

Plug-and-Display Photo-Switchable Systems on Plant Virus Nanoparticles
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of Proteins
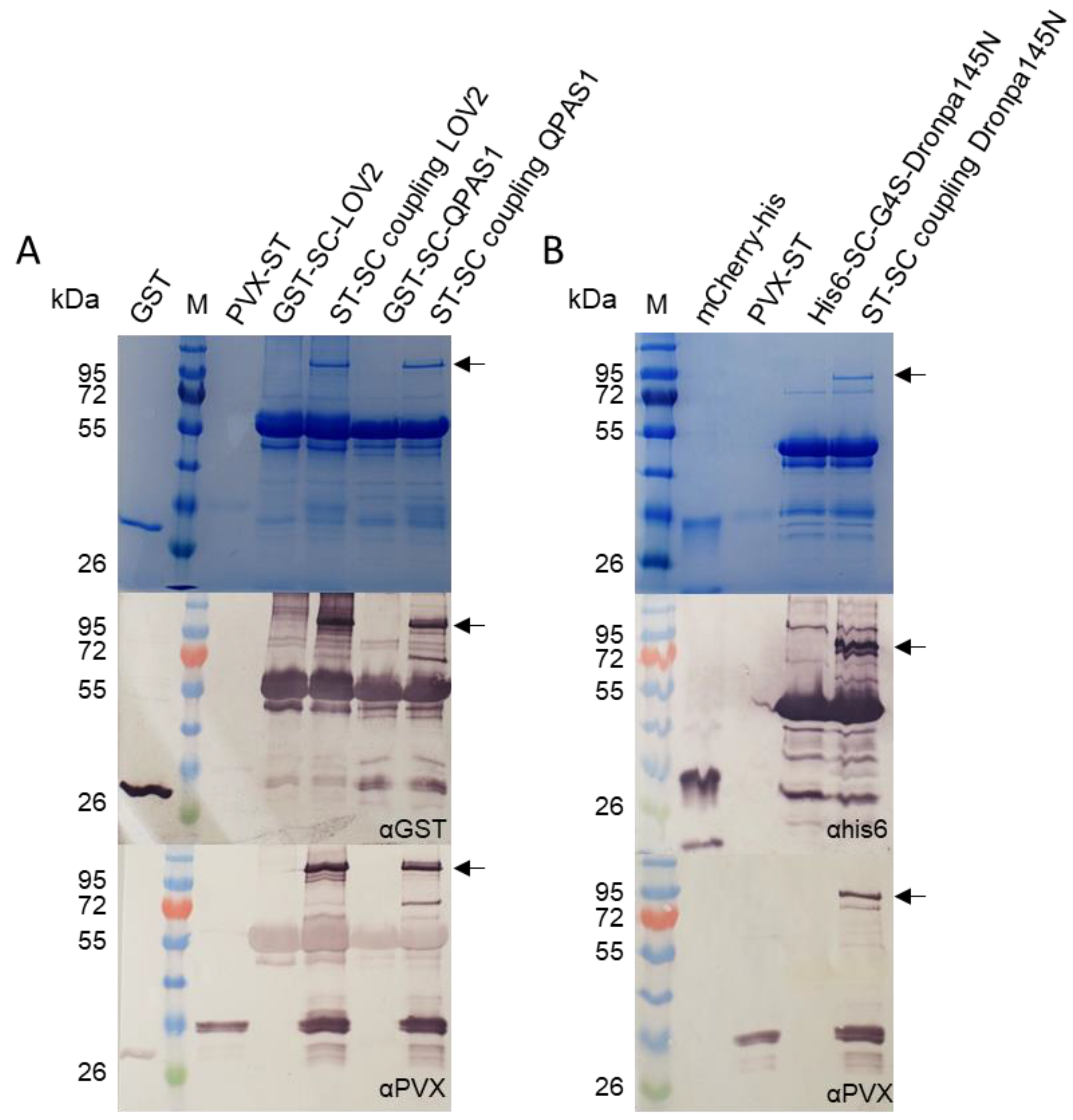
2.2. ST/SC Reaction with Purified Components
2.3. Coupling ST/SC-Tagged Components in Crude Extracts and Purification of the Complexes
2.4. In Vitro Interaction Analysis—LOVTRAP and BphP1/QPAS1 Systems
2.5. Optical Switching of Dronpa145N
2.6. Protein Interactions on the VNP Surface—LOVTRAP and BphP1/QPAS1 Systems
2.7. Protein Interactions on the VNP Surface—Dronpa145N System
3. Results
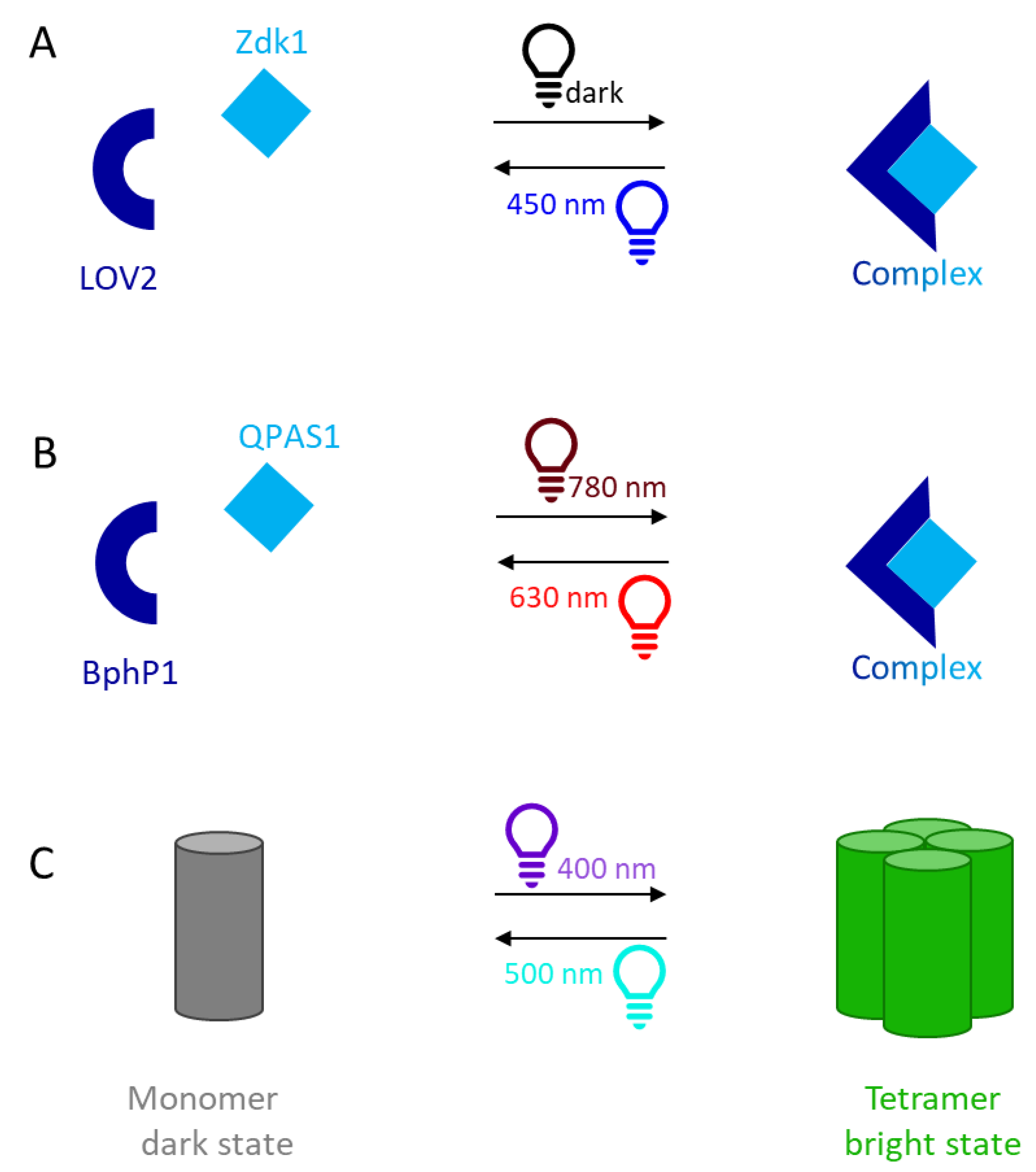
3.1. LOVTRAP
3.2. BphP1/QPAS1
3.3. Dronpa145N
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salinas, F.; Rojas, V.; Delgado, V.; Agosin, E.; Larrondo, L.F. Optogenetic switches for light-controlled gene expression in yeast. Appl. Microbiol. Biotechnol. 2017, 101, 2629–2640. [Google Scholar] [CrossRef]
- Dowbaj, A.M.; Jenkins, R.P.; Williamson, D.; Heddleston, J.M.; Ciccarelli, A.; Fallesen, T.; Hahn, K.M.; O’Dea, R.D.; King, J.R.; Montagner, M.; et al. An optogenetic method for interrogating YAP1 and TAZ nuclear–cytoplasmic shuttling. J. Cell Sci. 2021, 134, jcs253484. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Tan, P.; Zhu, L.; Huang, K.; Nguyen, N.T.; Wang, R.; Guo, L.; Li, L.; Yang, Y.; Huang, Z.; et al. Circularly permuted LOV2 as a modular photoswitch for optogenetic engineering. Nat. Chem. Biol. 2021, 17, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Yoshii, T.; Ikuta, M.; Tsukiji, S. Synthetic Protein Condensates That Inducibly Recruit and Release Protein Activity in Living Cells. J. Am. Chem. Soc. 2021, 143, 6434–6446. [Google Scholar] [CrossRef]
- Redchuk, T.A.; Karasev, M.M.; Verkhusha, P.V.; Donnelly, S.K.; Hülsemann, M.; Virtanen, J.; Moore, H.M.; Vartiainen, M.K.; Hodgson, L.; Verkhusha, V. Optogenetic regulation of endogenous proteins. Nat. Commun. 2020, 11, 605. [Google Scholar] [CrossRef] [Green Version]
