
The Role of Acetate Kinase in the Human Parasite Entamoeba histolytica

Abstract
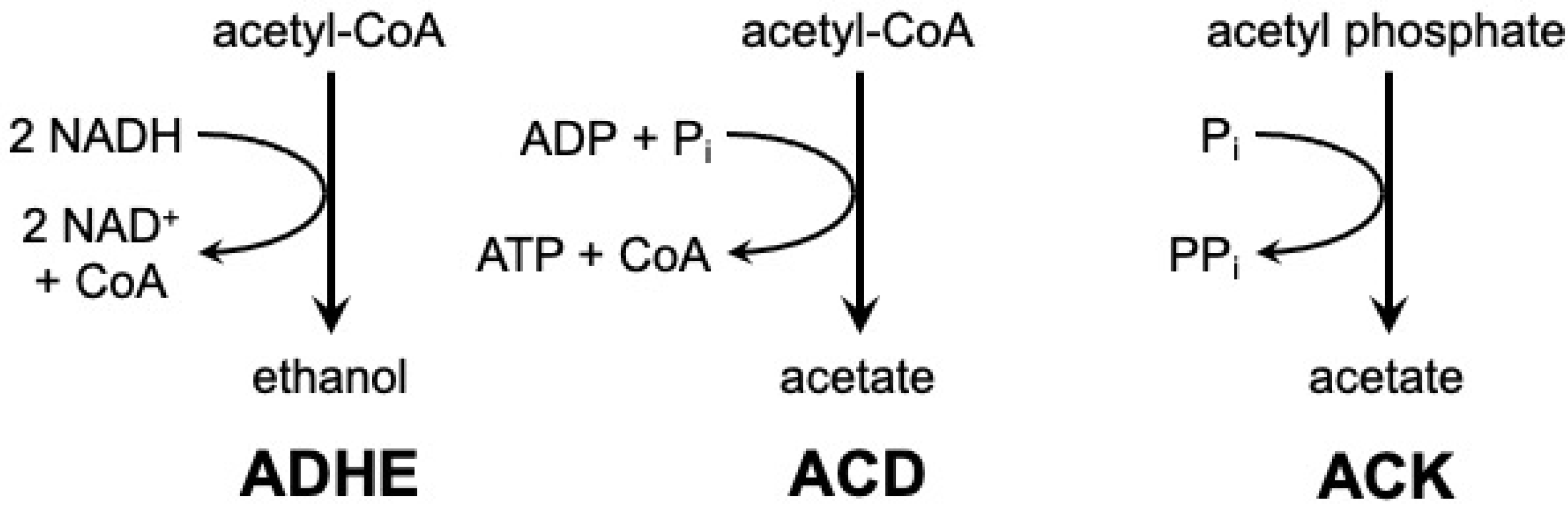
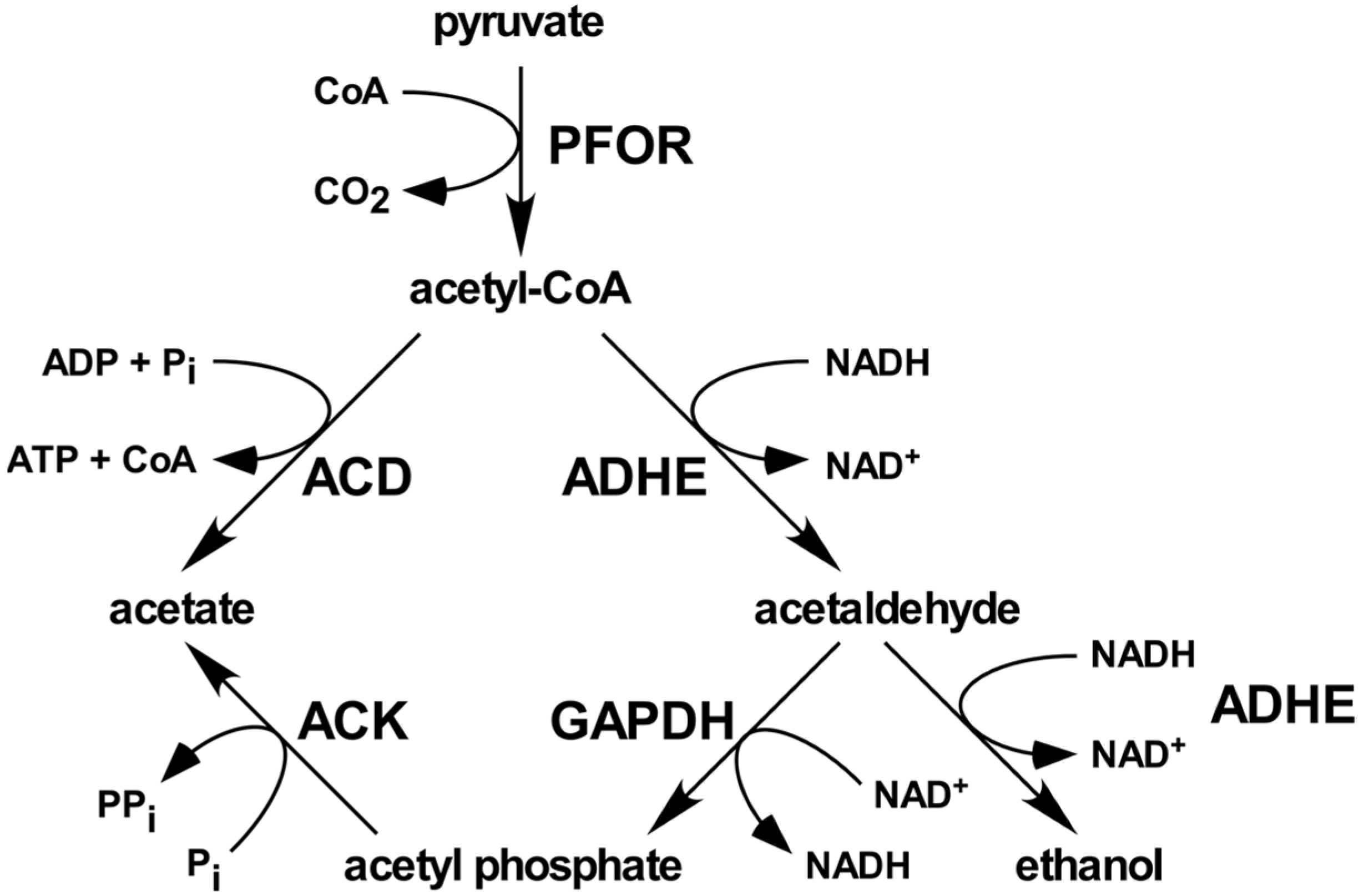
:1. Introduction
2. Results
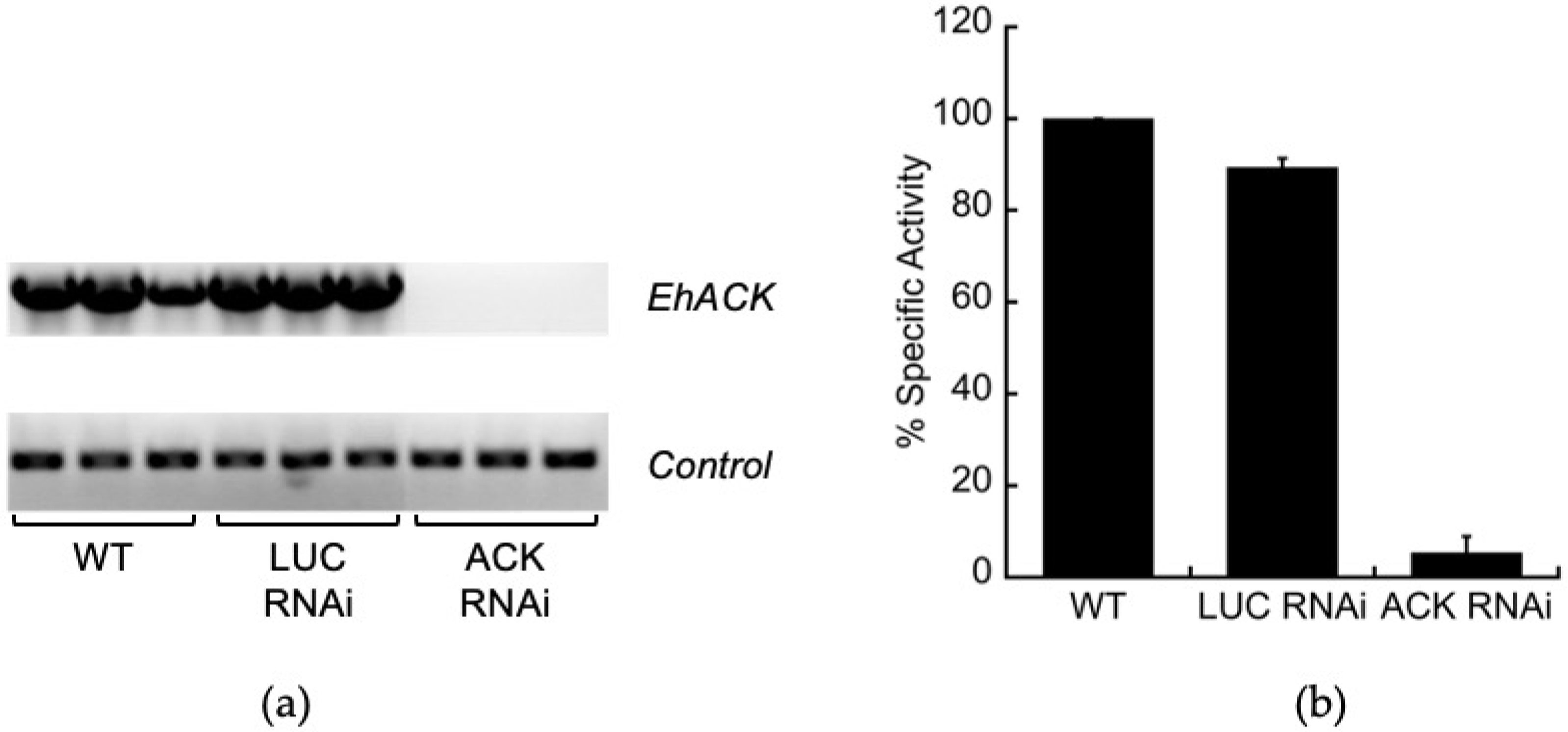
2.1. Trigger-Mediated ACK Gene Silencing
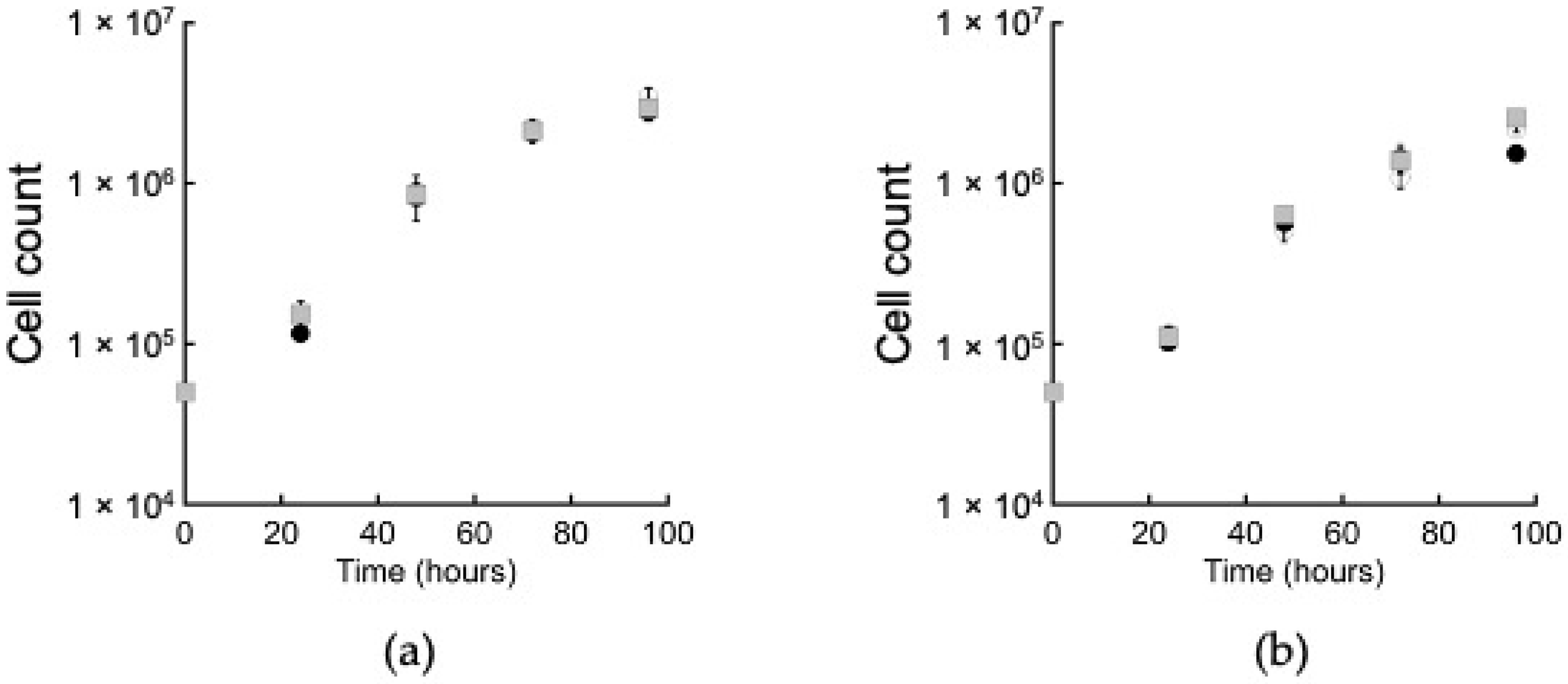
2.2. ACK Is Dispensable for Growth on Glucose
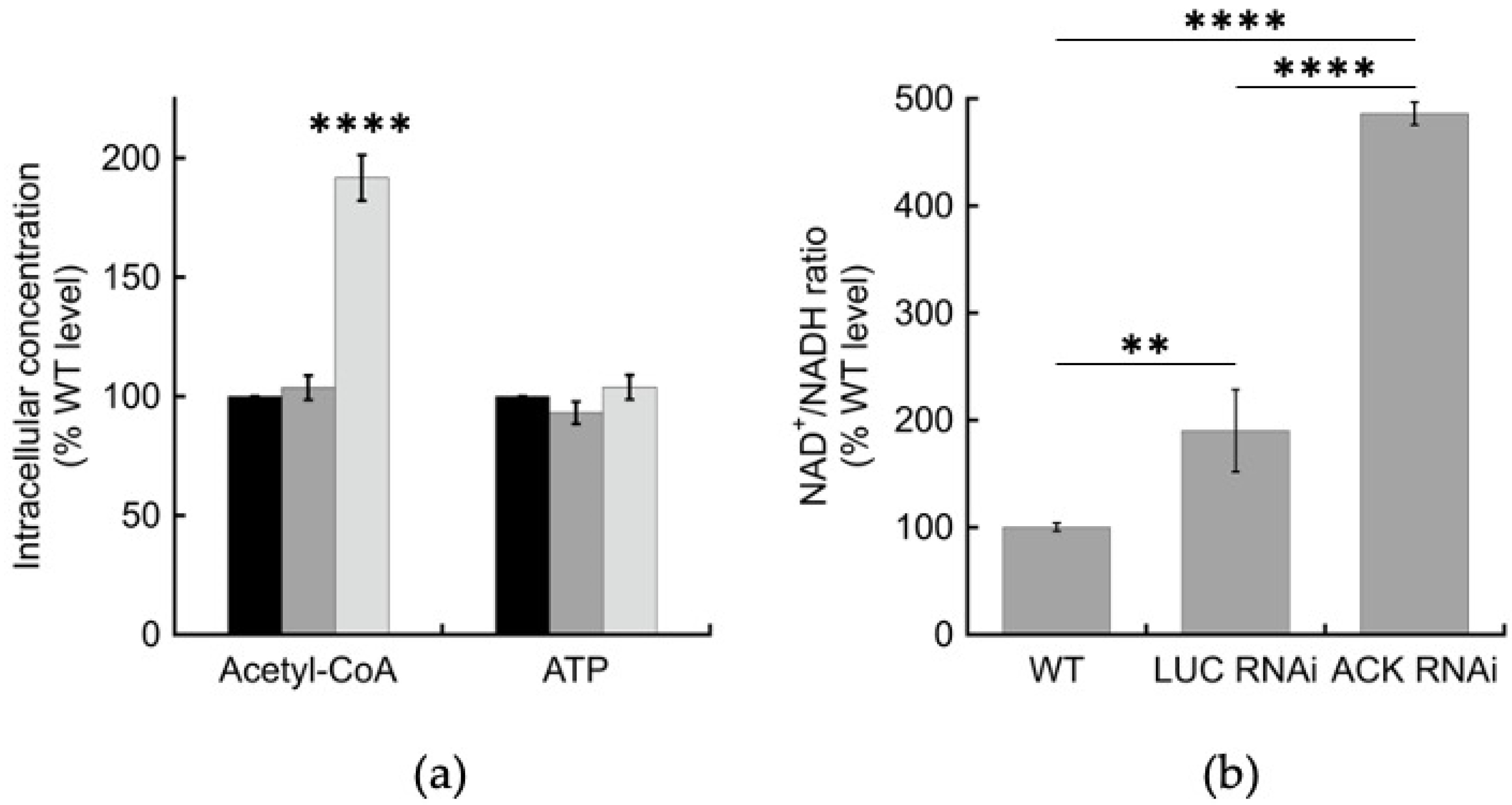
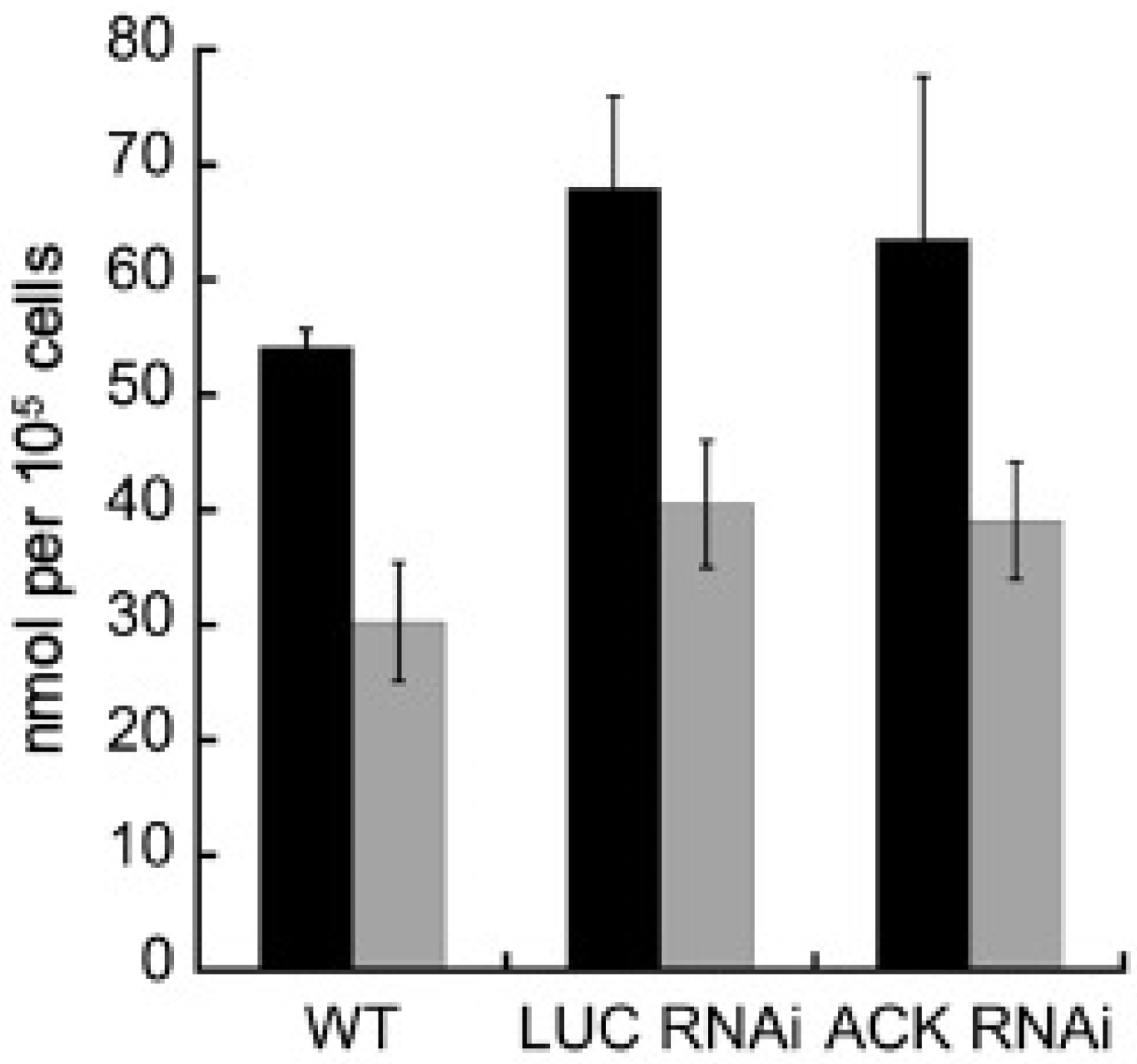
2.3. Intracellular but Not Extracellular Metabolite Levels Are Altered in the ACK RNAi Cell Line
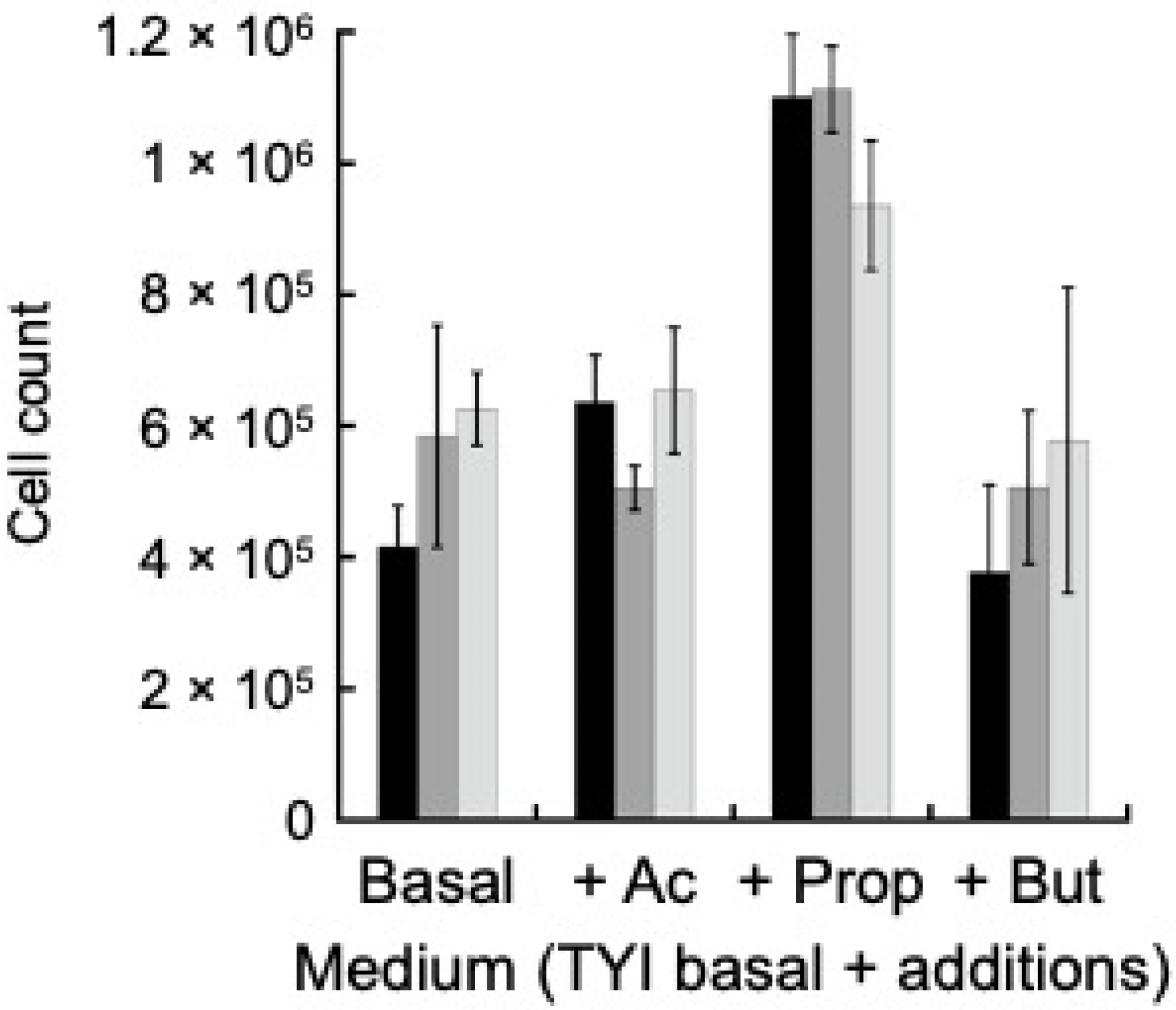
2.4. ACK Does Not Play a Role in Utilization of Short Chain Fatty Acids
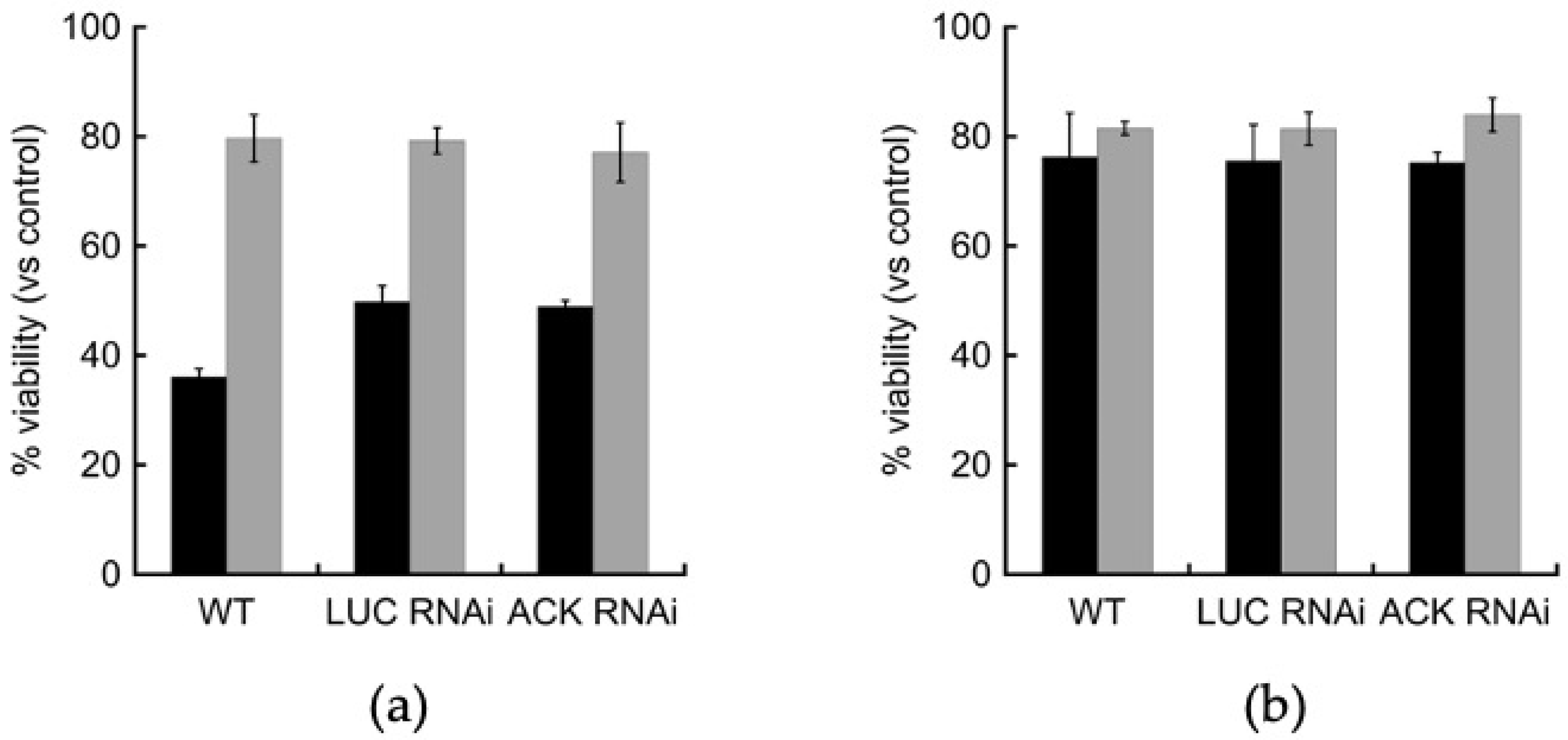
2.5. Oxidative and Nitrosative Stress Response Are Unaffected in the ACK RNAi Cell Line
2.6. EhGAPDH Displays Acetyl Phosphate-Forming Activity in the Presence of ACK
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines and Culture Conditions
4.3. Construct Cloning and Transfection
4.4. Reverse Transcriptase PCR (RT-PCR)
4.5. Enzyme Assays
4.6. Intracellular Metabolite Analysis
4.7. Extracellular Metabolite Analysis
4.8. Oxidative Stress and Nitrosative Stress Induction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization; Foodborne Disease Burden Epidemiology Reference Group. WHO Estimates of the Global Burden of Foodborne Diseases; World Health Organization: Geneva, Switzerland, 2015; p. 11. 254p. [Google Scholar]
