
Production, Kinetic/Thermodynamic Study, and Evaluation of the Influence of Static Magnetic Field on Kinetic Parameters of β-Fructofuranosidase from Aspergillus tamarii Kita UCP 1279 Produced by Solid-State Fermentation

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Maintenance and Preparation of Spore Suspension
2.2. Solid-State Fermentation Conditions for FFase Production
2.3. FFase Hydrolytic Activity
2.4. Optimum Reaction PH and Temperature of Sucrose Hydrolysis Catalyzed by FFase
2.5. Kinetics and Thermodynamics of Thermal Denaturation of FFase
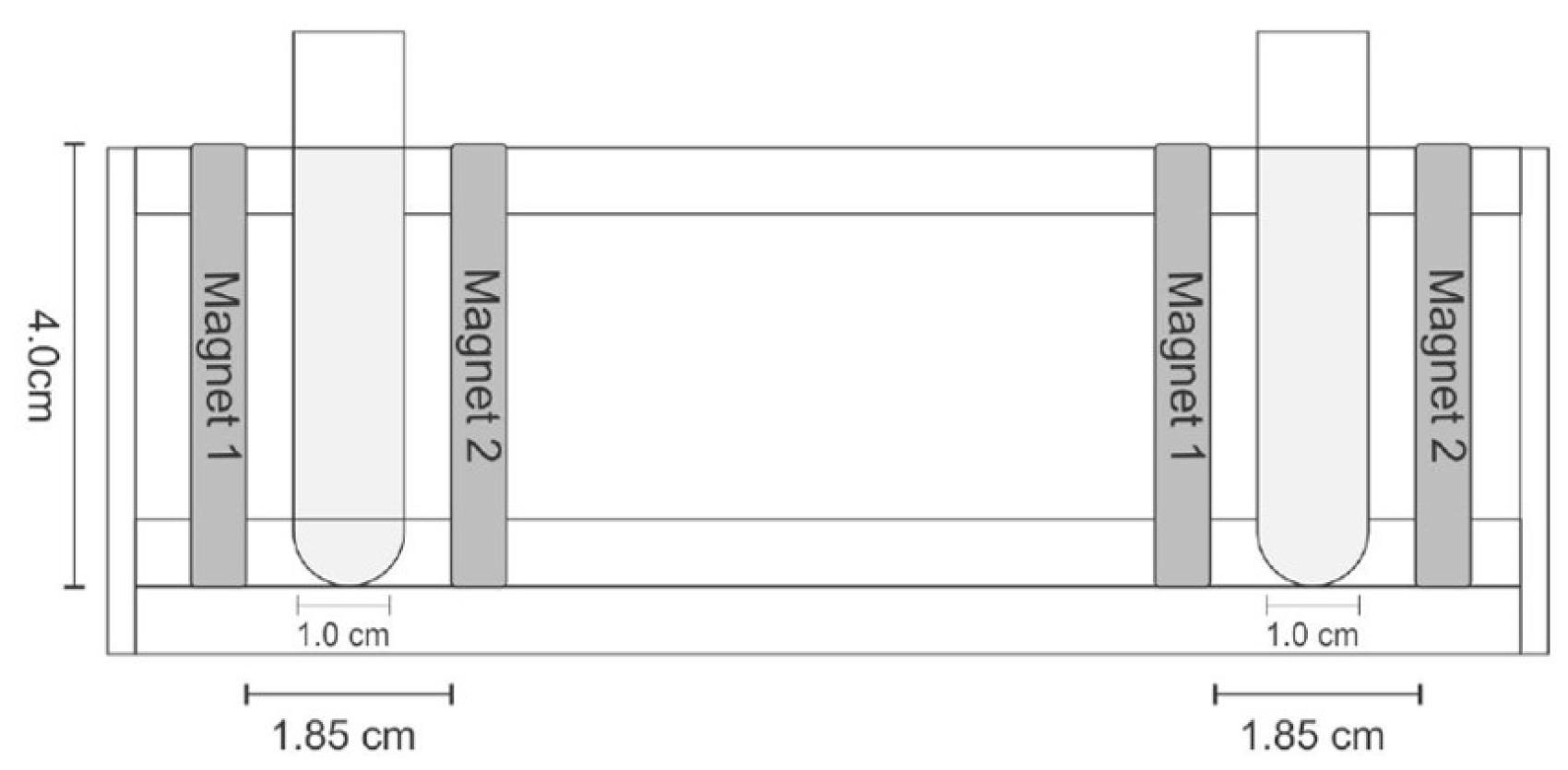
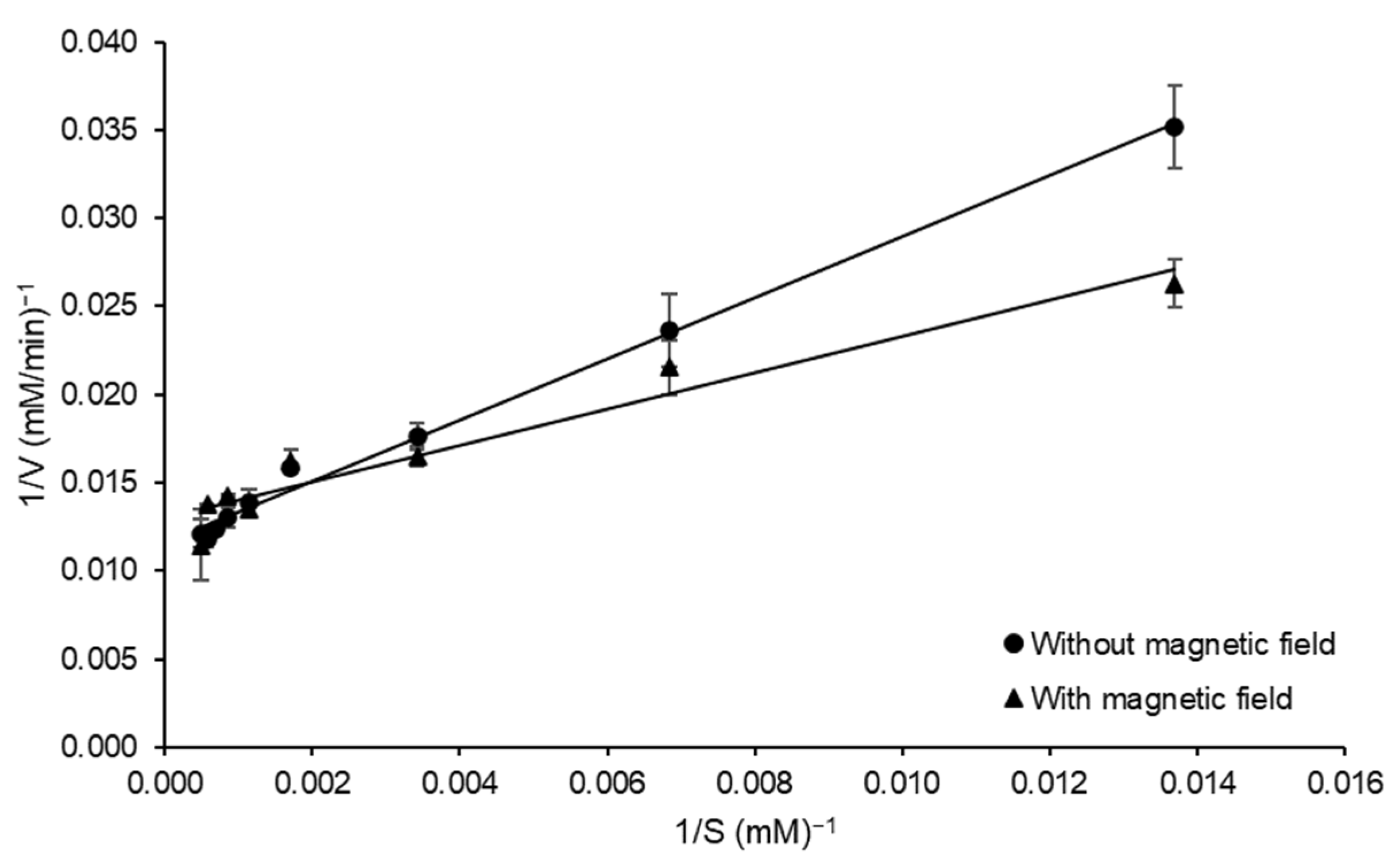
2.6. Effect of Static Magnetic Field of Kinetic Parameters of FFase Catalyzed Reaction
3. Results and Discussion
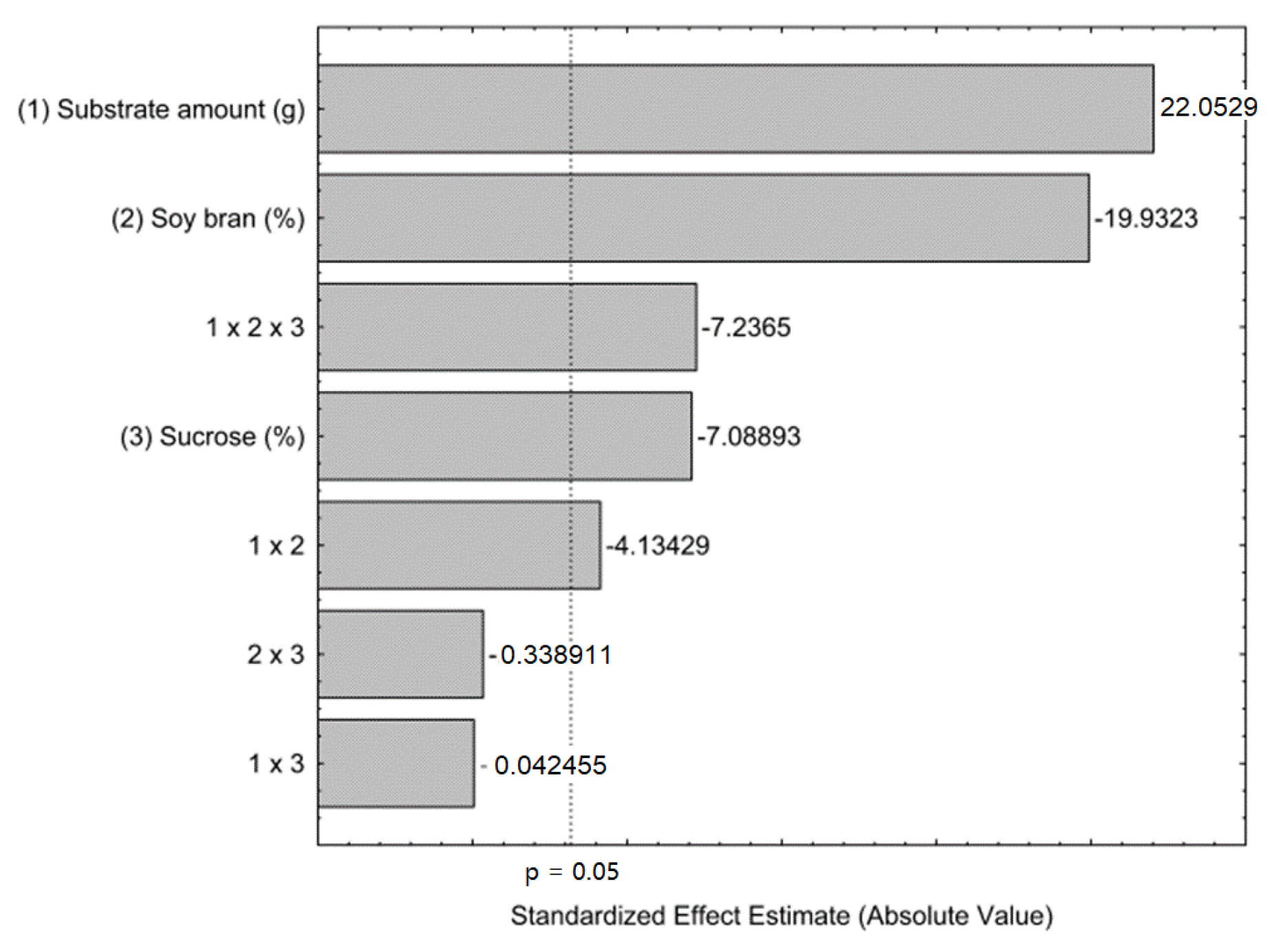
3.1. Production of FFase by Solid-State Fermentation
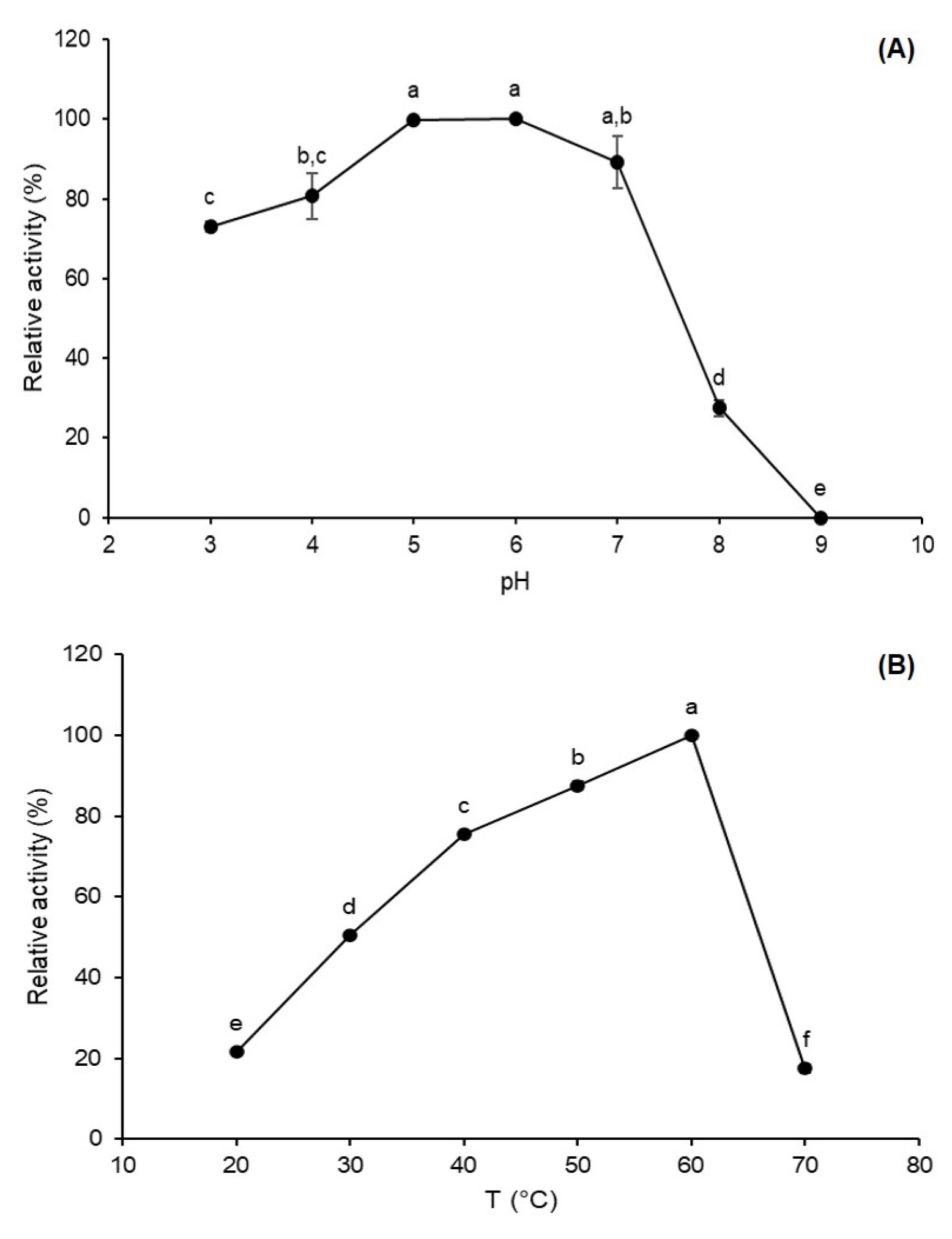
3.2. Effect of PH and Temperature on Sucrose Hydrolysis Catalyzed by FFase
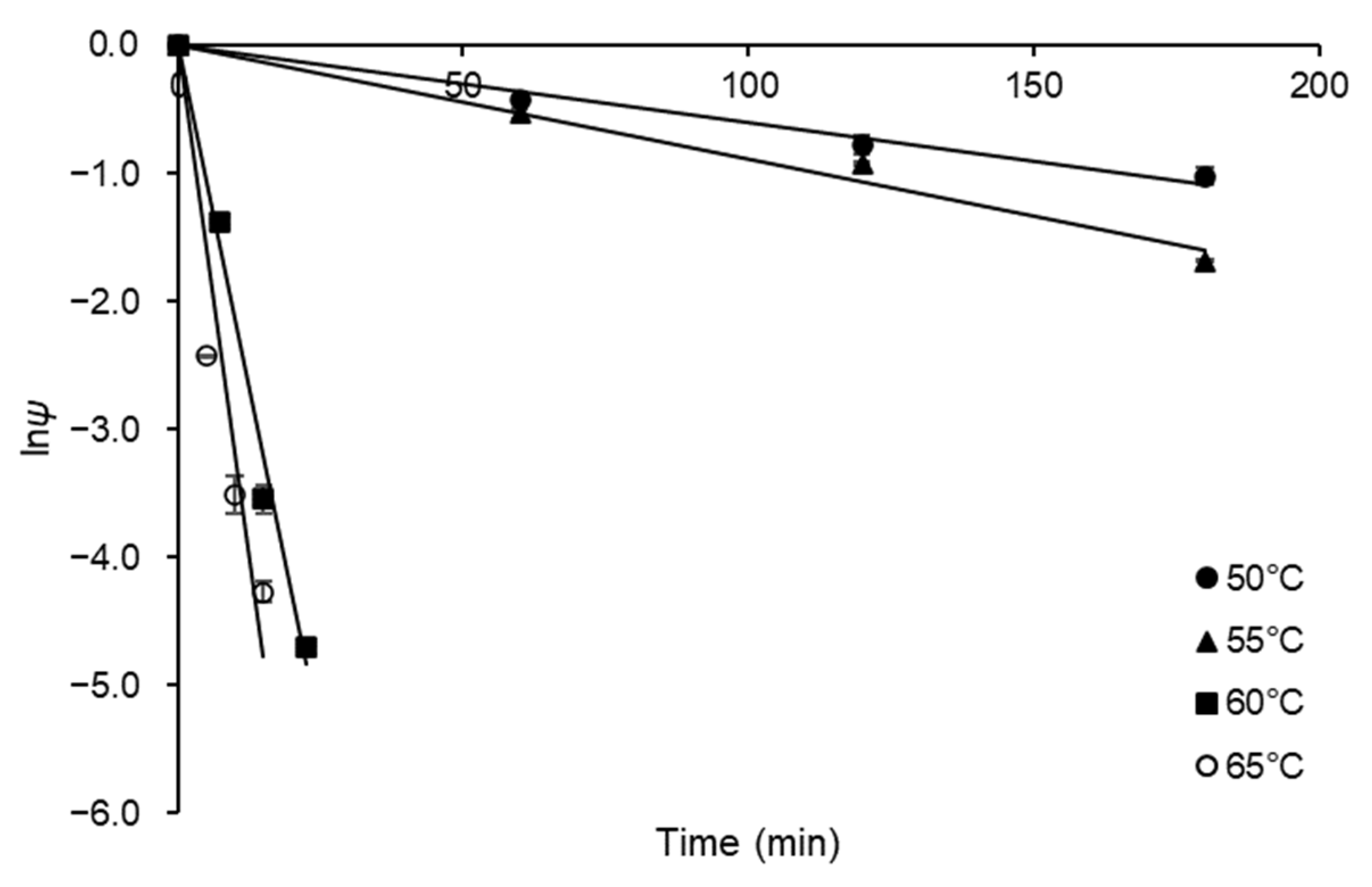
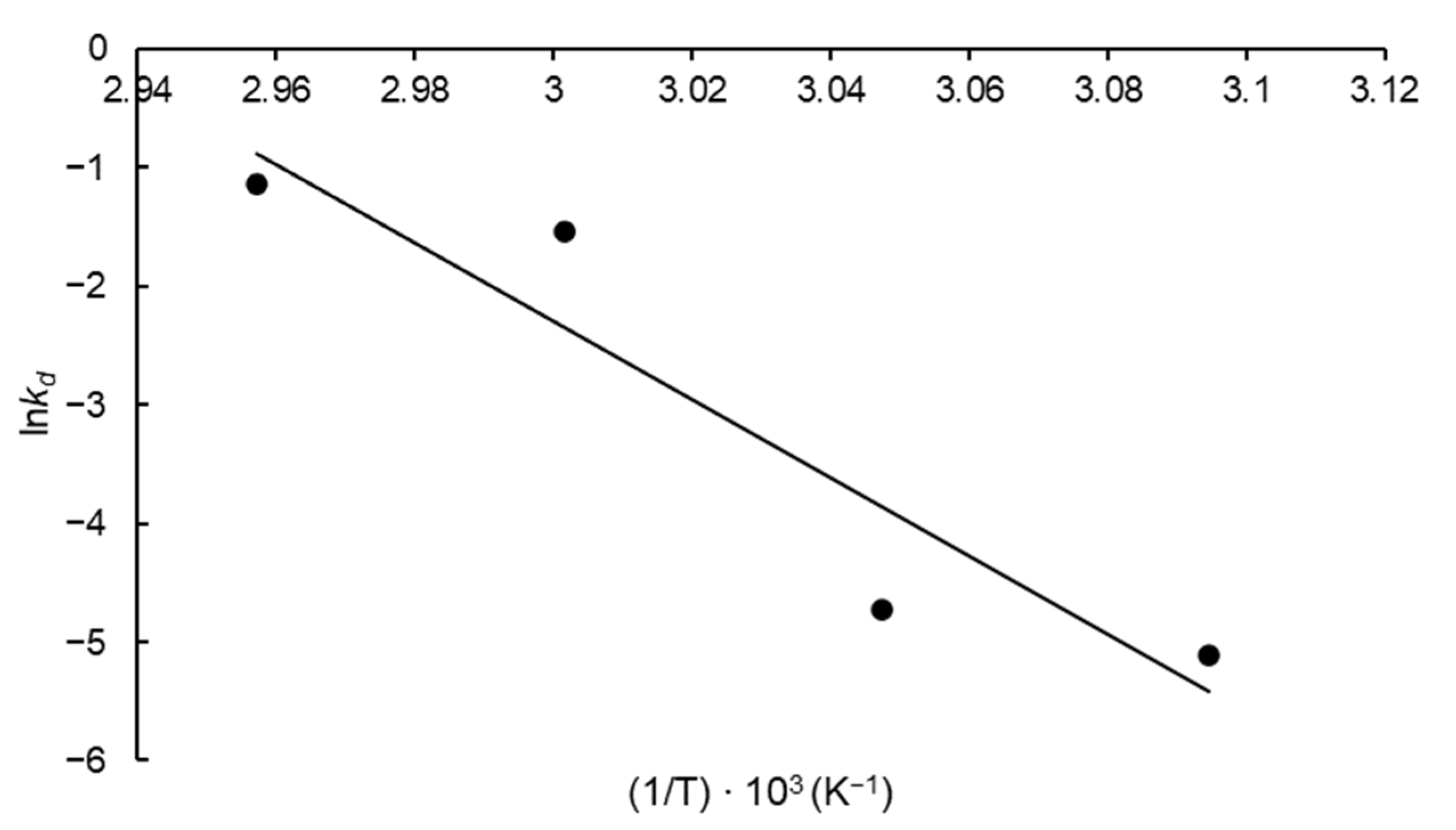
3.3. Kinetic and Thermodynamics of FFase Thermal Inactivation
3.4. Influence of Static Magnetic Field on Kinetic Constants of FFase for Sucrose Hydrolysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira, R.L.; da Silva, M.F.; da Silva, S.P.; de Araújo, A.C.V.; Cavalcanti, J.V.F.L.; Converti, A.; Porto, T.S. Fructo-oligosaccharides production by an Aspergillus aculeatus commercial enzyme preparation with fructosyltransferase activity covalently immobilized on Fe3O4–chitosan-magnetic nanoparticles. Int. J. Biol. Macromol. 2020, 150, 922–929. [Google Scholar] [CrossRef]
