

Predicted Secretome of the Monogenean Parasite Rhabdosynochus viridisi: Hypothetical Molecular Mechanisms for Host-Parasite Interactions

 ,
, 
Abstract
:1. Introduction
2. Results
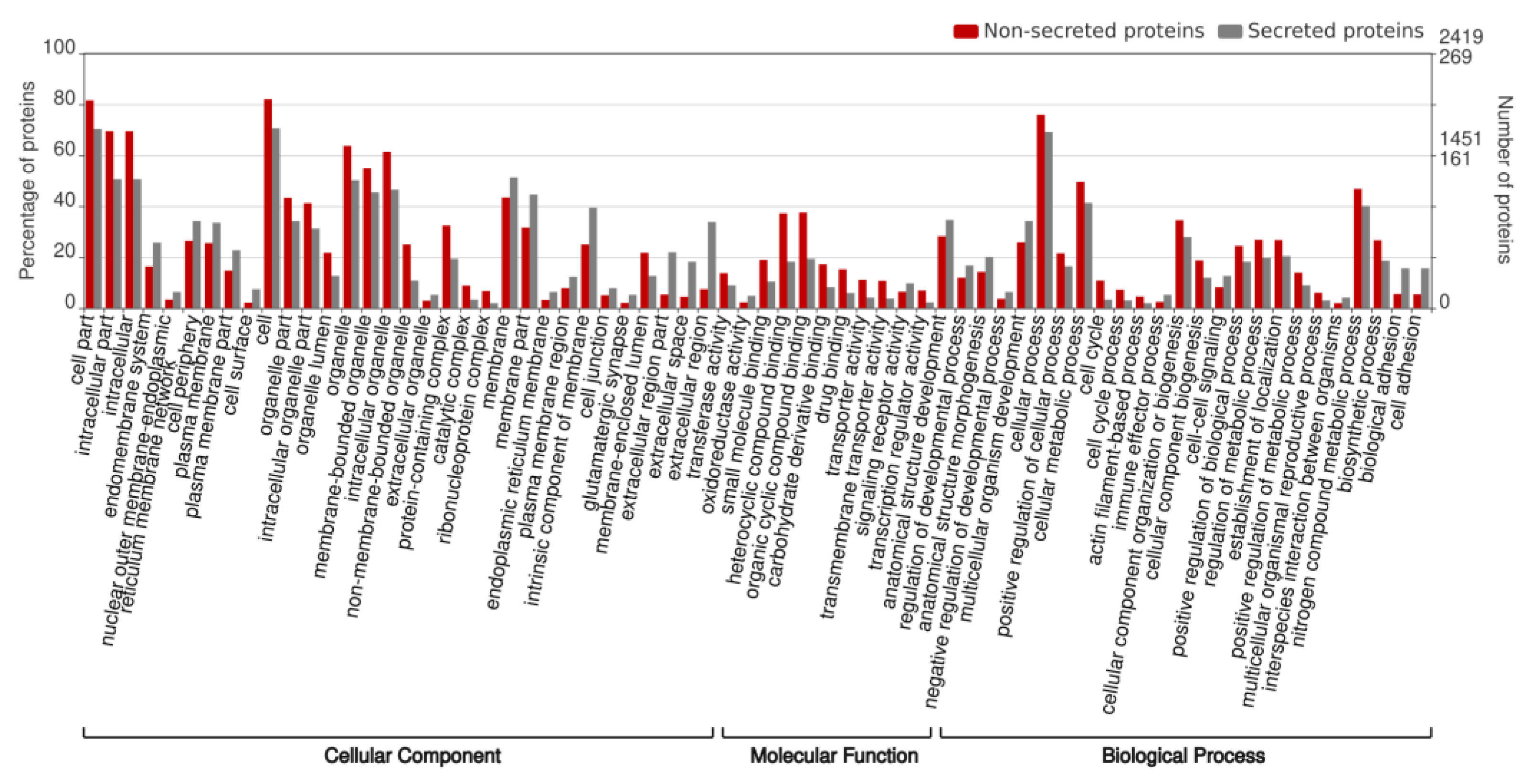
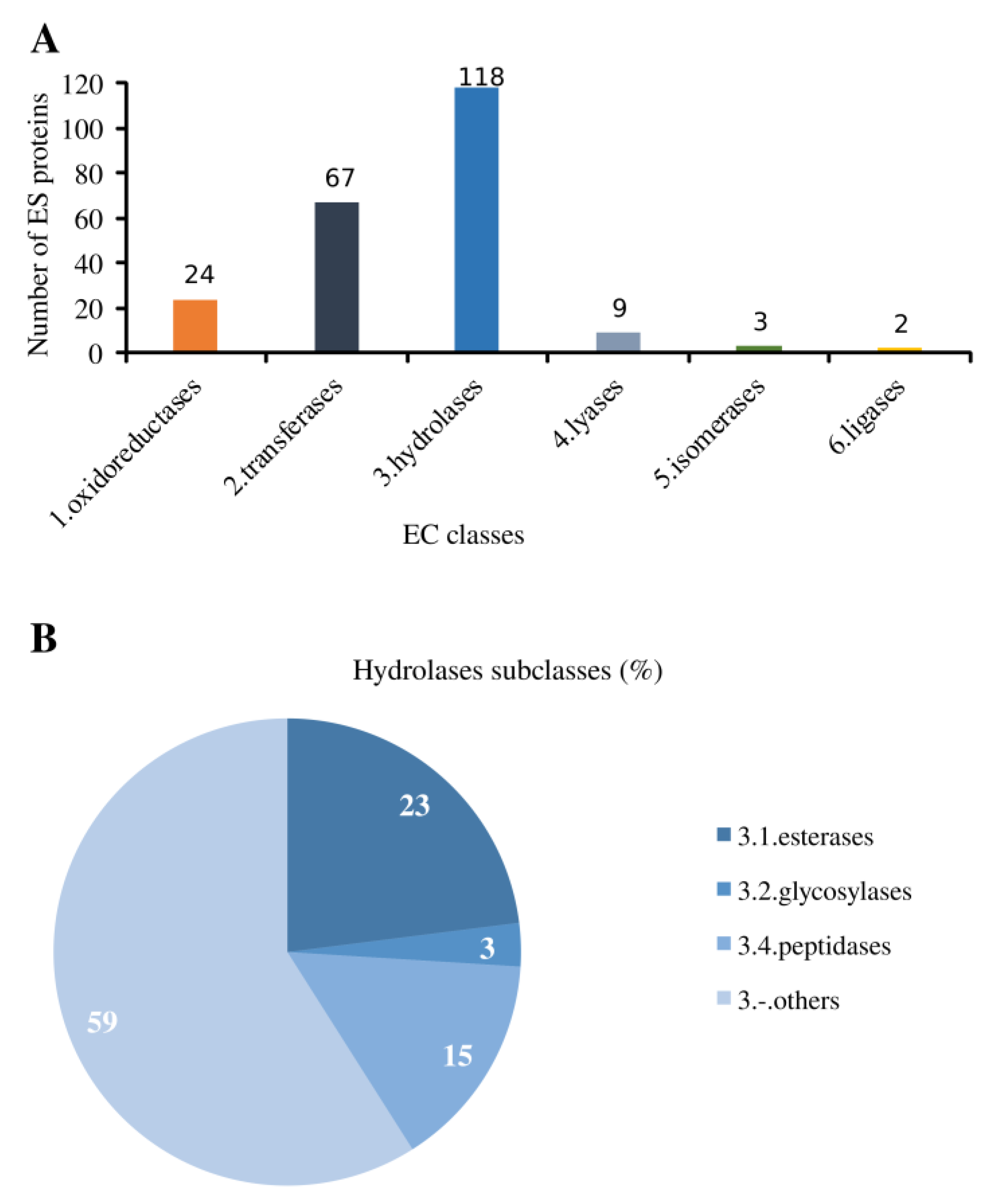
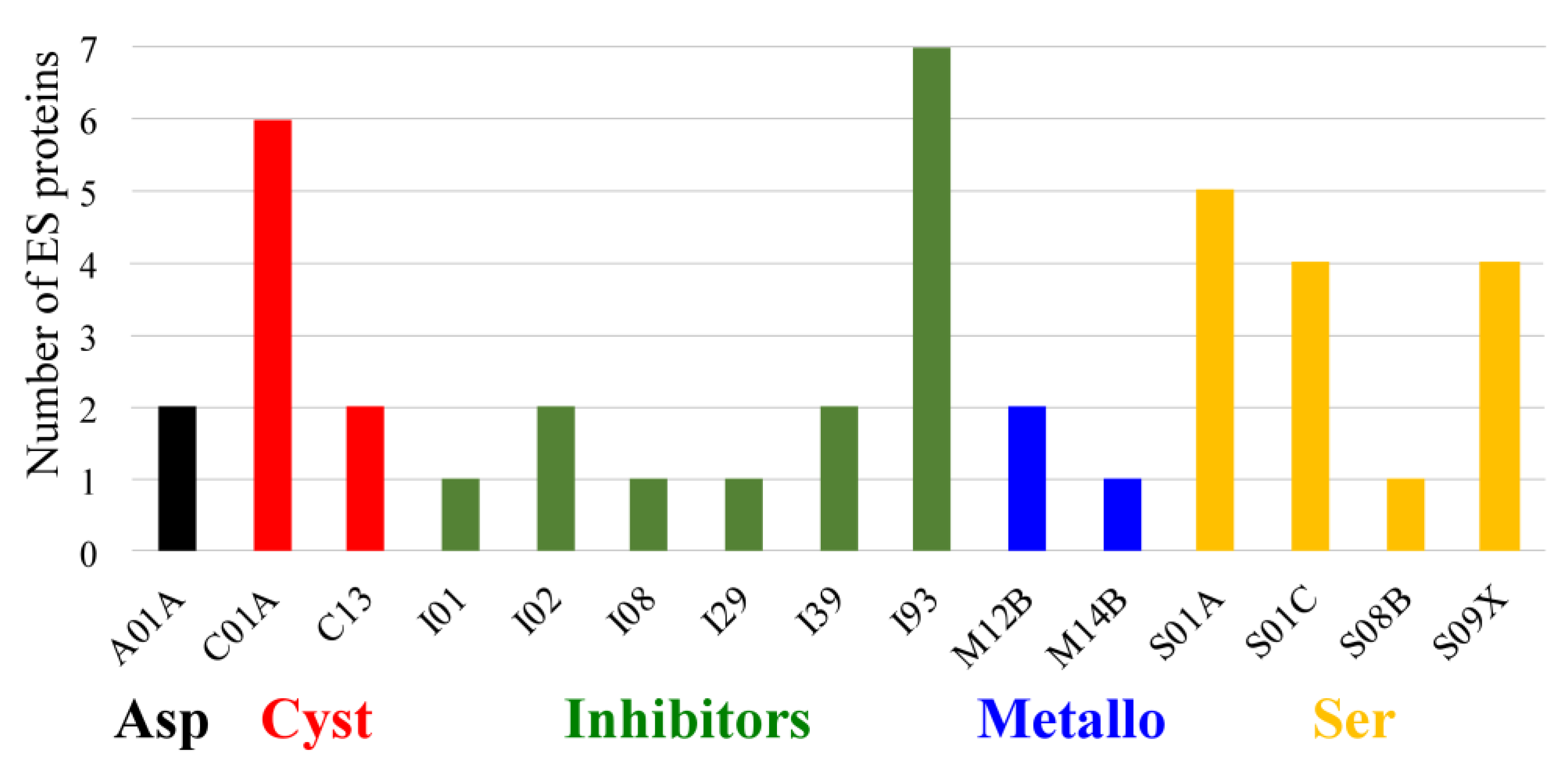
2.1. In Silico Identification and Functional Annotation of ES Proteins
2.2. Antigenicity Prediction of ES Proteins
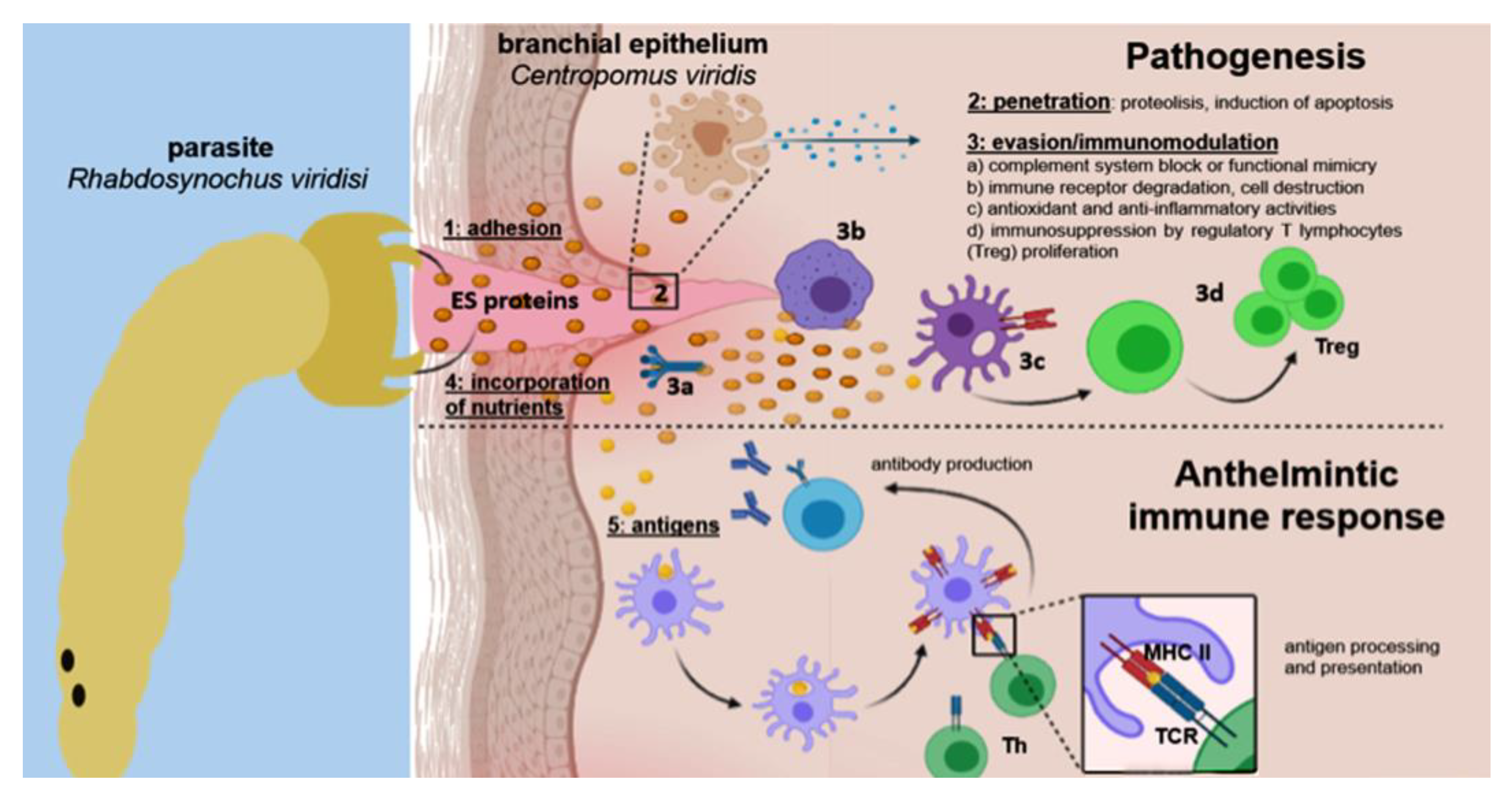
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whittington, I. Monogenea Monopisthocotylea (Ectoparasitic flukes). In Marine Parasitology; Rohde, K., Ed.; CSIRO Publishing: Melbourne, Australia, 2005; pp. 63–72. [Google Scholar]
- Busch, S.; Dalsgaard, I.; Buchmann, K. Concomitant exposure of rainbow trout fry to Gyrodactylus derjavini and Flavobacterium psychrophilum: Effects on infection and mortality of host. Vet. Parasitol. 2003, 117, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Dezfuli, B.S.; Giari, L.; Simoni, E.; Menegatti, R.; Shinn, A.P.; Manera, M. Gill histopathology of cultured European sea bass, Dicentrarchus labrax (L.), infected with Diplectanum aequans (Wagener 1857) Diesing 1958 (Diplectanidae: Monogenea). Parasitol. Res. 2007, 100, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Roumbedakis, K.; Marchiori, N.C.; Paseto, Á.; Gonçalves, E.L.; Luque, J.L.; Cepeda, P.B.; Sanches, E.G.; Martins, M.L. Parasite fauna of wild and cultured dusky-grouper Epinephelus marginatus (Lowe, 1834) from Ubatuba, southeastern Brazil. Braz. J. Biol. 2013, 73, 871–878. [Google Scholar] [CrossRef]
- Andree, K.B.; Roque, A.; Duncan, N.; Gisbert, E.; Estevez, A.; Tsertou, M.I.; Katharios, V. Diplectanum sciaenae (Van Beneden & Hesse, 1863) (Monogenea) infecting meagre, Argyrosomus regius (Asso, 1801) broodstock in Catalonia, Spain, A case report. Vet. Parasitol. Reg. Stud. Rep. 2015, 1–2, 75–79. [Google Scholar] [CrossRef]
- Khrukhayan, P.; Limsuwan, C.; Chuchird, N. Seasonal variation of diplectanid monogeneans in cage cultured seabass from Bangpakong River, Thailand. Kasetsart Univ. Fish. Res. Bull. 2016, 40, 68–78. [Google Scholar]
- Kritsky, D.C.; Bakenhaster, M.D.; Fajer-Avila, E.J.; Bullard, S.A. Rhabdosynochus spp. (Monogenoidea: Diplectanidae) infecting the gill lamellae of snooks, Centropomus spp. (Perciformes: Centropomidae), in Florida, and redescription of the type species, R. rhabdosynochus. J. Parasitol. 2010, 96, 879–886. [Google Scholar] [CrossRef]
