1. Introduction
Babesiosis is a tick-borne disease caused by intraerythrocytic protozoan parasites of the genus
Babesia. In particular,
Babesia bovis infection causes a severe disease characterized by hemolytic anemia and high mortality in adult bovines [
1], whereas
B. microti invades and replicates in red blood cells (RBCs) of animals, including humans, to cause zoonotic babesiosis [
2]. Infection of
B. microti in mouse has been used as an experimental infection model to evaluate the effect of potential compounds for the treatment of bovine babesiosis [
3,
4]. The control strategies currently applied for bovine babesiosis include treatment of the infected animal, tick control using acaricide and live attenuated vaccine administration, in which emerging resistance to drugs and acaricides and lack of effective vaccines are the main obstacles [
1,
5].
Babesia spp., along with many other species (e.g.,
Plasmodium spp. and
Toxoplasma sp.), belong to the phylum
Apicomplexa. Most apicomplexan parasites have a non-photosynthetic plastid called the apicoplast, which is involved in some essential metabolic pathways such as the heme biosynthesis pathway, and its functions are considered to be potential targets for chemotherapy [
6,
7].
5-Aminolevulenic acid (ALA) is an endogenous, non-proteinogenic, naturally occurring amino acid found in diverse organisms such as bacteria, fungi, plants, and animals, and is a precursor of heme biosynthesis. It is commercially available as a component of cosmetics, dietary supplements, and pharmaceuticals for cancer diagnosis and therapy [
8,
9].
Amongst protozoan parasites, ALA has only been evaluated against
Plasmodium falciparum infection. ALA was recently reported to efficiently inhibit in vitro growth of the human malaria parasite
P. falciparum in the presence of ferrous ion (Fe
2+) [
10]. Reactive oxygen species (ROS) are produced by interaction between the ALA-derived heme intermediates and Fe
2+, and ROS can damage DNA and membranes of the apicoplast. Therefore, ALA was suggested to be a potential antimalarial treatment in humans [
10]. In addition, the combination of ALA and sodium ferrous citrate (SFC) as a source of ferrous ions was found to be beneficial in improving symptoms in malaria patients, which led to fast recovery, thus suggesting ALA/SFC as a promising composition for malaria chemotherapy [
9].