- Baxt, L.A.; Singh, U. New insights into Entamoeba histolytica pathogenesis. Curr. Opin. Infect. Dis. 2008, 21, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, R.; Huston, C.D.; Hughes, M.; Houpt, E.; Petri, W.A., Jr. Amebiasis. N. Engl. J. Med. 2003, 348, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Stanley, S.L., Jr. Amoebiasis. Lancet 2003, 361, 1025–1034. [Google Scholar] [CrossRef]
- Anderson, I.J.; Loftus, B.J. Entamoeba histolytica: Observations on metabolism based on the genome sequence. Exp. Parasitol. 2005, 110, 173–177. [Google Scholar] [CrossRef]
- Clark, C.G.; Alsmark, U.C.; Tazreiter, M.; Saito-Nakano, Y.; Ali, V.; Marion, S.; Weber, C.; Mukherjee, C.; Bruchhaus, I.; Tannich, E.; et al. Structure and content of the Entamoeba histolytica genome. Adv. Parasitol. 2007, 65, 51–190. [Google Scholar]
- Reeves, R.E.; Serrano, R.; South, D.J. 6-phosphofructokinase (pyrophosphate). Properties of the enzyme from Entamoeba histolytica and its reaction mechanism. J. Biol. Chem. 1976, 251, 2958–2962. [Google Scholar] [CrossRef]
- Saavedra, E.; Encalada, R.; Pineda, E.; Jasso-Chavez, R.; Moreno-Sanchez, R. Glycolysis in Entamoeba histolytica. Biochemical characterization of recombinant glycolytic enzymes and flux control analysis. FEBS J. 2005, 272, 1767–1783. [Google Scholar] [CrossRef]
- Saavedra-Lira, E.; Perez-Montfort, R. Energy production in Entamoeba histolytica: New perspectives in rational drug design. Arch. Med. Res. 1996, 27, 257–264. [Google Scholar]
- Lo, H.S.; Reeves, R.E. Pyruvate-to-ethanol pathway in Entamoeba histolytica. Biochem. J. 1978, 171, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Montalvo, F.E.; Reeves, R.E.; Warren, L.G. Aerobic and anaerobic metabolism in Entamoeba histolytica. Exp. Parasitol. 1971, 30, 249–256. [Google Scholar] [CrossRef]