- Montero-Rodríguez, M.; Mendoza-Franco, E.F.; López-Téllez, N.A. New species of Rhabdosynochus Mizelle and Blatz 1941 (Monogenoidea: Diplectanidae) from the gills of the cultured Centropomus viridis (Perciformes) in the Mexican Eastern Tropical Pacific. Acta Parasitol. 2021, 66, 487–492. [Google Scholar] [CrossRef]
- Morales-Serna, F.N.; López-Moreno, D.G.; Medina-Guerrero, R.M.; Abad-Rosales, S.M.; Martínez-Brown, J.M.; Ibarra-Castro, L.; Fajer-Avila, E.J. Toxicity of formalin for juvenile Centropomus viridis and in vitro efficacy against the parasite Rhabdosynochus sp. (Monogenea: Diplectanidae). J. Appl. Ichthyol. 2020, 36, 740–744. [Google Scholar] [CrossRef]
- Baldini, G.; Santamaría-Miranda, A.; Martínez-Brown, J.M.; Ibarra-Castro, L. Technical-economic viability of white snook Centropomus viridis culture in floating cages in a coastal lagoon in northwestern Mexico. Aquac. Rep. 2022, 23, 101048. [Google Scholar] [CrossRef]
- Virginio, V.G.; Monteiro, K.M.; Drumond, F.; de Carvalho, M.O.; Vargas, D.M.; Zaha, A.; Ferreira, H.B. Excretory/secretory products from in vitro-cultured Echinococcus granulosus protoscoleces. Mol. Biochem. Parasitol. 2012, 183, 15–22. [Google Scholar] [CrossRef]
- Nono, J.K.; Pletinckx, K.; Lutz, M.B.; Brehm, K. Excretory/secretory-products of Echinococcus multilocularis larvae induce apoptosis and tolerogenic properties in dendritic cells in vitro. PLoS Negl. Trop. Dis. 2012, 6, e1516. [Google Scholar] [CrossRef]
