
A Mortality Event Involving Endangered Pallid Sturgeon (Scaphirhynchus albus) Associated with Gyrodactylus conei n. sp. (Monogenea: Gyrodactylidae) Effectively Treated with Parasite-S (Formalin)
Abstract
:1. Introduction
2. Results
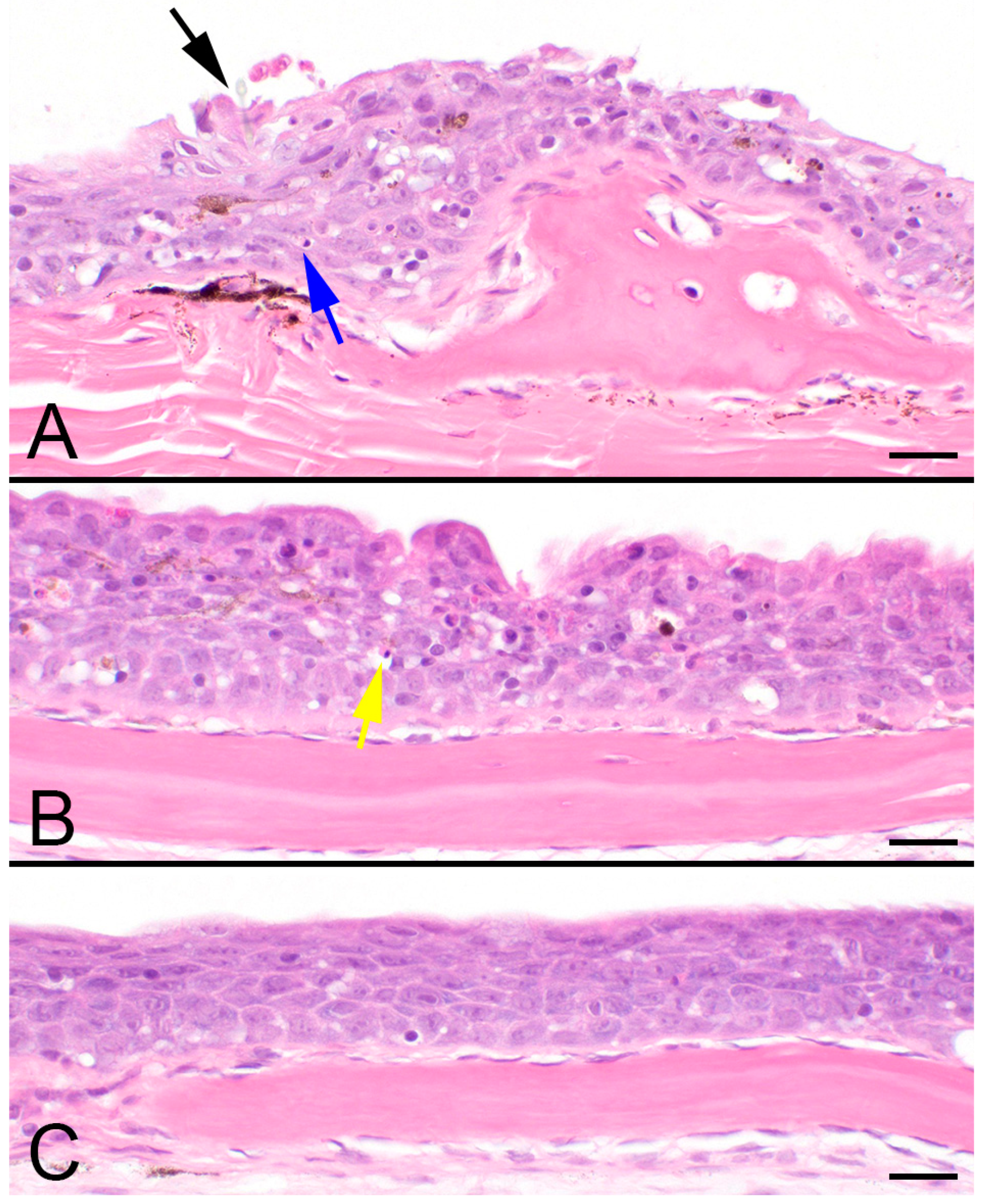
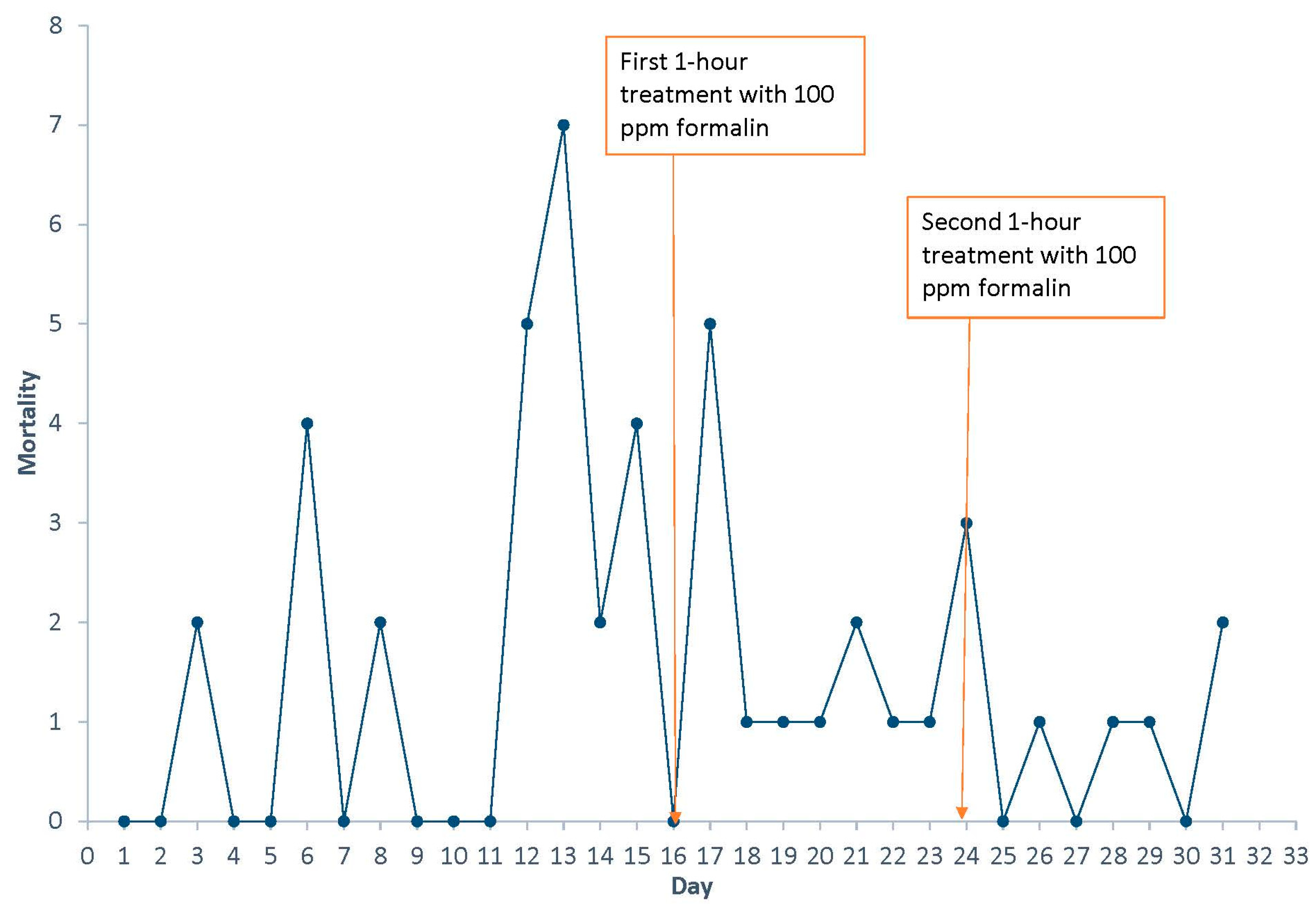
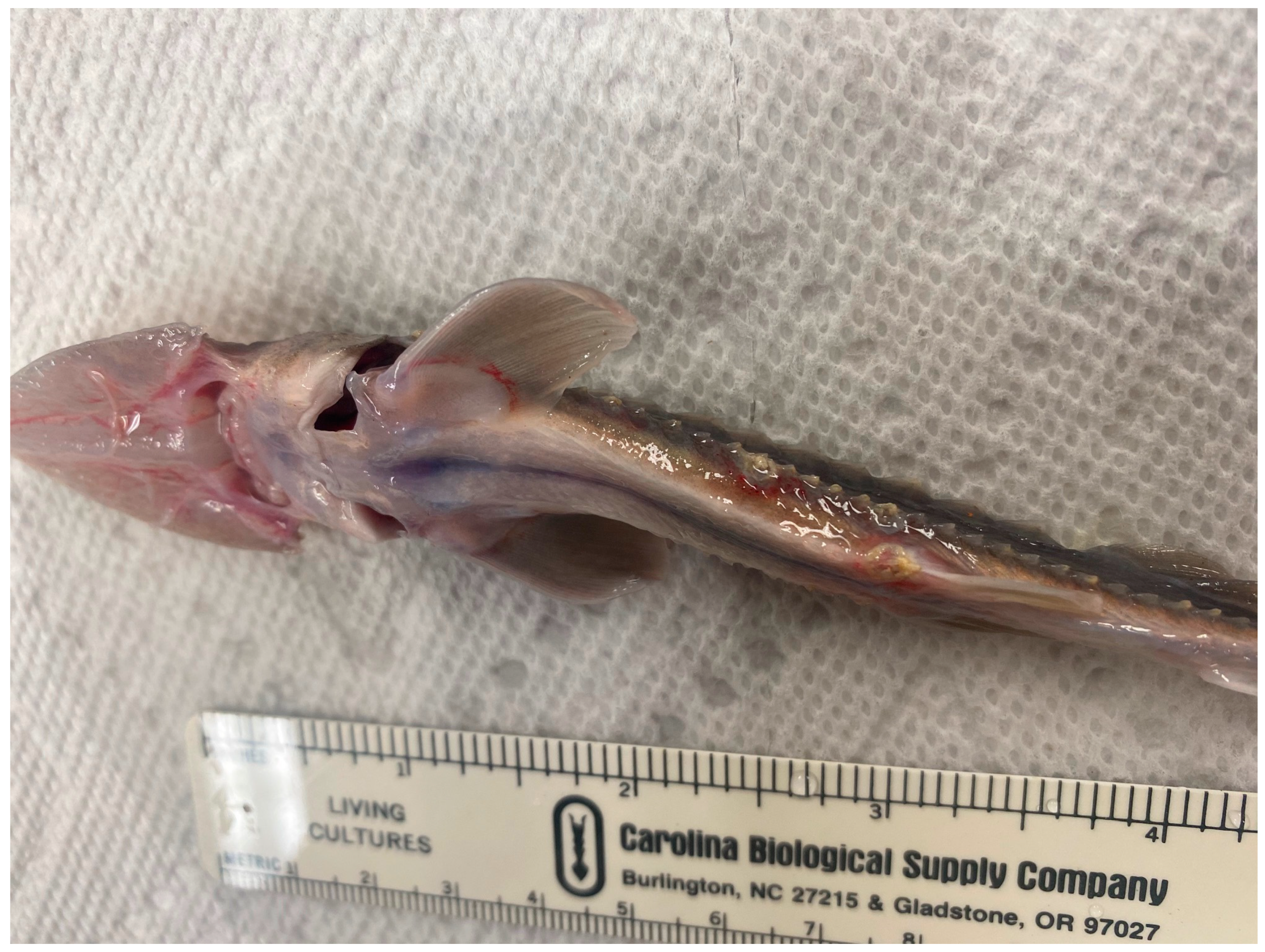
2.1. Diagnosis of Mortality Event
2.2. Treatment
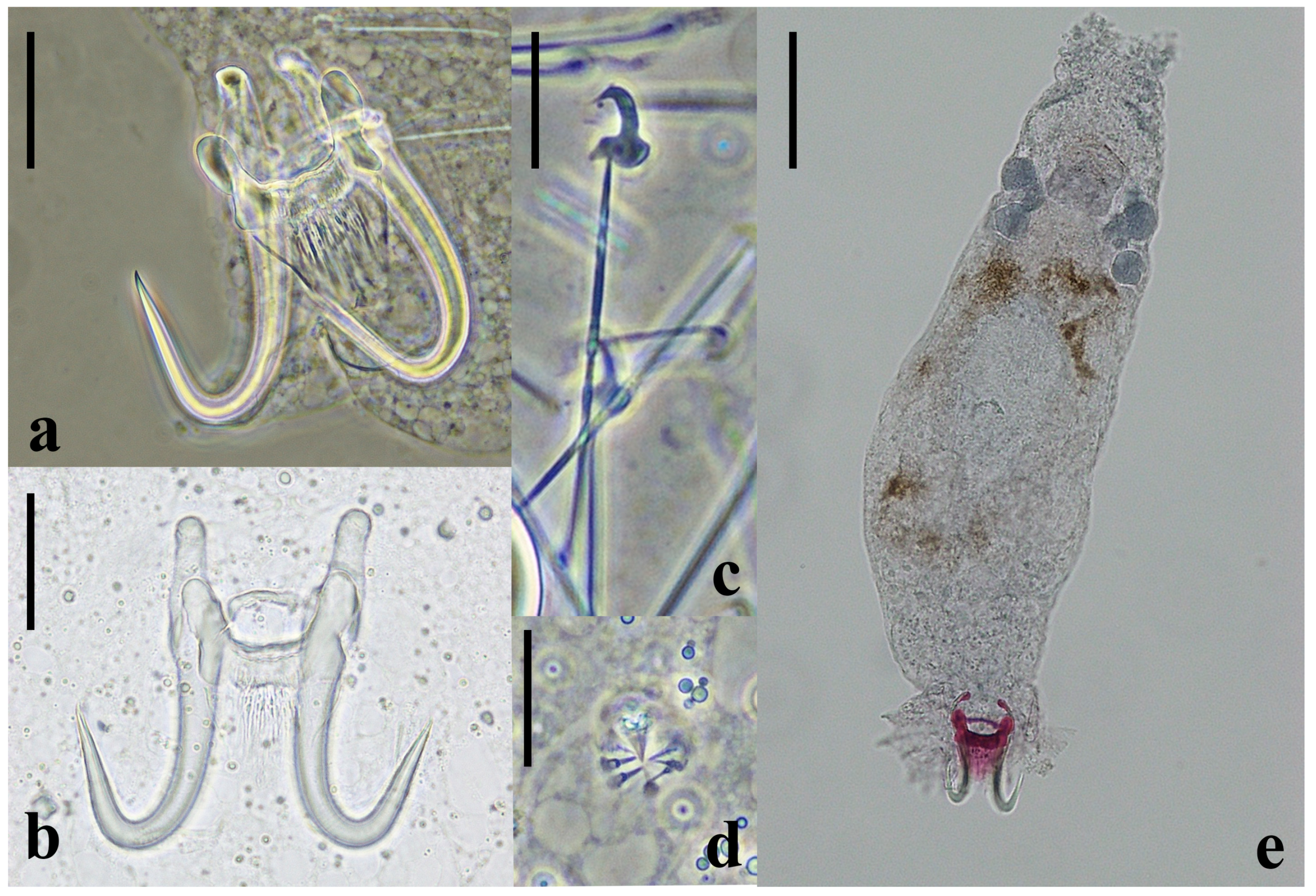
2.3. New Species Description
2.3.1. Molecular Identification
2.3.2. Remarks
3. Discussion
4. Materials and Methods
4.1. Description of Mortality Event
4.2. Treatment Protocol
4.3. Bacteriology Methods
4.4. Virology Methods
4.5. Histology Methods
4.6. Gyrodactylus Identification
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pflieger, W.L. The Fishes of Missouri; Missouri Department of Conservation: Jefferson City, MO, USA, 1975. [Google Scholar]