- Reeves, R.E.; Warren, L.G.; Susskind, B.; Lo, H.S. An energy-conserving pyruvate-to-acetate pathway in Entamoeba histolytica. Pyruvate synthase and a new acetate thiokinase. J. Biol. Chem. 1977, 252, 726–731. [Google Scholar] [CrossRef]
- Pineda, E.; Encalada, R.; Olivos-Garcia, A.; Nequiz, M.; Moreno-Sanchez, R.; Saavedra, E. The bifunctional aldehyde-alcohol dehydrogenase controls ethanol and acetate production in Entamoeba histolytica under aerobic conditions. FEBS Lett. 2013, 587, 178–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, E.; Encalada, R.; Rodriguez-Zavala, J.S.; Olivos-Garcia, A.; Moreno-Sanchez, R.; Saavedra, E. Pyruvate:ferredoxin oxidoreductase and bifunctional aldehyde-alcohol dehydrogenase are essential for energy metabolism under oxidative stress in Entamoeba histolytica. FEBS J. 2010, 277, 3382–3395. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, A.; Yan, L.; Zhang, Z.; Foster, L.; Clark, D.; Li, E.; Stanley, S.L., Jr. The bifunctional Entamoeba histolytica alcohol dehydrogenase 2 (EhADH2) protein is necessary for amebic growth and survival and requires an intact C-terminal domain for both alcohol dahydrogenase and acetaldehyde dehydrogenase activity. J. Biol. Chem. 2001, 276, 20136–20143. [Google Scholar] [CrossRef] [Green Version]
- Fowler, M.L.; Ingram-Smith, C.; Smith, K.S. Novel pyrophosphate-forming acetate kinase from the protist Entamoeba histolytica. Eukaryot. Cell 2012, 11, 1249–1256. [Google Scholar] [CrossRef] [Green Version]
