
Two New Species of Pristimantis (Anura: Strabomantidae) from Amazonas Department in Northeastern Peru



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics and Field Research
2.2. Morphology
2.3. Bioacoustics
2.4. Species Delimitation
2.5. Nomenclatural Act
3. Results
3.1. Systematics
3.1.1. Holotype
3.1.2. Paratypes
3.1.3. Diagnosis
3.1.4. Comparison with Other Species
3.1.5. Description of the Holotype


3.1.6. Intraspecific Variation

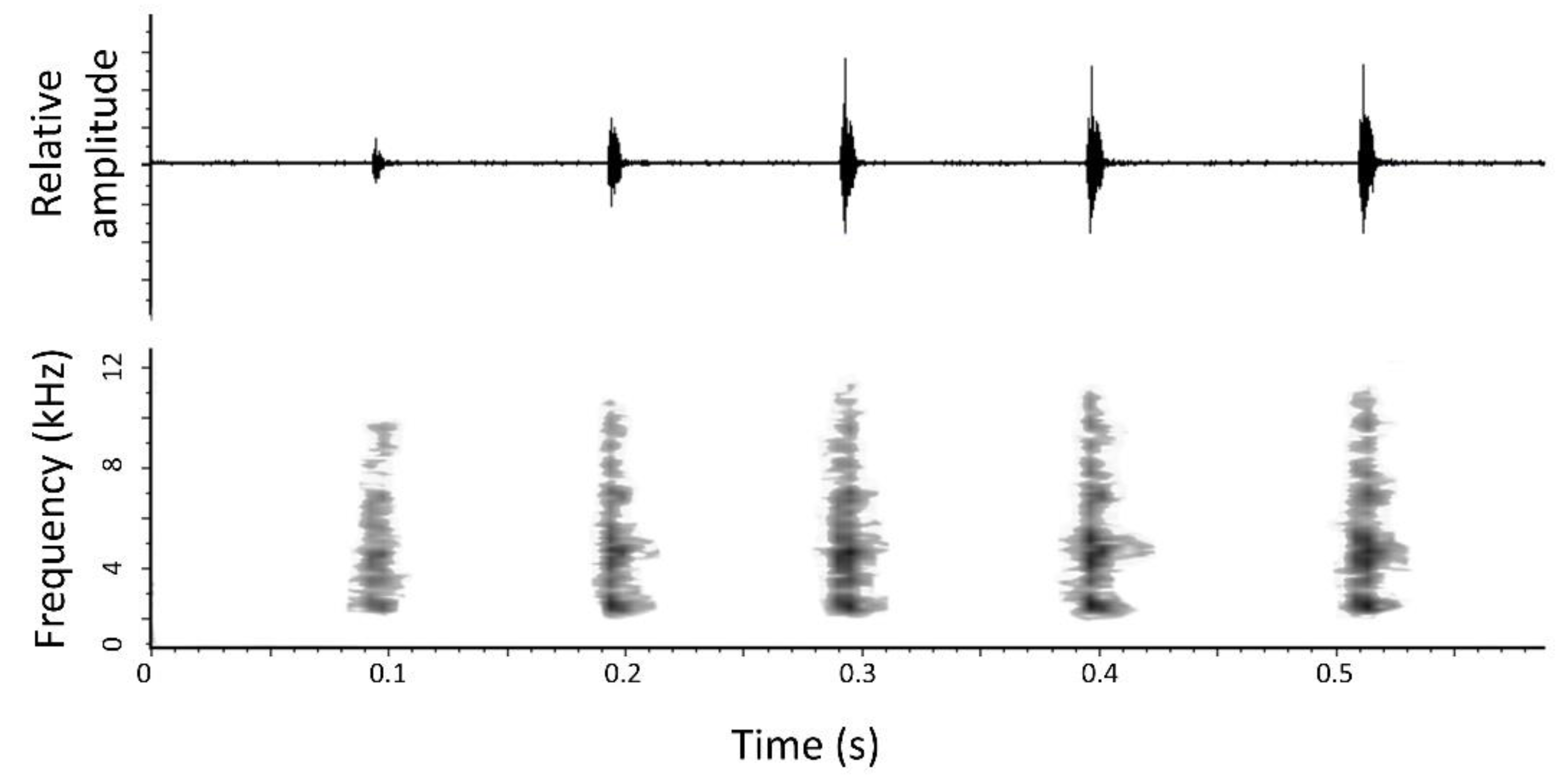