- US Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; Determination of Endangered Status for the Pallid Sturgeon. Fed. Regist. 1990, 55, 36641–36647. [Google Scholar]
- Bakke, T. Host Specificity Dynamics: Observations on Gyrodactylid Monogeneans. Int. J. Parasitol. 2002, 32, 281–308. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, G.L. Parasites of North American Freshwater Fish; Cornell University Press: Ithaca, NY, USA, 1999. [Google Scholar]
- Popielarczyk, R.; Kolman, R. Preliminary Analysis of Ectoparasites of the Sturgeon Acipenser Oxyrinchus Oxyrinchus (Mitchill, 1815) Originating from Different Water Habitats. Ann. Parasitol. 2013, 59, 139–141. [Google Scholar] [PubMed]
- Johnsen, B.O.; Jensen, A.J. The Gyrodactylus Story in Norway. Aquaculture 1991, 98, 289–302. [Google Scholar] [CrossRef]
- Leis, E.; Chi, T.K.; Lumme, J. Global Phylogeography of Salmonid Ectoparasites of the Genus Gyrodactylus, with an Emphasis on the Origin of the Circumpolar Gyrodactylus salmonis (Platyhelminthes: Monogenea). Comp. Parasitol. 2021, 88, 130–143. [Google Scholar] [CrossRef]