- Ren, H.N.; Zhuo, T.X.; Bai, S.J.; Bai, Y.; Sun, X.Y.; Liu, R.D.; Long, S.R.; Cui, J.; Wang, Z.Q. Proteomic analysis of hydrolytic proteases in excretory/secretory proteins from Trichinella spiralis intestinal infective larvae using zymography combined with shotgun LC-MS/MS approach. Acta Trop. 2021, 216, 105825. [Google Scholar] [CrossRef] [PubMed]
- Ilgová, J.; Salát, J.; Kašný, M. Molecular communication between the monogenea and fish immune system. Fish Shellfish Immunol. 2021, 112, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Prasad, A. Taenia solium proteins: A beautiful kaleidoscope of pro and anti-inflammatory antigens. Expert Rev. Proteom. 2020, 17, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Sitjà-Bobadilla, A. Living off a fish: A trade-off between parasites and the immune system. Fish Shellfish Immunol. 2008, 25, 358–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakeri, A.; Hansen, E.P.; Andersen, S.D.; Williams, A.R.; Nejsum, P. Immunomodulation by helminths: Intracellular pathways and extracellular vesicles. Front. Immunol. 2018, 9, 2349. [Google Scholar] [CrossRef] [Green Version]
- Host-Pathogen Interaction and Immune Evasion. Available online: https://www.immunology.org/file/2350/download?token=Xrx18BQ2 (accessed on 14 March 2022).