3.1.7. Advertisement Call

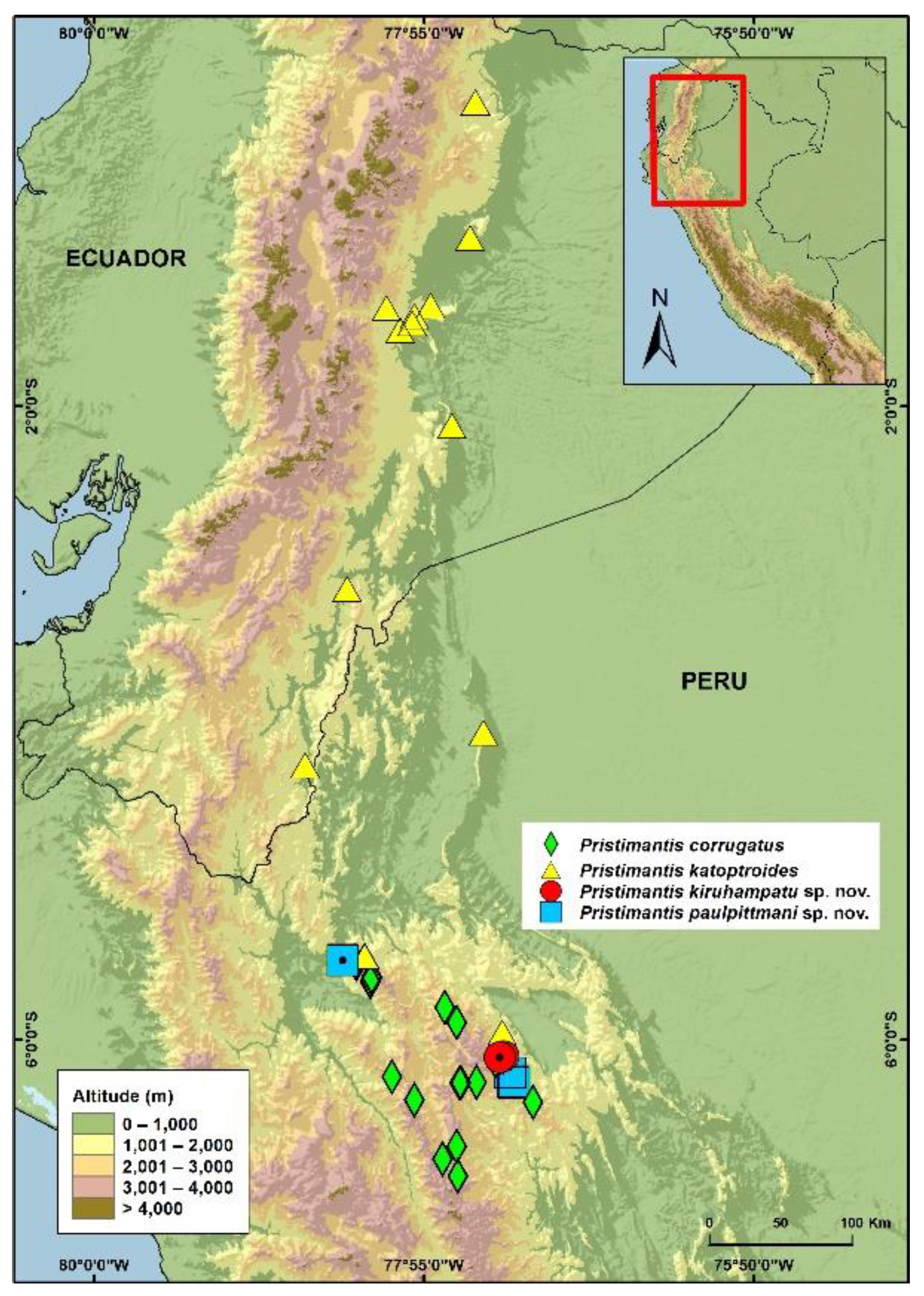
3.1.8. Distribution and Ecology
3.1.9. Etymology
3.2. Systematics
3.2.1. Holotype
3.2.2. Paratypes
3.2.3. Diagnosis
3.2.4. Comparison with Other Species
3.2.5. Description of the Holotype


3.2.6. Intraspecific Variation

3.2.7. Distribution and Natural History
3.2.8. Etymology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- AmphibiaWeb. AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 22 June 2021).