- Reeves, R.E.; Guthrie, J.D. Acetate kinase (pyrophosphate). A fourth pyrophosphate-dependent kinase from Entamoeba histolytica. Biochem. Biophys. Res. Commun. 1975, 66, 1389–1395. [Google Scholar] [CrossRef]
- Jones, C.P.; Ingram-Smith, C. Biochemical and kinetic characterization of the recombinant ADP-forming acetyl coenzyme A synthetase from the amitochondriate protozoan Entamoeba histolytica. Eukaryot. Cell 2014, 13, 1530–1537. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, A.J. The acetate switch. Microbiol. Mol. Biol. Rev. 2005, 69, 12–50. [Google Scholar] [CrossRef] [Green Version]
- Terlesky, K.C.; Barber, M.J.; Aceti, D.J.; Ferry, J.G. EPR properties of the Ni-Fe-C center in an enzyme complex with carbon monoxide dehydrogenase activity from acetate-grown Methanosarcina thermophila. Evidence that acetyl-CoA is a physiological substrate. J. Biol. Chem. 1987, 262, 15392–15395. [Google Scholar] [CrossRef]
- Singh-Wissmann, K.; Ferry, J.G. Transcriptional regulation of the phosphotransacetylase-encoding and acetate kinase-encoding genes (pta and ack) from Methanosarcina thermophila. J. Bacteriol. 1995, 177, 1699–1702. [Google Scholar] [CrossRef] [Green Version]
- Ingram-Smith, C.; Martin, S.R.; Smith, K.S. Acetate kinase: Not just a bacterial enzyme. Trends Microbiol. 