- Martins, G.N.; Ureta, M.M.; Tymczyszyn, E.E.; Castilho, P.C.; Gomez-Zavaglia, A. Technological Aspects of the Production of Fructo and Galacto-Oligosaccharides. Enzymatic Synthesis and Hydrolysis. Front. Nutr. 2019, 6, 54. [Google Scholar] [CrossRef]
- Antošová, M.; Polakovič, M. Fructosyltransferases: The Enzymes Catalyzing Production of Fructooligosaccharides. Chem. Pap. 2001, 55, 350–358. [Google Scholar]
- Kumar, C.G.; Sripada, S.; Poornachandra, Y. Status and Future Prospects of Fructooligosaccharides as Nutraceuticals. In Role of Materials Science in Food Bioengineering; Grumezescu, A., Holban, A.-M., Eds.; Elsevier: London, UK, 2018; pp. 451–503. ISBN 9780128114483. [Google Scholar]
- Picazo, B.; Flores-Gallegos, A.C.; Muñiz-Márquez, D.B.; Flores-Maltos, A.; Michel-Michel, M.R.; de la Rosa, O.; Rodríguez-Jasso, R.M.; Rodríguez-Herrera, R.; Aguilar-González, C.N. Enzymes for Fructooligosaccharides Production: Achievements and Opportunities. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Elsevier: London, UK, 2019; pp. 303–320. ISBN 9780128132807. [Google Scholar]
- Choukade, R.; Kango, N. Characterization of a mycelial fructosyltransferase from Aspergillus tamarii NKRC 1229 for efficient synthesis of fructooligosaccharides. Food Chem. 2019, 286, 434–440. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.L.; da Silva, M.F.; Converti, A.; Porto, T.S. Production of β-fructofuranosidase with transfructosylating activity by Aspergillus tamarii URM4634 Solid-State Fermentation on agroindustrial by-products. Int. J. Biol. Macromol. 2020, 144, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.M.S.; Brandão-Costa, R.M.P.; Carneiro da Cunha, M.N.; Rodrigues, H.O.S.; Porto, A.L.F. Purification and biochemical characterization of an extracellular fructosyltransferase-rich extract produced by Aspergillus tamarii Kita UCP1279. Biocatal. Agric. Biotechnol. 2020, 26, 101647. [Google Scholar] [CrossRef]
- Batista, J.M.S.; Brandão-Costa, R.M.P.; Cardoso, K.B.B.; Nascimento, T.P.; Albuquerque, W.W.C.; Carneiro-da-Cunha, M.N.; Porto, C.S.; Bezerra, R.P.; Figueiredo Porto, A.L. Biotechnological purification of a β-fructofuranosidase (β-FFase) from Aspergillus tamarii kita: Aqueous two-phase system (PEG/Citrate) and biochemical characterization. Biocatal. Agric. Biotechnol. 2021, 35, 102070. [Google Scholar] [CrossRef]
- Soccol, C.R.; da Costa, E.S.; Letti, L.A.; Karp, S.G.; Woiciechowski, A.L.; de Souza Vandenberghe, L.P. Recent developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- De la Rosa, O.; Flores-Gallegos, A.C.; Muñíz-Marquez, D.; Nobre, C.; Contreras-Esquivel, J.C.; Aguilar, C.N. Fructooligosaccharides production from agro-wastes as alternative low-cost source. Trends Food Sci. Technol. 2019, 91, 139–146. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R.L.; Silva, M.F.; Converti, A.; Porto, T.S. Biochemical characterization and kinetic/thermodynamic study of Aspergillus tamarii URM4634 β-fructofuranosidase with transfructosylating activity. Biotechnol. Prog. 2019, 35, e2879. [Google Scholar] [CrossRef]