- Duellman, W.E.; Lehr, E. Terrestrial Breeding Frogs (Strabomantidae) in Peru; Natur und Tier-Verlag GmbH: Münster, Germany, 2009; p. 382. [Google Scholar]
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.1. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 10 July 2021).
- Hedges, S.B.; Duellman, W.E.; Heinicke, M.P. New World direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 2008, 1737, 1–182. [Google Scholar] [CrossRef] [Green Version]
- Brito-Zapata, D.; Reyes-Puig, C. A new species of terrestrial-breeding frog Pristimantis (Anura: Strabomantidae) from the Cordillera del Cóndor, Zamora Chinchipe, Ecuador. Neotrop. Biodivers. 2021, 7, 213–222. [Google Scholar] [CrossRef]
- Chávez, G.; García-Ayachi, L.A.; Catenazzi, A. Beauty is in the eye of the beholder: Cruciform eye reveals new species of direct-developing frog (Strabomantidae, Pristimantis) in the Amazonian Andes. Evol. Syst. 2021, 5, 81–92. [Google Scholar] [CrossRef]
- Navarrete, M.J.; Venegas, P.J.; Ron, S.R. Two new species of frogs of the genus Pristimantis from Llanganates National Park in Ecuador with comments on the regional diversity of Ecuadorian Pristimantis (Anura, Craugastoridae). ZooKeys 2016, 593, 139–162. [Google Scholar] [CrossRef]
- Páez, N.B.; Ron, S.R. Systematics of Huicundomantis, a new subgenus of Pristimantis (Anura, Strabomantidae) with extraordinary cryptic diversity and eleven new species. ZooKeys 2019, 868, 1. [Google Scholar] [CrossRef] [Green Version]
- Beaupre, S.J.; Jacobson, E.R.; Lillywhite, H.B.; Zamudio, K. Guidelines for Use of Live Amphibians and Reptiles in Field and Laboratory Research, 2nd ed.; The American Society of Ichthyologists and Herpetologist: Lawrence, KS, USA, 2004; p. 43. [Google Scholar]
- Scott, N.J. Complete species inventories. In Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.C., Foster, M.S., Eds.; Smithsonian Institution Press: Washinton, DC, USA, 1994; pp. 78–84. [Google Scholar]
- Lynch, J.D.; Duellman, W.E. Frogs of the Genus Eleutherodactylus (Leptodactylidae) in Western Ecuador: Systematic, Ecology, and Biogeography; Natural History Museum, The University of Kansas: Lawrence, KS, USA, 1997; p. 236. [Google Scholar]
- de Queiroz, K. The general lineage concept of species, species criteria, and the process of speciation. In Endless Forms: Species and Speciation; Howard, D.J., Berlocher, S.H., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 57–75. [Google Scholar]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.R.; Kluge, A.G. A consideration of epistomology in Systematic Biology, with special reference to species. Cladistics 1994, 10, 259–294. [Google Scholar] [CrossRef]
- Beraún, A.M.; Durán, V.; Álvarez, S.C.; Venegas, P.J. Distribution extension and an updated map for Pristimantis corrugatus, P. schultei and P. wagteri in northern Peru (Amphibia, Anura, Craugastoridae). Herpetol. Notes 2014, 7, 281–285. [Google Scholar]