2006, 14, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Catalanotti, C.; D’Adamo, S.; Wittkopp, T.M.; Ingram-Smith, C.J.; Mackinder, L.; Miller, T.E.; Heuberger, A.L.; Peers, G.; Smith, K.S.; et al. Alternative acetate production pathways in Chlamydomonas reinhardtii during dark anoxia and the dominant role of chloroplasts in fermentative acetate production. Plant Cell 2014, 26, 4499–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harting, J.; Velick, S.F. Transfer reactions of acetyl phosphate catalyzed by glyceraldehyde-3-phosphate dehydrogenase. J. Biol. Chem. 1954, 207, 867–878. [Google Scholar] [CrossRef]
- Harting, J.; Velick, S.F. Acetyl phosphate formation catalyzed by glyceraldehyde-3-phosphate dehydrogenase. J. Biol. Chem. 1954, 207, 857–865. [Google Scholar] [CrossRef]
- Morf, L.; Pearson, R.J.; Wang, A.S.; Singh, U. Robust gene silencing mediated by antisense small RNAs in the pathogenic protist Entamoeba histolytica. Nucleic Acids Res. 2013, 41, 9424–9437. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Dang, T.; Ingram-Smith, C. Investigation of pyrophosphate versus ATP substrate selection in the Entamoeba histolytica acetate kinase. Sci. Rep. 2017, 7, 5912. [Google Scholar] [CrossRef] [Green Version]
- Aceti, D.J.; Ferry, J.G. Purification and characterization of acetate kinase from acetate-grown Methanosarcina thermophila. Evidence for regulation of synthesis. J. Biol. Chem. 1988, 263, 15444–15448. [Google Scholar] [CrossRef]
- Latimer, M.T.; Ferry, J.G. Cloning, sequence analysis, and hyperexpression of the genes encoding phosphotransacetylase and acetate kinase from Methanosarcina thermophila. J. Bacteriol. 1993, 175, 6822–6829. [Google Scholar] [CrossRef] [Green Version]