Accordingly, this study was conducted to evaluate the growth inhibitory effect of the ALA/SFC combination against B. bovis under in vitro culture conditions. In addition, the curative effect of orally administered ALA/SFC in mice against B. microti infection was investigated to evaluate the potential of ALA/SFC to offer symptomatic improvement of bovine babesiosis.
3. Discussion
The growth inhibitory effect of the ALA/SFC combination against
P. falciparum under in vitro culture condition [
10] and its curative potential against
P. berghei infection in mice [
9] have recently been reported. The apicoplast, which is involved in the heme biosynthesis pathway of malaria parasites, is essential for parasite development [
7], and ALA was suggested to be a potential antimalarial drug that targets the heme biosynthesis pathway in this organelle [
10]. Given that
Plasmodium and
Babesia are apicomplexan parasites that both have apicoplasts [
6], it could be hypothesized that ALA might have potential in the treatment of bovine babesiosis.
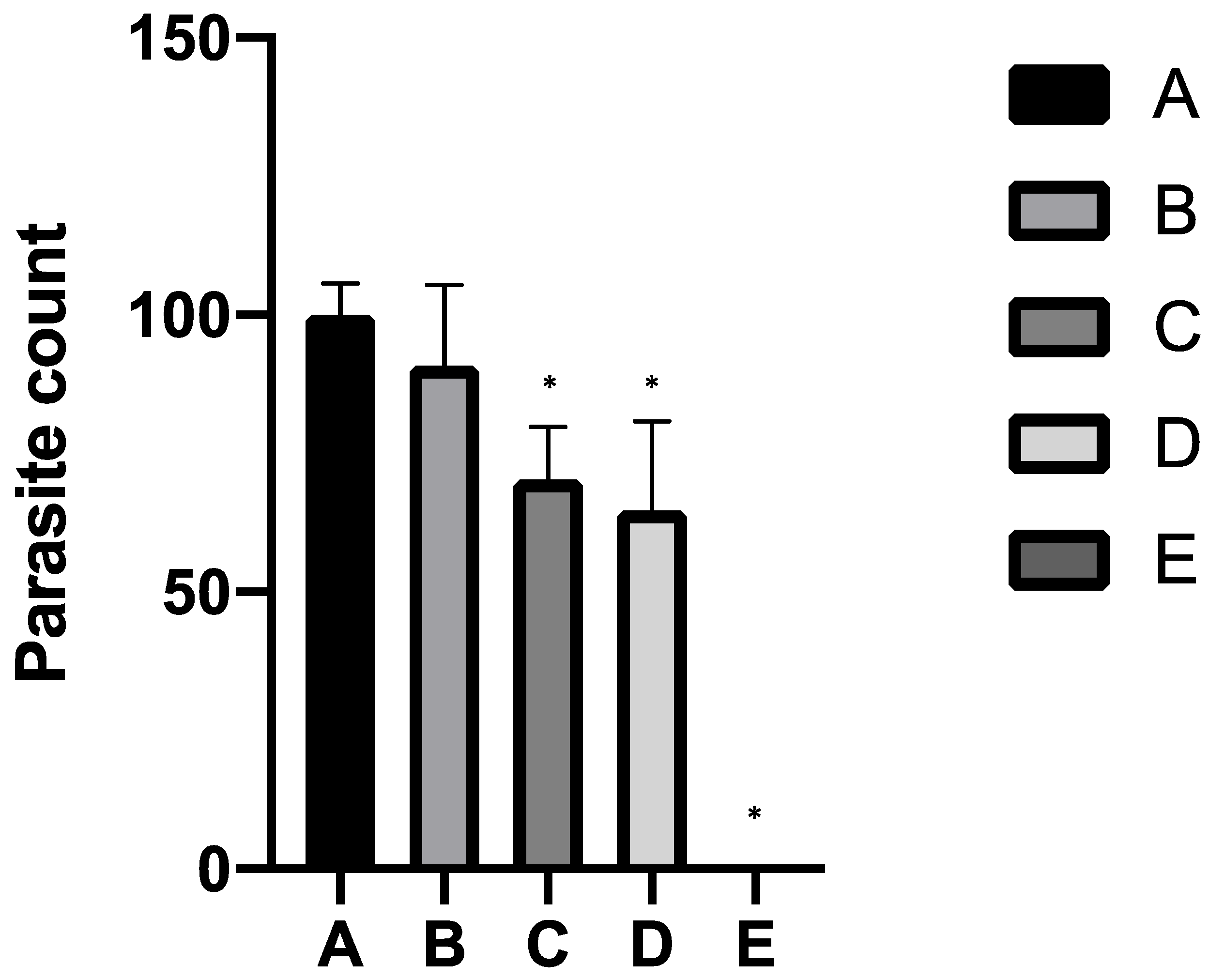
The growth inhibitory efficacy of the combination of ALA/SFC against
B. bovis was evaluated and found to significantly inhibit parasite growth under an in vitro culture condition. The addition of ALA/SFC 100/10 µM and ALA/SFC 500/10 µM into the culture medium inhibited parasite growth to 64% and 54%, respectively, of the control culture without the compounds. These concentrations of ALA/SFC were comparable to those showing >50% inhibitory effect against
P. falciparum under an in vitro culture condition [
10]. The mode of action proposed for ALA/SFC to inhibit the growth of
P. falciparum is that accumulation of intermediates such as coproporphyrin III and coexisting Fe
2+ that form the heme biosynthesis pathway produce ROS, which damages the DNA and membrane of the apicoplast. Analysis of the
B. bovis genome predicted that the heme biosynthesis pathway found in
P. falciparum was absent in this parasite species [
11]. However, the parasite genome contains the gene coding for delta-aminolevulinic acid dehydratase, which catalyzes the reaction producing porphobilinogen from ALA as the second step in the heme biosynthesis [
11,
12]. This step in
P. falciparum takes place in the apicoplast [
12].
Babesia parasites possess several heme-dependent enzymes that require heme as a prosthetic group for their catalytic activities; however, no transporter for heme salvage has been identified so far in the parasite genome [
11,
12]. Re-evaluation of the genome with appropriate prediction algorithms may provide additional information on the babesia heme biosynthesis pathway and may explain the mode of action of ALA/SFC in this parasite species.