- Lyu, S.; Fang, J.; Duan, T.; Fu, L.; Liu, J.; Li, H. Optically controlled reversible protein hydrogels based on photoswitchable fluorescent protein Dronpa. Chem. Commun. 2017, 53, 13375–13378. [Google Scholar] [CrossRef]
- Kolar, K.; Knobloch, C.; Stork, H.; Znidaric, M.; Weber, W. OptoBase: A Web Platform for Molecular Optogenetics. ACS Synth. Biol. 2018, 7, 1825–1828. [Google Scholar] [CrossRef]
- Wang, H.; Vilela, M.; Winkler, A.; Tarnawski, M.; Schlichting, I.; Yumerefendi, H.; Kuhlman, B.; Liu, R.; Danuser, G.; Hahn, K.M. LOVTRAP: An optogenetic system for photoinduced protein dissociation. Nat. Methods 2016, 13, 755–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Hahn, K.M. LOVTRAP: A Versatile Method to Control Protein Function with Light. Curr. Protoc. Cell Biol. 2016, 73, 21.10.1–21.10.14. [Google Scholar] [CrossRef]
- Crosson, S.; Moffat, K. Photoexcited Structure of a Plant Photoreceptor Domain Reveals a Light-Driven Molecular Switch. Plant Cell 2002, 14, 1067–1075. [Google Scholar] [CrossRef]
- Harper, S.M.; Neil, L.C.; Day, I.J.; Hore, P.J.; Gardner, K.H. Conformational Changes in a Photosensory LOV Domain Monitored by Time-Resolved NMR Spectroscopy. J. Am. Chem. Soc. 2004, 126, 3390–3391. [Google Scholar] [CrossRef] [PubMed]
- Zoltowski, B.D.; Vaccaro, B.; Crane, B.R. Mechanism-based tuning of a LOV domain photoreceptor. Nat. Chem. Biol. 2009, 5, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Ulijasz, A.T.; Vierstra, R.D. Phytochrome structure and photochemistry: Recent advances toward a complete molecular picture. Curr. Opin. Plant Biol. 2011, 14, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Redchuk, T.; Omelina, E.S.; Chernov, K.G.; Verkhusha, V.V. Near-infrared optogenetic pair for protein regulation and spectral multiplexing. Nat. Chem. Biol. 2017, 13, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Bellini, D.; Papiz, M.Z. Structure of a bacteriophytochrome and light-stimulated protomer swapping with a gene repressor. Structure 2012, 20, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Redchuk, T.A.; Karasev, M.M.; Omelina, E.S.; Verkhusha, V.V. Near-Infrared Light-Controlled Gene Expression and Protein Targeting in Neurons and Non-neuronal Cells. ChemBioChem 2018, 19, 1334–1340. [Google Scholar] [CrossRef]