- Ingram-Smith, C.; Gorrell, A.; Lawrence, S.H.; Iyer, P.; Smith, K.; Ferry, J.G. Characterization of the acetate binding pocket in the Methanosarcina thermophila acetate kinase. J. Bacteriol. 2005, 187, 2386–2394. [Google Scholar] [CrossRef] [Green Version]
- Ingram-Smith, C.; Barber, R.D.; Ferry, J.G. The role of histidines in the acetate kinase from Methanosarcina thermophila. J. Biol. Chem. 2000, 275, 33765–33770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibeaux, R.; Weber, C.; Hon, C.C.; Dillies, M.A.; Ave, P.; Coppee, J.Y.; Labruyere, E.; Guillen, N. Identification of the virulence landscape essential for Entamoeba histolytica invasion of the human colon. PLoS Pathog. 2013, 9, e1003824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrenkaufer, G.M.; Haque, R.; Hackney, J.A.; Eichinger, D.J.; Singh, U. Identification of developmentally regulated genes in Entamoeba histolytica: Insights into mechanisms of stage conversion in a protozoan parasite. Cell Microbiol. 2007, 9, 1426–1444. [Google Scholar] [CrossRef] [PubMed]
- Hon, C.C.; Weber, C.; Sismeiro, O.; Proux, C.; Koutero, M.; Deloger, M.; Das, S.; Agrahari, M.; Dillies, M.A.; Jagla, B.; et al. Quantification of stochastic noise of splicing and polyadenylation in Entamoeba histolytica. Nucleic Acids Res. 2013, 41, 1936–1952. [Google Scholar] [CrossRef] [Green Version]
- Pineda, E.; Vazquez, C.; Encalada, R.; Nozaki, T.; Sato, E.; Hanadate, Y.; Nequiz, M.; Olivos-Garcia, A.; Moreno-Sanchez, R.; Saavedra, E. Roles of acetyl-CoA synthetase (ADP-forming) and acetate kinase (PPi-forming) in ATP and PPi supply in Entamoeba histolytica. Biochim. Biophys. Acta 2016, 1860, 1163–1172. [Google Scholar] [CrossRef]