- Wasak, A.; Drozd, R.; Jankowiak, D.; Rakoczy, R. Rotating magnetic field as tool for enhancing enzymes properties—Laccase case study. Sci. Rep. 2019, 9, 3707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jia, S.; Ran, J.; Wu, S. Effects of static magnetic field on activity and stability of immobilized α-amylase in chitosan bead. Catal. Commun. 2010, 11, 364–367. [Google Scholar] [CrossRef]
- Emamdadi, N.; Gholizadeh, M.; Housaindokht, M.R. Investigation of static magnetic field effect on horseradish peroxidase enzyme activity and stability in enzymatic oxidation process. Int. J. Biol. Macromol. 2021, 170, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Fraga, F.C.; Valério, A.; de Oliveira, V.A.; Di Luccio, M.; de Oliveira, D. Effect of magnetic field on the Eversa® Transform 2.0 enzyme: Enzymatic activity and structural conformation. Int. J. Biol. Macromol. 2019, 122, 653–658. [Google Scholar] [CrossRef]
- Albuquerque, W.; Nascimento, T.; Brandão-Costa, R.; Fernandes, T.; Porto, A. Static magnetic field effects on proteases with fibrinolytic activity produced by Mucor subtilissimus. Bioelectromagnetics 2017, 38, 109–120. [Google Scholar] [CrossRef]
- Sangeetha, P.T.; Ramesh, M.N.; Prapulla, S.G. Production of fructosyl transferase by Aspergillus oryzae CFR 202 in solid-state fermentation using agricultural by-products. Appl. Microbiol. Biotechnol. 2004, 65, 530–537. [Google Scholar] [CrossRef]