- Cuesta-Astroz, Y.; Oliveira, F.S.; Nahum, L.A.; Oliveira, G. Helminth secretomes reflect different lifestyles and parasitized hosts. Int. J. Parasitol. 2017, 47, 529–544. [Google Scholar] [CrossRef]
- Młocicki, D.; Sulima, A.; Bień, J.; Näreaho, A.; Zawistowska-Deniziak, A.; Basałaj, K.; Sałamatin, R.; Conn, D.B.; Savijoki, K. Immunoproteomics and surfaceomics of the adult tapeworm Hymenolepis diminuta. Front. Immunol. 2018, 9, 2487. [Google Scholar] [CrossRef] [Green Version]
- Mazanec, H.; Koník, P.; Gardian, Z.; Kuchta, R. Extracellular vesicles secreted by model tapeworm Hymenolepis diminuta: Biogenesis, ultrastructure and protein composition. Int. J. Parasitol. 2021, 51, 327–332. [Google Scholar] [CrossRef]
- Wititkornkul, B.; Hulme, B.J.; Tomes, J.J.; Allen, N.R.; Davis, C.N.; Davey, S.D.; Cookson, A.R.; Phillips, H.C.; Hegarty, M.J.; Swain, M.T.; et al. Evidence of immune modulators in the secretome of the equine tapeworm Anoplocephala perfoliata. Pathogens 2021, 10, 912. [Google Scholar] [CrossRef]
- Caña-Bozada, V.; Chapa-López, M.; Díaz-Martín, R.D.; García-Gasca, A.; Huerta-Ocampo, J.Á.; de Anda-Jáuregui, G.; Morales-Serna, F.N. In silico identification of excretory/secretory proteins and drug targets in monogenean parasites. Infect. Genet. Evol. 2021, 93, 104931. [Google Scholar] [CrossRef] [PubMed]
- Vorel, J.; Cwiklinski, K.; Roudnický, P.; Ilgová, J.; Jedličková, L.; Dalton, J.P.; Mikeš, L.; Gelnar, M.; Kašný, M. Eudiplozoon nipponicum (Monogenea, Diplozoidae) and its adaptation to haematophagy as revealed by transcriptome and secretome profiling. BMC Genom. 2021, 22, 274. [Google Scholar] [CrossRef] [PubMed]
- Gahoi, S.; Singh, S.; Gautam, B. Genome-wide identification and comprehensive analysis of Excretory/Secretory proteins in nematodes provide potential drug targets for parasite control. Genomics 2019, 111, 297–309. [Google Scholar] [CrossRef]
- Caña-Bozada, V.; Morales-Serna, F.N.; Fajer-Ávila, E.J.; Llera-Herrera, R. De novo transcriptome assembly and identification of G-Protein-Coupled-Receptors (GPCRs) in two species of monogenean parasites of fish. Parasite 2022, 29, 51. [Google Scholar] [CrossRef] [PubMed]
- Whittington, I.D.; Cribb, B.W. Adhesive secretions in the Platyhelminthes. Adv. Parasitol. 2001, 48, 101–224. [Google Scholar] [CrossRef]
- Kearn, G.C. Some aspects of the biology of monogenean (platyhelminth) parasites of marine and freshwater fishes. Oceanography 2014, 2, 117. [Google Scholar] [CrossRef]
- Yadati, T.; Houben, T.; Bitorina, A.; Shiri-Sverdlov, R. The ins and outs of cathepsins: Physiological function and role in disease management. Cells 2020, 9, 1679. [Google Scholar] [CrossRef]
- Liu, R.D.; Meng, X.Y.; Li, C.L.; Long, S.R.; Cui, J.; Wang, Z.Q. Molecular characterization and determination of the biochemical properties of cathepsin L of Trichinella spiralis. Vet. Res. 2022, 53, 48. [Google Scholar] [CrossRef]
- Rao, Y.; Yang, T. cDNA cloning, mRNA expression and recombinant expression of a cathepsin L-like cysteine protease from Neobenedenia melleni (Monogenea: Capsalidae). Aquaculture 2007, 269, 41–53. [Google Scholar] [CrossRef]
- Jedličková, L.; Dvořáková, H.; Dvořák, J.; Kašný, M.; Ulrychová, L.; Vorel, J.; Žárský, V.; Mikeš, L. Cysteine peptidases of Eudiplozoon nipponicum: A broad repertoire of structurally assorted cathepsins L in contrast to the scarcity of cathepsins B in an invasive species of haematophagous monogenean of common carp. Parasites Vectors 2018, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Cantacessi, C.; Hofmann, A.; Pickering, D.; Navarro, S.; Mitreva, M.; Loukas, A. TIMPs of parasitic helminths—A large-scale analysis of high-throughput sequence datasets. Parasites Vectors 2013, 6, 156. [Google Scholar] [CrossRef] [PubMed]
- Roudnický, P.; Vorel, J.; Ilgová, J.; Benovics, M.; Norek, A.; Jedličková, L.; Mikeš, L.; Potěšil, D.; Zdráhal, Z.; Dvořák, J.; et al. Identification and partial characterization of a novel serpin from Eudiplozoon nipponicum (Monogenea, Polyopisthocotylea). Parasite 2018, 25, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilgová, J.; Kavanová, L.; Matiašková, K.; Salát, J.; Kašný, M. Effect of cysteine peptidase inhibitor of Eudiplozoon nipponicum (Monogenea) on cytokine expression of macrophages in vitro. Mol. Biochem. Parasitol. 2020, 235, 111248. [Google Scholar] [CrossRef] [PubMed]
- Franco-Serrano, L.; Sánchez-Redondo, D.; Nájar-García, A.; Hernández, S.; Amela, I.; Perez-Pons, J.A.; Piñol, J.; Mozo-Villarias, A.; Cedano, J.; Querol, E. Pathogen moonlighting proteins: From ancestral key metabolic enzymes to virulence factors. Microorganisms 2021, 9, 1300. [Google Scholar] [CrossRef]