- Vicente, J.B.; Ehrenkaufer, G.M.; Saraiva, L.M.; Teixeira, M.; Singh, U. Entamoeba histolytica modulates a complex repertoire of novel genes in response to oxidative and nitrosative stresses: Implications for amebic pathogenesis. Cell Microbiol. 2009, 11, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Diamond, L.S.; Harlow, D.R.; Cunnick, C.C. A new medium for the axenic cultivation of Entamoeba histolytica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 431–432. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21, A.3B.1–A.3B.2. [Google Scholar] [CrossRef]
- Hamann, L.; Nickel, R.; Tannich, E. Transfection and continuous expression of heterologous genes in the protozoan parasite Entamoeba histolytica. Proc. Natl. Acad. Sci. USA 1995, 92, 8975–8979. [Google Scholar] [CrossRef] [Green Version]
- Vines, R.R.; Purdy, J.E.; Ragland, B.D.; Samuelson, J.; Mann, B.J.; Petri, W.A., Jr. Stable episomal transfection of Entamoeba histolytica. Mol. Biochem. Parasitol. 1995, 71, 265–267. [Google Scholar] [CrossRef]
- Koushik, A.B.; Welter, B.H.; Rock, M.L.; Temesvari, L.A. A genomewide overexpression screen identifies genes involved in the phosphatidylinositol 3-kinase pathway in the human protozoan parasite Entamoeba histolytica. Eukaryot. Cell 2014, 13, 401–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of protein. Methods Enzymol. 1990, 182, 50–68. [Google Scholar] [PubMed]