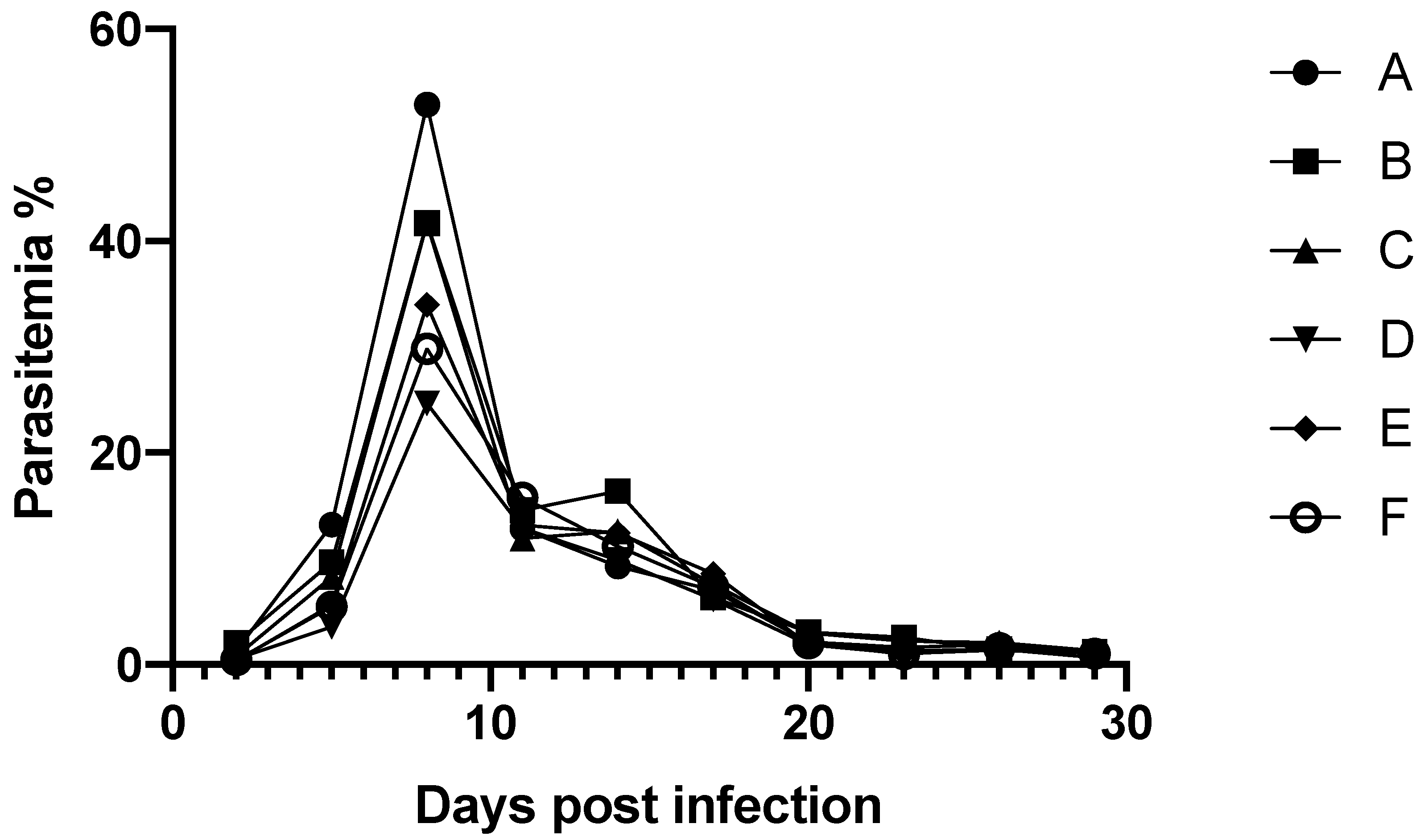
The curative effect of ALA/SFC oral administration in mice was evaluated in vivo with a
B. microti experimental infection model. Oral administration was chosen because the application method of ALA/SFC as a “feed additive” to animals such as dairy cows and beef cattle was expected. The safety of ALA/SFC as a dietary supplement has been reviewed [
8]. All administrations including either ALA or SFC alone or their combinations inhibited development of
B. microti in the mice blood circulation compared with that in the control (
Figure 2 and
Table 1). The oral administration of ALA/SFC 4/0.4 mg/kg showed a curative effect, and the dose was much lower than that showed curative effect against the mouse malaria infection (ALA/SFC 300/300 mg/kg) [
9]. ALA may have other modes of action against babesia parasites other than that against malaria parasites. The antiviral activity of ALA against feline infectious peritonitis virus with unidentified factors other than oxidative stress has been reported [
13]. The mice administered ALA alone showed significantly lower parasitemia than that of the control. The curative effect of ALA alone at a high dose (600 mg/kg) without SFC was also observed in mice infected with malaria, and it was explained by that the mice had ingested sufficient ferrous ion from their diet under the ordinal experimental condition [
9]. The curative effect of ALA alone in our experiment was most probably attributable to the same reason as the animals were ordinally fed with an experimental animal diet that contained sufficient ferrous ion (34.24 mg of ferrous ion/100 g of CLEA Rodent Diet CE-2; Japan CLEA Inc., Tokyo, Japan,
https://www.clea-japan.com/en/products/general_diet/item_d0030 (accessed on 15 October 2021).