- Ditgen, D.; Anandarajah, E.M.; Hansmann, J.; Winter, D.; Schramm, G.; Erttmann, K.D.; Liebau, E.; Brattig, N.W. Multifunctional thioredoxin-like protein from the gastrointestinal parasitic nematodes Strongyloides ratti and Trichuris suis affects mucosal homeostasis. J. Parasitol. Res. 2016, 2016, 8421597. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Dee, Z.; Pidcock, K.; Gutierrez, L.S. Thrombospondin-1: Multiple paths to inflammation. Mediat. Inflamm. 2011, 2011, 296069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haçarız, O.; Akgün, M.; Kavak, P.; Yüksel, B.; Sağıroğlu, M.Ş. Comparative transcriptome profiling approach to glean virulence and immunomodulation-related genes of Fasciola hepatica. BMC Genom. 2015, 16, 366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Li, M.; Yin, L.; Fu, G.; Liu, Z. Role of thrombospondin-1 and thrombospondin-2 in cardiovascular diseases (Review). Int. J. Mol. Med. 2020, 45, 1275–1293. [Google Scholar] [CrossRef] [Green Version]
- Hebert, F.O.; Phelps, L.; Samonte, I.; Panchal, M.; Grambauer, S.; Barber, I.; Kalbe, M.; Landry, C.R.; Aubin-Horth, N. Identification of candidate mimicry proteins involved in parasite-driven phenotypic changes. Parasites Vectors 2015, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Mei, S.; Zhang, K. In silico unravelling pathogen-host signaling cross-talks via pathogen mimicry and human protein-protein interaction networks. Comput. Struct. Biotechnol. J. 2020, 18, 100–113. [Google Scholar] [CrossRef]
- Teichmann, S.A.; Chothia, C. Immunoglobulin superfamily proteins in Caenorhabditis elegans. J. Mol. Biol. 2000, 296, 1367–1383. [Google Scholar] [CrossRef] [PubMed]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschellà, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, C.M.; Dernburg, A.F. A family of zinc-finger proteins is required for chromosome-specific pairing and synapsis during meiosis in C. elegans. Dev. Cell 2006, 11, 817–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Li, J.; Xiang, M.; Hong, Q.; Xu, B.; Hu, W. Identification and characterization of the zinc finger protein SjZF in Schistosoma japonicum. Biochem. Biophys. Res. Commun. 2018, 501, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Rakhra, G.; Rakhra, G. Zinc finger proteins: Insights into the transcriptional and post transcriptional regulation of immune response. Mol. Biol. Rep. 2021, 48, 5735–5743. [Google Scholar] [CrossRef]
- Matsushima, N.; Takatsuka, S.; Miyashita, H.; Kretsinger, R.H. Leucine rich repeat proteins: Sequences, mutations, structures and diseases. Protein Pept. Lett. 2019, 26, 108–131. [Google Scholar] [CrossRef]
- Westrich, J.A.; Warren, C.D.; Pyeon, D. Evasion of host immune defenses by human papillomavirus. Virus Res. 2017, 231, 21–33. [Google Scholar] [CrossRef]
- Kim, J.G.; Shin, H.-C.; Seo, T.; Nawale, L.; Han, G.; Kim, B.Y.; Kim, S.J.; Cha-Molstad, H. Signaling Pathways Regulated by UBR Box-Containing E3 Ligases. Int. J. Mol. Sci. 2021, 22, 8323. [Google Scholar] [CrossRef]
- Monni, G.; Cognetti-Varriale, A.M. Antigenic activity of Diplectanum aequans (Monogenea) in sea bass (Dicentrarchus labrax L.) held under different oxygenation conditions. Bull. Eur. Assoc. Fish Pathol. 2001, 21, 241–245. [Google Scholar]
- Hatanaka, A.; Umeda, N.; Yamashita, S.; Hirazawa, N. A small ciliary surface glycoprotein of the monogenean parasite Neobenedenia girellae acts as an agglutination/immobilization antigen and induces an immune response in the Japanese flounderParalichthys olivaceus. Parasitology 2005, 131, 591. [Google Scholar] [CrossRef]
- Sandoval-Gío, J.J.; Rodríguez-Canul, R.; Vidal-Martínez, V.M. Humoral antibody response of the tilapia Oreochromis niloticus against Cichlidogyrus spp. (Monogenea). J. Parasitol. 2008, 94, 404–409. [Google Scholar] [CrossRef]
- Kishimori, J.M.; Takemura, A.; Leong, J.A. Neobenedenia melleni-specific antibodies are associated with protection after continuous exposure in Mozambique tilapia. J. Immunol. Res. 2015, 2015, 635387. [Google Scholar] [CrossRef] [PubMed]
- Gomez, S.; Adalid-Peralta, L.; Palafox-Fonseca, H.; Cantu-Robles, V.A.; Soberón, X.; Sciutto, E.; Fragoso, G.; Bobes, R.J.; Laclette, J.P.; Yauner, L.; et al. Genome analysis of Excretory/Secretory proteins in Taenia solium reveals their Abundance of Antigenic Regions (AAR). Sci. Rep. 2015, 5, 9683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motran, C.C.; Silvane, L.; Chiapello, L.S.; Theumer, M.G.; Ambrosio, L.F.; Volpini, X.; Celias, D.P.; Cervi, L. Helminth infections: Recognition and modulation of the immune response by innate immune cells. Front. Immunol. 2018, 9, 664. [Google Scholar] [CrossRef] [PubMed]
- Moreau, E.; Chauvin, A. Immunity against helminths: Interactions with the host and the intercurrent infections. J. Biomed. Biotechnol. 2010, 2010, 428593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.J.; Ha, G.H.; Seo, W.Y.; Kim, C.K.; Kim, K.; Lee, S.B. Human collagen alpha-2 type I stimulates collagen synthesis, wound healing, and elastin production in normal human dermal fibroblasts (HDFs). BMB Rep. 2020, 53, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, A.; Kalluri, R. Molecular Mechanisms of Angiostasis. In Encyclopedia of the Eye, Four-Volume Set; Elsevier: Amsterdam, The Netherlands, 2010; pp. 52–59. [Google Scholar]
