A.; Rose, M.T.; Williams, M.; Williams, H.W.; et al. Rapid detection of Galba truncatula in water sources on pasture-land using loop-mediated isothermal amplification for control of trematode infections. Parasites Vectors 2020, 13, 496. [Google Scholar] [CrossRef]
- Caron, Y.; Righi, S.; Lempereur, L.; Saegerman, C.; Losson, B. An optimized DNA extraction and multiplex PCR for the detection of Fasciola sp. in lymnaeid snails. Vet. Parasitol. 2011, 178, 93–99. [Google Scholar] [CrossRef]
- Caron, Y.; Lasri, S.; Losson, B. Fasciola hepatica: An assessment on the vectorial capacity of Radix labiata and R. balthica commonly found in Belgium. Vet. Parasitol. 2007, 149, 95–103. [Google Scholar] [CrossRef]
- Novobilský, A.; Kašný, M.; Beran, L.; Rondelaud, D.; Höglund, J. Lymnaea palustris and Lymnaea fuscus are potential but uncommon intermediate hosts of Fasciola hepatica in Sweden. Parasites Vectors 2013, 6, 251. [Google Scholar] [CrossRef]
- Kasny, M.; Beran, L.; Siegelova, V.; Siegel, T.; Leontovyc, R.; Berankova, K.; Pankrac, J.; Kostakova, M.; Horak, P. Geographical distribution of the giant liver fluke (Fascioloides magna) in the Czech Republic and potential risk of its further spread. Vet. Med. (Praha) 2012, 57, 101–109. [Google Scholar] [CrossRef]
- Sindou, P.; Rondelaud, D.; Barthe, D. Fasciola hepatica L.: Comparative studies of tissue lesions in seven lymnaeid species submitted to individual monomiracidial exposures at hatching. Bull. de la Soc. Zool. de France 1990, 115, 331–340. [Google Scholar]
- Georgieva, K.; Georgieva, L.; Mizinska-Boevska, Y.; Stoitsova, S.R. Study of surface carbohydrates in Galba truncatula tissues before and after infection with Fasciola hepatica. Memórias Do Inst. Oswaldo Cruz 2016, 111, 475–483. [Google Scholar] [CrossRef]
- Krull, W.H. The intermediate hosts of Fasciola hepatica and Fascioloides magna in the United States. N. Am. Vet. 1934, 15, 13–17. [Google Scholar]
- Correa, A.C.; De Meeûs, T.; Dreyfuss, G.; Rondelaud, D.; Hurtrez-Boussès, S. Galba truncatula and Fasciola hepatica: Genetic costructures and interactions with intermediate host dispersal. Infect. Genet. Evol. 2017, 55, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, E.; Trouve, S.; Facon, B.; Degen, L.; Goudet, J. High quantitative and no molecular differentiation of a freshwater snail (Galba truncatula) between temporary and permanent water habitats. Mol. Ecol. 2007, 16, 3484–3496. [Google Scholar] [CrossRef]
- Alda, P.; Lounnas, M.; Vázquez, A.A.; Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon, R.T., Jr.; González Ramírez, L.C.; Loker, E.S.; Muzzio-Aroca, J.; et al. Systematics and geographical distribution of Galba species, a group of cryptic and worldwide freshwater snails. Mol. Phylogenetics Evolut. 2021, 157, 107035. [Google Scholar] [CrossRef]
- Ebbs, E.T.; Loker, E.S.; Brant, S.V. Phylogeography and genetics of the globally invasive snail Physa acuta Draparnaud 1805, and its potential to serve as an intermediate host to larval digenetic trematodes. BMC Evol. Biol. 2018, 18, 103. [Google Scholar] [CrossRef]
- Kengne-Fokam, A.C.; Nana-Djeunga, H.C.; Djuikwo-Teukeng, F.F.; Njiokou, F. Analysis of mating system, fecundity, hatching and survival rates in two Schistosoma mansoni intermediate hosts (Biomphalaria pfeifferi and Biomphalaria camerunensis) in Cameroon. Parasites Vectors 2016, 9, 10. [Google Scholar] [CrossRef]
- Burgarella, C.; Gayral, P.; Ballenghien, M.; Bernard, A.; David, P.; Jarne, P.; Correa, A.; Hurtrez-Bousses, S.; Escobar, J.; Galtier, N.; et al. Molecular evolution of freshwater snails with contrasting mating systems. Mol. Biol. Evol. 2015, 32, 2403–2416. [Google Scholar] [CrossRef]