The progression of parasitemia was accompanied by body weight loss in all of the infected mice. The body weight of the mice gradually decreased until the peak of parasitemia was reached and then increased thereafter. The body weight losses at the peak of parasitemia in the mice administered ALA/SFC 4/0.4 mg/kg and ALA/SFC 20/2 mg/kg were significantly improved compared with that of the control mice. It is worth mentioning that administration of SFC alone also significantly improved body weight loss in the mice at the peak of parasitemia and thereafter. The effect of ALA on improving milk protein content has been reported in dairy cows that were fed the compound as a “dietary supplement” [
14]. The effect of ALA/SFC administration on the improvement of body weight loss of infected animals is favorable to its application in dairy cows and beef cattle, although the reason for such effect with SFC alone is unclear.
RBC, HGB, and HCT decreased as a result of anemia as the parasitemia progressed. The mice administered ALA/SFC 4/0.4 mg/kg showed moderate decrease in these parameters at the peak of parasitemia compared with the control mice. This finding indicated that oral administration of ALA/SFC could moderate the progression of anemia, one of the most important clinical signs of bovine babesiosis [
1].
4. Materials and Methods
4.1. Chemicals
ALA and SFC were obtained from neopharma Japan Co., Ltd. (Tokyo, Japan). Hydrochloric acid (HCl) and dimethyl sulfoxide (DMSO) were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). DA was purchased from Sigma-Aldrich Japan Co., Ltd. (Tokyo, Japan). ALA was dissolved in ultrapure water (Direct-Q 3UV, Merck Millipore, Darmstadt, Germany) to make a 100 mM solution, and SFC was dissolved in 0.004 N HCl (final concentration) to make a 10 mM solution. Both were filtrated with a 0.2-µm filter (Dismic-25CS, Advantec Co., Ltd., Tokyo, Japan) for sterilization and diluted with the culture medium at the specified concentrations. DA was dissolved in DMSO at 10 mM and stored at 4 °C. For animal experimentation, ALA and SFC were dissolved in phosphate-buffered saline (PBS) containing 0.004 N HCl (final concentration) at the specified concentrations for oral administration. All drugs except 10 mM DA were freshly prepared on the day of use.
4.2. Parasite Culture and Growth Inhibition Assay
B. bovis Texas strain was maintained in culture using a microaerophilic stationary-phase culture system composed of bovine RBCs at 10% hematocrit and GIT medium (FUJIFILM Wako Pure Chemical Industries, Osaka, Japan) [
15]. Parasite cultures (1 mL) were incubated with different concentrations of ALA (100 and 500 µM) coupled with SFC (10 µM). The cultures with SFC (10 µM) alone and DA (1 µM) served as iron and anti-babesial drug controls, respectively. The non-treatment control contained DMSO at 0.01%. Each experimental group with initial parasitemia of 0.1% was cultured in triplicate using 24-well culture plate for 3 days. Culture medium with drug was changed daily, and the number of parasitized RBCs per 1000 RBCs was counted in triplicate on thin smears which had been prepared on the third day of incubation.
4.3. Animals
Six-week-old female BALB/c mice (body weight 19–23 g) were used. The animals were maintained in the animal facilities at the National Research Center for Protozoan Diseases (NRCPD).
4.4. Parasite Infection and Animal Experimentations
The Munich strain of
B. microti, which has been maintained at the NRCPD, was used [
4]. The parasite-infected RBCs (1 × 10
7) were injected intraperitoneally in each mouse included in the experiment. Five mice were used for each experimental group. One day before infection, body weight and blood parameters (RBC, HGB, and HCT) of each mouse were measured with CellTaq α (Nihon Kohden Corporation, Tokyo, Japan). These parameters in each mouse were then measured every 3 days after infection for 30 days until the end of the experiment. The parasitemia of
B. microti was monitored every 3 days from 2 days after infection until the end of the experiment by counting the number of parasitized RBCs per 1000 RBCs in triplicate on Giemsa-stained thin blood smears. Administration of the compounds was started on the day of infection with a single oral dose (200 µL) of the different combinations (
Table 1). The administration continued daily for 30 days until the end of the experiment.
The animal experimental protocols were performed with permission from and in accordance with the standards for animal experimentation of Obihiro University of Agriculture and Veterinary Medicine, Obihiro, Hokkaido Japan (Approval number of Animal exp. 20–33).
4.5. Statistical Analysis
In vitro parasite count, parasitemia, body weight, and blood parameters are expressed as the mean + SD, and values were compared between the control (group A) and the other groups using one-way ANOVA followed by Dunnett’s multiple comparisons test for the in vitro and in vivo studies. Differences were considered significant when the P value was less than 0.05. All tests were performed using GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA, USA;
www.graphpad.com (accessed on 15 October 2021).



 ,
, 




{kind=link}
{kind=link}