- Rustiguel, C.B.; de Oliveira, A.H.C.; Terenzi, H.F.; Jorge, J.A.; Souza Guimarães, L.H. Biochemical properties of an extracellular β-D-fructofuranosidase II produced by Aspergillus phoenicis under Solid-Sate Fermentation using soy bran as substrate. Electron. J. Biotechnol. 2011, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dhake, A.B.; Patil, M.B. Effect of substrate feeding on production of fructosyltransferase by Penicillium purpurogenum. Braz. J. Microbiol. 2007, 38, 194–199. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Dapper, T.B.; Arfelli, C.V.; Henn, C.; Simões, M.R.; dos Santos, M.F.; Della Torre, C.L.; da Conceição Silva, J.L.; de Cássia Garcia Simão, R.; Kadowaki, M.K. β-Fructofuranosidase production by Aspergillus versicolor isolated from Atlantic forest and grown on apple pomace. Afr. J. Microbiol. Res. 2016, 10, 938–948. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Zheng, X.; Huang, M.; Wu, M.; Yan, Y.; Pan, J.; Yang, Q.; Duan, C.J.; Liu, J.L.; Feng, J.X. Purification and biochemical characterization of a novel β-fructofuranosidase from Penicillium oxalicum with transfructosylating activity producing neokestose. Process Biochem. 2015, 50, 1237–1246. [Google Scholar] [CrossRef]
- Smaali, I.; Jazzar, S.; Soussi, A.; Muzard, M.; Aubry, N.; Nejib Marzouki, M. Enzymatic synthesis of fructooligosaccharides from date by-products using an immobilized crude enzyme preparation of β-d-fructofuranosidase from Aspergillus awamori NBRC 4033. Biotechnol. Bioprocess Eng. 2012, 17, 385–392. [Google Scholar] [CrossRef]