- Tuttle-Lau, M.; Leis, E.M.; Cupp, A.; Peterman, L.; Hebert, J.; Erickson, R.; Schleis, S.; Gaikowski, M. Efficacy of Hydrogen Peroxide to Reduce Gyrodactylus Species Infestation Density on Four Fish Species. J. Aquat. Anim. Health 2023. [Google Scholar] [CrossRef]
- Mizelle, J.D.; Kritsky, D.C. Studies on Monogenetic Trematodes. XXXIII. New Species of Gyrodactylus and a Key to the North American Species. Trans. Am. Microsc. Soc. 1967, 86, 390. [Google Scholar] [CrossRef]
- Zietara, M.S.; Lumme, J. Speciation by Host Switch and Adaptive Radiation in a Fish Parasite Genus Gyrodactylus (Monogenea, Gyrodactylidae). Evolution 2002, 56, 2445–2458. [Google Scholar] [CrossRef]
- Lumme, J.; Zietara, M.S.; Lebedeva, D. Ancient and Modern Genome Shuffling: Reticulate Mito-Nuclear Phylogeny of Four Related Allopatric Species of Gyrodactylus von Nordmann, 1832 (Monogenea: Gyrodactylidae), Ectoparasites on the Eurasian Minnow Phoxinus phoxinus (L.) (Cyprinidae). Syst. Parasitol. 2017, 94, 183–200. [Google Scholar] [CrossRef]
- Bruno, D.W.; Noguera, P.A.; Poppe, T.T. A Colour Atlas of Salmonid Diseases, 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Teruo, M. Color Atlas of Fish Histopathology; Shin-Suisan Shinbun-Sha, Ltd.: Tokyo, Japan, 2007; Volume 2. [Google Scholar]
- Leis, E.; King, S.; Leis, S.; Cone, D. Infections of Gyrodactylus crysoleucas and Gyrodactylus sp. (Monogenea) at a Golden Shiner (Notemigonus crysoleucas) Farm in Minnesota. Comp. Parasitol. 2016, 83, 105–110. [Google Scholar] [CrossRef]
- Cone, D.K.; Beverley-Burton, M.; Wiles, M.; McDonald, T.E. The Taxonomy of Gyrodactylus (Monogenea) Parasitizing Certain Salmonid Fishes of North America, with a Description of Gyrodactylus nerkae n. Sp. Can. J. Zool. 1983, 61, 2587–2597. [Google Scholar] [CrossRef]
- Rózsa, L.; Vas, Z. Co-Extinct and Critically Co-Endangered Species of Parasitic Lice, and Conservation-Induced Extinction: Should Lice Be Reintroduced to Their Hosts? Oryx 2015, 49, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Windsor, D.A. Conservation of Parasites. Int. J. Parasitol. Parasites Wildl. 2021, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Nichols, E. Neglected Wild Life: Parasitic Biodiversity as a Conservation Target. Int. J. Parasitol. Parasites Wildl. 2013, 2, 222–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- U.S. Fish and Wildlife Service. American Fisheries Society-Fish Health Section. Standard Procedures for Aquatic Animal Health Inspections. In AFS-FHS. FHS Blue Book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens, 2020 Edition. Available online: https://units.fisheries.org/fhs/fish-health-section-blue-book-2020/ (accessed on 25 May 2023).