- Cusi, J.C.; Barboza, A.C.; Vredenburg, V.; von May, R. A new locality, range extension and record of Batrachochytrium dendrobatidis in the endangered terrestrial breeding frog Pristimantis katoptroides Flores, 1988 (Anura: Craugastoridae) in Peru. Check List 2015, 11, 1608. [Google Scholar] [CrossRef]
- Brito, J.; Batallas, D.; Yánez-Muñoz, M.H. Ranas terrestres Pristimantis (Anura: Craugastoridae) de los bosques montanos del río Upano, Ecuador: Lista anotada, patrones de diversidad y descripción de cuatro especies nuevas. Neotrop. Biodivers. 2017, 3, 125–156. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.D.; Duellman, W.E. The Eleutherodactylus of the Amazonian slopes of the ecuadorian Andes (Anura: Leptodactylidae). Misc. Pub. Nat. Hist. Mus. Univ. Kan. 1980, 69, 1–86. [Google Scholar]
- Urgilés, V.L.; Sánchez-Nivicela, J.C.; Nieves, C.; Yánez-Muñoz, M.H. Ranas terrestres en los ecosistemas surandinos de Ecuador I: Dos nuevas especies de Pristimantis (Anura: Craugastoridae) de la ladera oriental. ACI Av. Cienc. Ing. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D'Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Duellman, W.E.; Pramuk, J.B. Frogs of the genus Eleutherodactylus (Anura: Leptodactylidae) in the Andes of Northern Peru. Nat. Hist. Mus. Univ. Kans. 1999, 13, 1–78. [Google Scholar] [CrossRef]
- Chaparro, J.C.; Motta, A.P.; Gutierrez, R.C.; Padial, J.M. A new species of Pristimantis (Anura: Strabomantidae) from Andean cloud forests of northern Peru. Zootaxa 2012, 3192, 39–48. [Google Scholar] [CrossRef]
- Mueses-Cisneros, J.J. Two new species of the genus Eleutherodactylus (Anura: Brachycephalidae) from Valle de Sibundoy, Putumayo, Colombia. Zootaxa 2007, 1498, 35–43. [Google Scholar] [CrossRef]
- Reyes-Puig, J.P.; Yánez-Muñoz, M.H. Una nueva especie de Pristimantis (Anura: Craugastoridae) del corredor ecológico llangantes-sangay, Andes de Ecuador. Papéis Avulsos Zool. 2012, 52, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Yánez-Muñoz, M.H.; Bejarano-Muñoz, P.; Brito, J.; Batallas, D. Ranas terrestres de los Andes Surorientales de Ecuador II: Una nueva especie de Pristimantis verde espinosa de los bosques montanos del Parque Nacional Sangay (Anura: Craugastoridae). ACI Av. Cienc. Ing. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Hutter, C.R.; Guayasamin, J.M. Cryptic diversity concealed in the Andean cloud forests: Two new species of rainfrogs (Pristimantis) uncovered by molecular and bioacoustic data. Neotrop. Biodivers. 2015, 1, 36–59. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Andrade, H.M.; Rojas-Soto, O.R.; Valencia, J.H.; Espinosa de los Monteros, A.; Morrone, J.J.; Ron, S.R.; Cannatella, D.C. Insights from integrative systematics reveal cryptic diversity in Pristimantis frogs (Anura: Craugastoridae) from the Upper Amazon Basin. PLoS ONE 2015, 10, e0143392. [Google Scholar] [CrossRef] [Green Version]
- Carrión-Olmedo, J.C.; Ron, S.R. A new cryptic species of the Pristimantis lacrimosus group (Anura, Strabomantidae) from the eastern slopes of the Ecuadorian Andes. Evol. Syst. 2021, 5, 151. [Google Scholar] [CrossRef]