- Vaňková, K.; Onderková, Z.; Antošová, M.; Polakovič, M. Design and economics of industrial production of fructooligosaccharides. Chem. Pap. 2008, 62, 375–381. [Google Scholar] [CrossRef]
- Converti, A.; Pessoa, A.; Silva, J.C.; de Oliveira, R.L.; Porto, T.S. Thermodynamics Applied to Biomolecules. In Pharmaceutical Biotechnology; Pessoa, A., Vitolo, M., Long, P.F., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 29–42. [Google Scholar]
- Tayefi-Nasrabadi, H.; Asadpour, R. Effect of Heat Treatment on Buffalo (Bubalus bubalis) Lactoperoxidase Activity in Raw Milk. J. Biol. Sci. 2008, 8, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Pace, C.N. Contribution of the hydrophobic effect to globular protein stability. J. Mol. Biol. 1992, 226, 29–35. [Google Scholar] [CrossRef]
- Melikoglu, M.; Lin, C.S.K.; Webb, C. Kinetic studies on the multi-enzyme solution produced via solid state fermentation of waste bread by Aspergillus awamori. Biochem. Eng. J. 2013, 80, 76–82. [Google Scholar] [CrossRef]
- Souza, P.M.; Aliakbarian, B.; Ximenes, E.; Filho, F.; Oliveira, P.; Pessoa, A.; Converti, A. Kinetic and thermodynamic studies of a novel acid protease from Aspergillus foetidus. Int. J. Biol. Macromol. 2015, 81, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Liu, Y.; Wang, W.; Song, G.; Ren, Y. Regulation mechanism of magnetic field on pectinase and its preliminary application in postharvest sapodilla (Manilkara zapota). Food Chem. 2023, 409, 135300. [Google Scholar] [CrossRef] [PubMed]