- Bout, D.; Carlier, Y. Helminth functional antigens (with special reference to S. mansoni). Pathol. Biol. 1982, 30, 176–187. [Google Scholar] [PubMed]
- Mahendran, R.; Suganya, J.; Gayathri, S.; Dinakaran, M.; Vincent, A. Computer -aided vaccine designing approach against fish pathogens Edwardsiella tarda and Flavobacterium columnare using bioinformatics softwares. Drug Des. Dev. Ther. 2016, 10, 1703–1714. [Google Scholar] [CrossRef] [Green Version]
- Jo, C.H.; Son, J.; Kim, S.; Oda, T.; Kim, J.; Lee, M.R.; Sato, M.; Kim, H.T.; Unzai, S.; Park, S.Y.; et al. Structural insights into a 20.8-kDa tegumental-allergen-like (TAL) protein from Clonorchis sinensis. Sci. Rep. 2017, 7, 1764. [Google Scholar] [CrossRef] [Green Version]
- Maiti, B.; Dubey, S.; Munang’andu, H.M.; Karunasagar, I.; Karunasagar, I.; Evensen, Ø. Application of outer membrane protein-based vaccines against major bacterial fish pathogens in India. Front. Immunol. 2020, 11, 1362. [Google Scholar] [CrossRef]
- Sanches, R.C.O.; Tiwari, S.; Ferreira, L.C.G.; Oliveira, F.M.; Lopes, M.D.; Passos, M.J.F.; Maia, E.H.B.; Taranto, A.G.; Kato, R.; Azevedo, V.A.C.; et al. Immunoinformatics design of multi-epitope peptide-based vaccine against Schistosoma mansoni using transmembrane proteins as a target. Front. Immunol. 2021, 12, 621706. [Google Scholar] [CrossRef] [PubMed]
- Wilbers, R.H.P.; Schneiter, R.; Holterman, M.H.M.; Drurey, C.; Smant, G.; Asojo, O.A.; Maizels, R.M.; Lozano-Torres, J.L. Secreted venom allergen-like proteins of helminths: Conserved modulators of host responses in animals and plants. PLoS Pathog. 2018, 14, e1007300. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Filbey, K.J.; Esser-von Bieren, J.; Camberis, M.; Schwartz, C.; Murray, J.; Reynolds, L.A.; Blair, N.; Robertson, E.; Harcus, Y.; et al. Concerted activity of IgG1 antibodies and IL-4/IL-25-dependent effector cells trap helminth larvae in the tissues following vaccination with defined secreted antigens, providing sterile immunity to challenge infection. PLoS Pathog. 2015, 11, e1004676. [Google Scholar] [CrossRef] [PubMed]
- Darwiche, R.; Lugo, F.; Drurey, C.; Varossieau, K.; Smant, G.; Wilbers, R.; Maizels, R.M.; Schneiter, R.; Asojo, O.A. Crystal structure of Brugia malayi venom allergen-like protein-1 (BmVAL-1), a vaccine candidate for lymphatic filariasis. Int. J. Parasitol. 2018, 48, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Farias, L.P.; Chalmers, I.W.; Perally, S.; Rofatto, H.K.; Jackson, C.J.; Brown, M.; Khouri, M.I.; Barbosa, M.; Hensbergen, P.J.; Hokke, C.H.; et al. Schistosoma mansoni venom allergen-like proteins: Phylogenetic relationships, stage-specific transcription and tissue localization as predictors of immunological cross-reactivity. Int. J. Parasitol. 2019, 49, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, I.W.; Hoffmann, K.F. Platyhelminth Venom Allergen-Like (VAL) proteins: Revealing structural diversity, class-specific features and biological associations across the phylum. Parasitology 2012, 139, 1231–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regn, M.; Laggerbauer, B.; Jentzsch, C.; Ramanujam, D.; Ahles, A.; Sichler, S.; Calzada-Wack, J.; Koenen, R.R.; Braun, A.; Nieswandt, B.; et al. Peptidase inhibitor 16 is a membrane-tethered regulator of chemerin processing in the myocardium. J. Mol. Cell. Cardiol. 2016, 99, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, G.K.; Kaur, J.; Adya, R.; Miras, A.D.; Mattu, H.S.; Hattersley, J.G.; Kaltsas, G.; Tan, B.K.; Randeva, H.S. Chemerin induces endothelial cell inflammation: Activation of nuclear factor-kappa beta and monocyte-endothelial adhesion. Oncotarget 2018, 9, 16678–16690. [Google Scholar] [CrossRef] [Green Version]
- Mohandas, N.; Young, N.D.; Jabbar, A.; Korhonen, P.K.; Koehler, A.V.; Amani, P.; Hall, R.S.; Sternberg, P.W.; Jex, A.R.; Hofmann, A.; et al. The barber’s pole worm CAP protein superfamily—A basis for fundamental discovery and biotechnology advances. Biotechnol. Adv. 2015, 33, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Sotillo, J.; Pearson, M.S.; Becker, L.; Mekonnen, G.G.; Amoah, A.S.; van Dam, G.; Corstjens, P.L.A.M.; Murray, J.; Mduluza, T.; Mutapi, F.; et al. In-depth proteomic characterization of Schistosoma haematobium: Towards the development of new tools for elimination. PLoS Negl. Trop. Dis. 2019, 13, e0007362. [Google Scholar] [CrossRef] [Green Version]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISY1 Protein Expression Summary. Available online: https://www.proteinatlas.org/ENSG00000240682-ISY1 (accessed on 23 February 2022).