- Ron, S.R.; Carrión, J.; Caminer, M.A.; Sagredo, Y.; Navarrete, M.J.; Ortega, J.A.; Varela-Jaramillo, A.; Maldonado-Castro, G.A.; Terán, C. Three new species of frogs of the genus Pristimantis (Anura, Strabomantidae) with a redefinition of the P. lacrimosus species group. ZooKeys 2020, 993, 121. [Google Scholar] [CrossRef] [PubMed]
- Lehr, E.; Gregory, C.; Catenazzi, A. A new species of Pristimantis (Amphibia: Anura: Strabomantidae) from the Rio Abiseo National Park, Peru. Zootaxa 2013, 3731, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Lehr, E.; Lyu, S.; Catenazzi, A. A new, critically endangered species of Pristimantis (Amphibia: Anura: Strabomantidae) from a mining area in the Cordillera Occidental of northern Peru (Región Cajamarca). Salamandra 2021, 57, 15–26. [Google Scholar]
- Venegas, P.J.; Duellman, W.E. Two syntopic new species of the Pristimantis orestes Group (Anura: Strabomantidae) from Northwestern Peru. Zootaxa 2012, 3249, 47–59. [Google Scholar] [CrossRef]
- Duellman, W.E.; Wild, E.R. Anuran amphibians from the Cordillera de Huancabamba, northern Peru: Systematics, ecology, and biogeography. Mus. Nat. Hist. Univ. Kans 1993, 157, 1–53. [Google Scholar]
- Lehr, E.; Aguilar, C.; Siu-Ting, K.; Carlos Jordán, J. Three new species of Pristimantis (Anura: Leptodactylidae) from the Cordillera de Huancabamba in northern Peru. Herpetologica 2007, 63, 519–536. [Google Scholar] [CrossRef]
- Lehr, E. New eleutherodactyline frogs (Leptodactylidae: Pristimantis, Phrynopus) from Peru. Bull. Mus. Comp. Zool. 2007, 159, 145–178. [Google Scholar] [CrossRef]
- Dillon, M.O.; Sagástegui A., A.; Sánchez, I.; Llatas Q., S.; Hensold, N. Floristic inventory and biogeographic analysis of montane forests in northwestern Peru. In Biodiversity and Conservation of Neotropical Montane Forest; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; The New York Botanical Garden Press: New York, NY, USA, 1995; pp. 251–270. [Google Scholar]
- Venegas, P.J. A new species of Eleutherodactylus (Anura: Leptodactylidae) from the Cordillera Central in Northern Peru. J. Herpetol. 2007, 41, 394–400. [Google Scholar] [CrossRef]
- Weigend, M.; Rodríguez, E.F.; Arana, C. The relict forests of northwest Peru and southwest Ecuador. Rev. Peru. Biol. 2005, 12, 185–194. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-1. Available online: https://www.iucnredlist.org (accessed on 7 July 2021).
- IUCN_SSC_Amphibian_Specialist_Group. Pristimantis rhodostichus. The IUCN Red List of Threatened Species 2018: E.T56912A89209326. 2018. Available online: https://doi.org/10.2305/IUCN.UK.2018-1.RLTS.T136162A89221263.en (accessed on 7 July 2021).
- Páez-Rosales, N.; Varela-Jaramillo, A. Pristimantis rhodostichus. In Anfibios del Ecuador; Version 2019.0; Ron, S.R., Merino-Viteri, A., Ortiz, D.A., Eds.; Museo de Zoología, Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2018. [Google Scholar]
- Varela-Jaramillo, A. Pristimantis muscosus. In Anfibios del Ecuador; Version, 2019.0; Ron, S.R., Merino-Viteri, A., Ortiz, D.A., Eds.; Museo de Zoología, Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2019. [Google Scholar]