- Lannan, C.N.; Winton, J.R.; Fryer, J.L. Fish Cell Lines: Establishment and Characterization of Nine Cell Lines from Salmonids. Vitro 1984, 20, 671–676. [Google Scholar] [CrossRef]
- Winton, J.; Batts, W.; DeKinkelin, P.; LeBerre, M.; Bremont, M.; Fijan, N. Current Lineages of the Epithelioma Papulosum Cyprini (EPC) Cell Line Are Contaminated with Fathead Minnow, Pimephales Promelas, Cells: Contamination of EPC Cells. J. Fish Dis. 2010, 33, 701–704. [Google Scholar] [CrossRef]
- Watson, L.; Yun, S.; Groff, J.; Hedrick, R. Characteristics and Pathogenicity of a Novel Herpesvirus Isolated from Adult and Subadult White Sturgeon Acipenser Transmontanus. Dis. Aquat. Org. 1995, 22, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Kurobe, T.; Kwak, K.; MacConnell, E.; McDowell, T.; Mardones, F.; Hedrick, R. Development of PCR Assays to Detect Iridovirus Infections among Captive and Wild Populations of Missouri River Sturgeon. Dis. Aquat. Org. 2010, 93, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Kritsky, D.C.; Leiby, P.D.; Kayton, R.J. A Rapid Stain Technique for the Haptoral Bars of Gyrodactylus Species (Monogenea). J. Parasitol. 1978, 64, 172–174. [Google Scholar] [CrossRef]
- Leis, E.; Easy, R.; Cone, D. A Variant of Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013 (Monogenea) Identified from Weed Shiner (Notropis texanus) in Wisconsin Portions of the Upper Mississippi River. Syst. Parasitol. 2021, 98, 247–253. [Google Scholar] [CrossRef]
- Malmberg, G. The Excretory Systems and the Marginal Hooks as a Basis for the Systematics of Gyrodactylus (Trematoda, Monogenea). Ark. Zool. 1970, 23, 1–235. [Google Scholar]
- Cone, D.; Abbott, C.; Gilmore, S.; Burt, M. A New Genus and Species of Gyrodactylid (Monogenea) from Silver Hake, Merluccius bilinearis, in the Bay of Fundy, New Brunswick, Canada. J. Parasitol. 2010, 96, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Cable, J.; Harris, P.D.; Tinsley, R.C.; Lazarus, C.M. Phylogenetic Analysis of Gyrodactylus spp. (Platyhelminthes: Monogenea) Using Ribosomal DNA Sequences. Can. J. Zool. 1999, 77, 1439–1449. [Google Scholar]
- Use of Fishes in Research Committee (Joint committee of the American Fisheries Society, the American Institute of Fishery Research Biologists, and the American Society of Ichthyologists and Herpetologists). Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MD, USA, 2014. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G. conei a | G. colemanensis b | G. colemanensis c | |
|---|---|---|---|
| Anchor | |||
| Length | 55(61) | 49 | 50 |
| Root | 14(17) | ‒ | 17 |
| Shaft | 43(46) | ‒ | 36 |
| Point | 23(27) | ‒ | 22 |
| Ventral Bar | |||
| Width | 25(27) | 31 | 26 |
| Length | 7(7) | ‒ | 7 |
| ALP Length | 10(9) | ‒ | 9 |
| Membrane Length | 17(17) | ‒ | 10 |
| Marginal Hook | |||
| Total Length | 31(32) | 29 | 31 |
| Sickle Length | 5(5) | 6 | 5 |
| Handle | 26(27) | ‒ | 27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leis, E.; Bailey, J.; Katona, R.; Standish, I.; Dziki, S.; McCann, R.; Perkins, J.; Eckert, N.; Baumgartner, W. A Mortality Event Involving Endangered Pallid Sturgeon (Scaphirhynchus albus) Associated with Gyrodactylus conei n. sp. (Monogenea: Gyrodactylidae) Effectively Treated with Parasite-S (Formalin). Parasitologia 2023, 3, 205-214. https://doi.org/10.3390/parasitologia3020021
Leis E, Bailey J, Katona R, Standish I, Dziki S, McCann R, Perkins J, Eckert N, Baumgartner W. A Mortality Event Involving Endangered Pallid Sturgeon (Scaphirhynchus albus) Associated with Gyrodactylus conei n. sp. (Monogenea: Gyrodactylidae) Effectively Treated with Parasite-S (Formalin). Parasitologia. 2023; 3(2):205-214. https://doi.org/10.3390/parasitologia3020021
Chicago/Turabian StyleLeis, Eric, Jennifer Bailey, Ryan Katona, Isaac Standish, Sara Dziki, Rebekah McCann, Justin Perkins, Nathan Eckert, and Wes Baumgartner. 2023. "A Mortality Event Involving Endangered Pallid Sturgeon (Scaphirhynchus albus) Associated with Gyrodactylus conei n. sp. (Monogenea: Gyrodactylidae) Effectively Treated with Parasite-S (Formalin)" Parasitologia 3, no. 2: 205-214. https://doi.org/10.3390/parasitologia3020021