- Neubauer, S.; Chu, D.B.; Marx, H.; Sauer, M.; Hann, S.; Koellensperger, G. LC-MS/MS-based analysis of coenzyme A and short-chain acyl-coenzyme A thioesters. Anal. Bioanal. Chem. 2015, 407, 6681–6688. [Google Scholar] [CrossRef]
- Rose, I.A.; Grunberg-Manago, M.; Korey, S.R.; Ochoa, S. Enzymatic phosphorylation of acetate. J. Biol. Chem. 1954, 211, 737–756. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specific Activity (nmol min−1 mg−1) 1 | ||
|---|---|---|
| Enzymes | No ADP | 10 mM ADP |
| GAPDH | ND | ND |
| GAPDH + EhACK | 14.1 ± 0.60 | 7.04 ± 0.90 |
| GAPDH + MtACK | 2.29 ± 0.16 | 128 ± 2.33 |
| Cloning Primers | |
|---|---|
| EhACK RNAi F | 5′ CTACCTAGGATGTCTAACGTACTAATATTCAACG |
| EhACK RNAi R | 5′ CTACTCGAGTTAAAACTGAAATAATTCTTTTCCTTTTTGTAA |
| RT-PCR primers | |
| EhACK RTPCR F | 5′ AGGGTAAATGTTACAGGAACAGA |
| EhACK RTPCR R | 5′ TGGTGCCACACAAACTTGAAC |
| ssrRNA F | 5′-AGGCGCGTAAATTACCCACTTTCG |
| ssRNA R | 5′-CACCAGACTTGCCCTCCAATTGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, T.; Angel, M.; Cho, J.; Nguyen, D.; Ingram-Smith, C. The Role of Acetate Kinase in the Human Parasite Entamoeba histolytica. Parasitologia 2022, 2, 147-159. https://doi.org/10.3390/parasitologia2020014
Dang T, Angel M, Cho J, Nguyen D, Ingram-Smith C. The Role of Acetate Kinase in the Human Parasite Entamoeba histolytica. Parasitologia. 2022; 2(2):147-159. https://doi.org/10.3390/parasitologia2020014
Chicago/Turabian StyleDang, Thanh, Matthew Angel, Jin Cho, Diana Nguyen, and Cheryl Ingram-Smith. 2022. "The Role of Acetate Kinase in the Human Parasite Entamoeba histolytica" Parasitologia 2, no. 2: 147-159. https://doi.org/10.3390/parasitologia2020014