- Vázquez, A.A.; Alba, A.; Alda, P.; Vittecoq, M.; Hurtrez-Boussès, S. On the arrival of fasciolosis in the Americas. Trends Parasitol. 2022, 38, 195–204. [Google Scholar] [CrossRef]
- Campbell, G.; Noble, L.R.; Rollinson, D.; Southgate, V.R.; Webster, J.P.; Jones, C.S. Low genetic diversity in a snail intermediate host (Biomphalaria pfeifferi Krass, 1848) and schistosomiasis transmission in the Senegal River Basin. Mol. Ecol. 2010, 19, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Soldánová, M.; Selbach, C.; Sures, B.; Kostadinova, A.; Pérez-del-Olmo, A. Larval trematode communities in Radix auricularia and Lymnaea stagnalis in a reservoir system of the Ruhr River. Parasites Vectors 2010, 3, 56. [Google Scholar] [CrossRef] [PubMed]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10, 2694. [Google Scholar] [CrossRef]
- Roldán, C.; Begovoeva, M.; López-Olvera, J.R.; Velarde, R.; Cabezón, Ó.; Molinar Min, A.R.; Pizzato, F.; Pasquetti, M.; Fernández Aguilar, X.; Mentaberre, G.; et al. Endemic occurrence of Fasciola hepatica in an alpine ecosystem, Pyrenees, Northeastern Spain. Transbound.Emerg. Dis. 2021, 68, 2589–2594. [Google Scholar] [CrossRef]
- Skowrońska-Ochmann, K.; Cuber, P.; Lewin, I. The first record and occurrence of Stagnicola turricula (Held, 1836) (Gastropoda: Pulmonata: Lymnaeidae) in Upper Silesia (Southern Poland) in relation to different environmental factors. Zoo. Anz. J. Comp. Zool. 2012, 251, 357–363. [Google Scholar] [CrossRef]
- Outa, J.O.; Sattmann, H.; Köhsler, M.; Walochnik, J.; Jirsa, F. Diversity of digenean trematode larvae in snails from Lake Victoria, Kenya: First reports and bioindicative aspects. Acta Trop. 2020, 206, 105437. [Google Scholar] [CrossRef] [PubMed]
- Sokolow, S.H.; Jones, I.J.; Jocque, M.; La, D.; Cords, O.; Knight, A.; Lund, A.; Wood, C.L.; Lafferty, K.D.; Hoover, C.M.; et al. Nearly 400 million people are at higher risk of schistosomiasis because dams block the migration of snail-eating river prawns. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160127. [Google Scholar] [CrossRef]
- Barakat, R.M. Epidemiology of schistosomiasis in Egypt: Travel through time. J. Adv. Res. 2013, 4, 425–432. [Google Scholar] [CrossRef]
- Hrivnák, R.; Medvecká, J.; Baláži, P.; Bubíková, K.; Oťaheľová, H.; Svitok, M. Alien aquatic plants in Slovakia over 130 years: Historical overview, current distribution and future perspectives. NeoBiota 2019, 49, 37–56. [Google Scholar] [CrossRef]
- Garcia-Murillo, P. Hydrocharis laevigata in Europe. Plants 2023, 12, 701. [Google Scholar] [CrossRef] [PubMed]
- Abe, E.M.; Guan, W.; Guo, Y.H.; Kassegne, K.; Qin, Z.Q.; Xu, J.; CHen, J.-H.; Ekpo, U.F.; Li, S.-Z.; Zhou, X.N. Differentiating snail intermediate hosts of Schistosoma spp. using molecular approaches: Fundamental to successful integrated control mechanism in Africa. Infect. Dis. Poverty 2018, 7, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.L. Impact of invasive water hyacinth (Eichhornia crassipes) on snail hosts of schistosomiasis in Lake Victoria, East Africa. EcoHealth 2005, 2, 81–86. [Google Scholar] [CrossRef]
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Preston, D.L.; Crone, E.R.; Miller-ter Kuile, A.; Lewis, C.D.; Sauer, E.L.; Trovillion, D.C. Non-native freshwater snails: A global synthesis of invasion status, mechanisms of introduction, and interactions with natural enemies. Freshw. Biol. 2022, 67, 227–239. [Google Scholar] [CrossRef]
- Vignoles, P.; Dreyfuss, G.; Rondelaud, D. Consequences of invasion by Pseudosuccinea columella on the dynamics of native lymnaeids living on the acid soils of central France. Molluscan Res. 2018, 38, 287–295. [Google Scholar] [CrossRef]