- Liu, Z.C.; Zhang, R.; Zhao, F.; Chen, Z.M.; Liu, H.W.; Wang, Y.J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.P.; et al. Venomic and transcriptomic analysis of centipede Scolopendra subspinipes dehaani. J. Proteome Res. 2012, 11, 6197–6212. [Google Scholar] [CrossRef]
- Wei, X.; Li, H.; Zhang, Y.; Li, C.; Li, K.; Ai, K.; Yang, J. Ca2+–calcineurin axis–controlled NFAT nuclear translocation is crucial for optimal T cell immunity in an early vertebrate. J Immunol. 2020, 204, 569–585. [Google Scholar] [CrossRef]
- WormBase ParaSite. Available online: https://parasite.wormbase.org (accessed on 21 August 2021).
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Franco-Serrano, L.; Hernández, S.; Calvo, A.; Severi, M.A.; Ferragut, G.; Pérez-Pons, J.A.; Piñol, J.; Pich, O.C.; Mozo-Villarias, A.; Amela, I.; et al. MultitaskProtDB-II: An update of a database of multitasking/moonlighting proteins. Nucleic Acids Res. 2018, 46, D645–D648. [Google Scholar] [CrossRef]
- Törönen, P.; Medlar, A.; Holm, L. PANNZER2: A rapid functional annotation web server. Nucleic Acids Res. 2018, 46, W84–W88. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A.; et al. WEGO 2.0: A web tool for analyzing and plotting GO annotations. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology and GO Annotations. Available online: https://www.ebi.ac.uk/QuickGO/ (accessed on 7 October 2021).
- Dalkiran, A.; Rifaioglu, A.S.; Martin, M.J.; Cetin-Atalay, R.; Atalay, V.; Doğan, T. ECPred: A tool for the prediction of the enzymatic functions of protein sequences based on the EC nomenclature. BMC Bioinform. 2018, 19, 334. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumor antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Phyre2. Available online: http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index (accessed on 30 November 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Immunomodulators | Protein ID | Annotation or PDB Template Name | Vaxijen Score |
|---|---|---|---|
| More antigenic proteins | DN725_c0_g1_i1.p1 | Collagen alpha-2(I) chain | 1.3983 |
| DN18884_c0_g6_i2.p1 | Unknown: nigellin-1.1 | 1.3867 | |
| DN2184_c0_g1_i6.p6 | Unknown: fkbp-type peptidyl-prolyl cis-trans isomerase slyd | 1.3846 | |
| DN146924_c0_g3_i1.p1 | Unknown: SH3-like barrel | 1.2806 | |
| DN9007_c0_g1_i4.p2 | Unknown: protein phosphatase 1 regulatory subunit 3a | 1.2422 | |
| DN8473_c0_g5_i3.p1 | Unknown: ribosomal protein L14e | 1.2028 | |
| Venom allergen-like (VAL) proteins | DN106_c0_g1_i4.p14 | GLIPR1-like protein 1 | 0.6280 |
| DN1638_c0_g1_i10.p1 | Pathogenesis-related protein | 0.5865 | |
| DN1553_c0_g1_i1.p1 | Peptidase inhibitor 16 | 0.6481 | |
| DN2123_c1_g1_i5.p1 | Unknown: venom allergen-like protein 4 | 0.4954 | |
| DN2793_c0_g1_i1.p7 | Pre-mRNA-splicing factor ISY1 homolog | 0.4541 | |
| DN537_c0_g1_i4.p3 | Scoloptoxin SSD976 | 0.4269 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirabent-Casals, M.; Caña-Bozada, V.H.; Morales-Serna, F.N.; García-Gasca, A. Predicted Secretome of the Monogenean Parasite Rhabdosynochus viridisi: Hypothetical Molecular Mechanisms for Host-Parasite Interactions. Parasitologia 2023, 3, 33-45. https://doi.org/10.3390/parasitologia3010004
Mirabent-Casals M, Caña-Bozada VH, Morales-Serna FN, García-Gasca A. Predicted Secretome of the Monogenean Parasite Rhabdosynochus viridisi: Hypothetical Molecular Mechanisms for Host-Parasite Interactions. Parasitologia. 2023; 3(1):33-45. https://doi.org/10.3390/parasitologia3010004
Chicago/Turabian StyleMirabent-Casals, Marian, Víctor Hugo Caña-Bozada, Francisco Neptalí Morales-Serna, and Alejandra García-Gasca. 2023. "Predicted Secretome of the Monogenean Parasite Rhabdosynochus viridisi: Hypothetical Molecular Mechanisms for Host-Parasite Interactions" Parasitologia 3, no. 1: 33-45. https://doi.org/10.3390/parasitologia3010004