- Timmel, C.R.; Till, U.; Brocklehurst, B.; Mclauchlan, K.A.; Hore, P.J. Effects of weak magnetic fields on free radical recombination reactions. Mol. Phys. 1998, 95, 71–89. [Google Scholar] [CrossRef]
- Jia, J.; Ma, H.; Zhao, W.; Wang, Z.; Tian, W.; Luo, L.; He, R. The use of ultrasound for enzymatic preparation of ACE-inhibitory peptides from wheat germ protein. Food Chem. 2010, 119, 336–342. [Google Scholar] [CrossRef]
- Leong, S.Y.; Oey, I. Effect of pulsed electric field treatment on enzyme kinetics and thermostability of endogenous ascorbic acid oxidase in carrots (Daucus carota cv. Nantes). Food Chem. 2014, 146, 538–547. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Substrate Amount (g) | Soy Bran (%) | Sucrose (%) | FFase Activity (U mL−1) |
|---|---|---|---|---|
| 1 | 3 | 80 | 25 | 123.90 |
| 2 | 7 | 80 | 25 | 202.47 |
| 3 | 3 | 20 | 25 | 29.88 |
| 4 | 7 | 20 | 25 | 134.15 |
| 5 | 3 | 80 | 15 | 66.14 |
| 6 | 7 | 80 | 15 | 204.31 |
| 7 | 3 | 20 | 15 | 29.27 |
| 8 | 7 | 20 | 15 | 73.23 |
| 9 (C) | 5 | 50 | 20 | 124.27 |
| 10 (C) | 5 | 50 | 20 | 119.34 |
| 11 (C) | 5 | 50 | 20 | 119.41 |
| 12 (C) | 5 | 50 | 20 | 110.23 |
| a T (°C) | bkd (min−1) | R2 | ct1/2 (min) | dD-Value (min) | e Z (°C) | fE*d (kJ mol−1) | g ΔG*d (kJ mol−1) | h ΔH*d (kJ mol−1) | i ΔS*d (J mol−1 K−1) |
|---|---|---|---|---|---|---|---|---|---|
| 50 | 0.006 | 0.993 | 115.52 | 383.76 | 7.61 | 274.29 | 104.10 | 271.60 | 518.33 |