- Yánez-Muñoz, M.H.; Toral-Contreras, E.; Meza-Ramos, P.A.; Reyes-Puig, J.P.; Bejarano-Muñoz, E.P.; Mueses-Cisneros, J.J.; Paucar, D. New country records for five species of Pristimantis Jiménez de la Espada, 1870 from Ecuador. Check List 2012, 8, 286–290. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pristimantis kiruhampatu sp. nov. | Pristimantis paulpittmani sp. nov. | |||
|---|---|---|---|---|
| Females (n = 4) | Males (n = 14) | Females (n = 10) | Males (n = 8) | |
| SVL | 19.5–23.5 (21.8 ± 1.7) | 13.0–18.3 (14.7 ± 1.8) | 18.1–23.8 (21.7 ± 1.7) | 15.2–17.4 (16.1 ± 0.7) |
| TL | 10.8–11.9 (11.4 ± 0.5) | 6.7–10.6 (8.3 ± 1.1) | 10–12.6 (11.2 ± 0.6) | 8.1–9.5 (8.7 ± 0.4) |
| FL | 9.7–11.2 (10.7 ± 0.7) | 5.3–9.3 (7.0 ± 1.2) | 8.3–10.9 (9.7 ± 0.8) | 6.7–7.7 (7.2 ± 0.4) |
| HL | 6.9–8.1 (7.6 ± 0.5) | 4.9–6.6 (5.5 ± 0.6) | 6.1–8.2 (7.4 ± 0.6) | 5.2–6.3 (5.6 ± 0.3) |
| HW | 7.8–8.7 (8.3 ± 0.4) | 4.8–7.5 (5.6 ± 0.8) | 7.4–9.7 (8.7 ± 0.6) | 6.1–6.8 (6.4 ± 0.3) |
| ED | 2.1–2.7 (2.4 ± 0.3) | 1.2–2.6 (2.0 ± 0.3) | 2.2–3.3 (2.6 ± 0.3) | 2.0–2.8 (2.3 ± 0.2) |
| IOD | 2.5–2.8 (2.7 ± 0.1) | 0.8–2.2 (1.7 ± 0.4) | 2.1–3.3 (2.6 ± 0.3) | 1.9–2.3 (2.0 ± 0.1) |
| EW | 2.0–2.5 (2.3 ± 0.2) | 0.5–2.0 (1.5 ± 0.4) | 1.5–2.3 (1.9 ± 0.2) | 1.4–2.0 (1.7 ± 0.2) |
| IND | 1.9–2.1 (2.1 ± 0.1) | 0.3–1.7 (1.3 ± 0.4) | 1.9–2.4 (2.2 ± 0.1) | 1.6–1.8 (1.7 ± 0.1) |
| E-N | 2.3–2.7 (2.4 ± 0.2) | 1.1–2.1 (1.5 ± 0.3) | 2.1–2.8 (2.5 ± 0.2) | 1.6–2 (1.8 ± 0.1) |
| TL/SVL | 0.50–0.55 (0.52 ± 0.03) | 0.52–0.60 (0.56 ± 0.03) | 0.46–0.55 (0.52 ± 0.02) | 0.51–0.57 (0.54 ± 0.03) |
| FL/SVL | 0.46–0.51 (0.49 ± 0.02) | 0.40–0.51 (0.47 ± 0.03) | 0.41–0.49 (0.45 ± 0.02) | 0.44–0.46 (0.45 ± 0.01) |
| HL/SVL | 0.32–0.36 (0.35 ± 0.02) | 0.32–0.40 (0.37 ± 0.02) | 0.33–0.36 (0.34 ± 0.01) | 0.34–0.36 (0.35 ± 0.01) |
| HW/SVL | 0.35–0.40 (0.38 ± 0.02) | 0.36–0.41 (0.38 ± 0.02) | 0.38–0.43 (0.40 ± 0.02) | 0.38–0.42 (0.40 ± 0.01) |
| HW/HL | 1.05–1.14 (1.10 ± 0.04) | 0.92–1.15 (1.02 ± 0.07) | 1.12–1.23 (1.18 ± 0.04) | 1.07–1.24 (1.14 ± 0.05) |
| E–N/ED | 0.85–1.29 (1.05 ± 0.20) | 0.50–1.00 (0.74 ± 0.17) | 0.79–1.09 (0.96 ± 0.12) | 0.63–0.92 (0.78 ± 0.09) |
| EW/IOD | 0.74–0.96 (0.84 ± 0.10) | 0.63–0.95 (0.84 ± 0.10) | 0.60–0.97 (0.74 ± 0.11) | 0.76–0.90 (0.83 ± 0.04) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venegas, P.J.; García-Ayachi, L.A.; Catenazzi, A. Two New Species of Pristimantis (Anura: Strabomantidae) from Amazonas Department in Northeastern Peru. Taxonomy 2022, 2, 20-40. https://doi.org/10.3390/taxonomy2010002
Venegas PJ, García-Ayachi LA, Catenazzi A. Two New Species of Pristimantis (Anura: Strabomantidae) from Amazonas Department in Northeastern Peru. Taxonomy. 2022; 2(1):20-40. https://doi.org/10.3390/taxonomy2010002
Chicago/Turabian StyleVenegas, Pablo J., Luis A. García-Ayachi, and Alessandro Catenazzi. 2022. "Two New Species of Pristimantis (Anura: Strabomantidae) from Amazonas Department in Northeastern Peru" Taxonomy 2, no. 1: 20-40. https://doi.org/10.3390/taxonomy2010002