- Schols, R.; Carolus, H.; Hammoud, C.; Muzarabani, K.C.; Barson, M.; Huyse, T. Invasive snails, parasite spillback, and potential parasite spillover drive parasitic diseases of Hippopotamus amphibius in artificial lakes of Zimbabwe. BMC Biol. 2021, 19, 160. [Google Scholar] [CrossRef]
- Pointier, J.P.; Coustau, C.; Rondelaud, D.; Theron, A. Pseudosuccinea columella (Say 1817) (Gastropoda, Lymnaeidae), snail host of Fasciola hepatica: First record for France in the wild. Parasitol. Res. 2007, 101, 1389–1392. [Google Scholar] [CrossRef]
- Varga, A.; Lőkkös, A. Mollusc fauna of the Lake Hévíz (in Hungarian). Soosiana 2021, 35, 3–18. [Google Scholar]
- Njiokou, F.; Mouafo, J.B.; Teukeng, F.; Njine, T.; Ekobo, A.S.; Jarne, P. The influence of self-fertilization and pairing on life-history traits in the freshwater snail Bulinus forskalii (Gastropoda, Planorbidae). Acta Trop. 2000, 76, 159–167. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Vignoles, P.; Rondelaud, D. Relationships between the distribution of Galba truncatula (Gastropoda: Lymnaeidae) climatic conditions and the altitude of municipalities in Haute Vienne (France). Ann. Limnol. 2018, 54, 19–28. [Google Scholar] [CrossRef]
- Crone, E.R.; Sauer, E.L.; Preston, D.L. Non-native fish facilitate non-native snails and alter food web structure in experimental pond communities. Funct. Ecol. 2023, 37, 947–958. [Google Scholar] [CrossRef]
- Beesley, N.J.; Williams, D.J.; Paterson, S.; Hodgkinson, J. Fasciola hepatica demonstrates high levels of genetic diversity, a lack of population structure and high gene flow: Possible implications for drug resistance. Int. J. Parasitol. 2017, 47, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.C.; Kamath, P.L.; Van Heerden, H.; Huang, Y.H.; Barandongo, Z.R.; Bruce, S.A.; Kausrud, K. The roles of environmental variation and parasite survival in virulence–transmission relationships. R. Soc. Open Sci. 2021, 8, 210088. [Google Scholar] [CrossRef] [PubMed]
- Morran, L.T.; Schmidt, O.G.; Gelarden, I.A.; Parrish, R.C.; Lively, C.M. Running with the Red Queen: Host-parasite coevolution selects for biparental sex. Science 2011, 333, 216–218. [Google Scholar] [CrossRef]
- Arakelyan, M.; Harutyunyan, T.; Aghayan, S.A.; Carretero, M.A. Infection of parthenogenetic lizards by blood parasites does not support the “Red Queen hypothesis” but reveals the costs of sex. Zoology 2019, 136, 125709. [Google Scholar] [CrossRef] [PubMed]
- Majewska, A.A.; Sims, S.; Schneider, A.; Altizer, S.; Hall, R.J. Multiple transmission routes sustain high prevalence of a virulent parasite in a butterfly host. Proc. R. Soc. B 2019, 286, 20191630. [Google Scholar] [CrossRef] [PubMed]
- White, P.S.; Choi, A.; Pandey, R.; Menezes, A.; Penley, M.; Gibson, A.K.; de Roode, J.; Morran, L. Host heterogeneity mitigates virulence evolution. Biol. Lett. 2020, 16, 20190744. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csivincsik, Á.; Halász, T.; Nagy, G. The Large American Liver Fluke (Fascioloides magna): A Survivor’s Journey through a Constantly Changing World. Parasitologia 2023, 3, 300-326. https://doi.org/10.3390/parasitologia3040031
Csivincsik Á, Halász T, Nagy G. The Large American Liver Fluke (Fascioloides magna): A Survivor’s Journey through a Constantly Changing World. Parasitologia. 2023; 3(4):300-326. https://doi.org/10.3390/parasitologia3040031
Chicago/Turabian StyleCsivincsik, Ágnes, Tibor Halász, and Gábor Nagy. 2023. "The Large American Liver Fluke (Fascioloides magna): A Survivor’s Journey through a Constantly Changing World" Parasitologia 3, no. 4: 300-326. https://doi.org/10.3390/parasitologia3040031