| 55 | 0.009 | 0.993 | 77.88 | 258.72 | 104.68 | 271.56 | 508.55 | ||
| 60 | 0.215 | 0.995 | 3.22 | 10.71 | 97.50 | 271.52 | 522.36 | ||
| 65 | 0.318 | 0.971 | 2.16 | 7.19 | 97.88 | 271.48 | 513.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, R.L.; dos Santos, A.F.A.; Cardoso, B.A.; da Silva Santos, T.S.; de Campos-Takaki, G.M.; Porto, T.S.; Porto, C.S. Production, Kinetic/Thermodynamic Study, and Evaluation of the Influence of Static Magnetic Field on Kinetic Parameters of β-Fructofuranosidase from Aspergillus tamarii Kita UCP 1279 Produced by Solid-State Fermentation. BioTech 2023, 12, 21. https://doi.org/10.3390/biotech12010021
de Oliveira RL, dos Santos AFA, Cardoso BA, da Silva Santos TS, de Campos-Takaki GM, Porto TS, Porto CS. Production, Kinetic/Thermodynamic Study, and Evaluation of the Influence of Static Magnetic Field on Kinetic Parameters of β-Fructofuranosidase from Aspergillus tamarii Kita UCP 1279 Produced by Solid-State Fermentation. BioTech. 2023; 12(1):21. https://doi.org/10.3390/biotech12010021
Chicago/Turabian Stylede Oliveira, Rodrigo Lira, Aldeci França Araújo dos Santos, Bianca Alencar Cardoso, Thayanne Samille da Silva Santos, Galba Maria de Campos-Takaki, Tatiana Souza Porto, and Camila Souza Porto. 2023. "Production, Kinetic/Thermodynamic Study, and Evaluation of the Influence of Static Magnetic Field on Kinetic Parameters of β-Fructofuranosidase from Aspergillus tamarii Kita UCP 1279 Produced by Solid-State Fermentation" BioTech 12, no. 1: 21. https://doi.org/10.3390/biotech12010021