- Czapar, A.E.; Zheng, Y.; Riddell, I.A.; Shukla, S.; Awuah, S.G.; Lippard, S.J.; Steinmetz, N.F. Tobacco Mosaic Virus Delivery of Phenanthriplatin for Cancer therapy. ACS Nano 2016, 10, 4119–4126. [Google Scholar] [CrossRef] [Green Version]
- Van Kan-Davelaar, H.E.J.; Van Hest, C.M.; Cornelissen, J.J.L.M.; Koay, M.S.T. Using viruses as nanomedicines. Br. J. Pharmacol. 2014, 171, 4001–4009. [Google Scholar] [CrossRef]
- Chen, M.Y.; Butler, S.S.; Chen, W.; Suh, J. Physical, chemical, and synthetic virology: Reprogramming viruses as controllable nanodevices. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1545. [Google Scholar] [CrossRef] [Green Version]
- Cormode, D.P.; Jarzyna, P.A.; Mulder, W.J.; Fayad, Z.A. Modified natural nanoparticles as contrast agents for medical imaging. Adv. Drug Deliv. Rev. 2010, 62, 329–338. [Google Scholar] [CrossRef]
- Koudelka, K.J.; Pitek, A.S.; Manchester, M.; Steinmetz, N.F. Virus-Based Nanoparticles as Versatile Nanomachines. Annu. Rev. Virol. 2015, 2, 379–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, P.; Steinmetz, N.F. Plant viral and bacteriophage delivery of nucleic acid therapeutics. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2018, 10, e1487. [Google Scholar] [CrossRef] [PubMed]
- Manchester, M.; Singh, P. Virus-based nanoparticles (VNPs): Platform technologies for diagnostic imaging. Adv. Drug Deliv. Rev. 2006, 58, 1505–1522. [Google Scholar] [CrossRef] [PubMed]
- Wen, A.M.; Steinmetz, N.F. Design of virus-based nanomaterials for medicine, biotechnology, and energy. Chem. Soc. Rev. 2016, 45, 4074–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wu, H.; Nakagawa, S.; Gao, J. Virus-derived materials: Bury the hatchet with old foes. Biomater. Sci. 2019, 8, 1058–1072. [Google Scholar] [CrossRef]
- Young, M.; Debbie, W.; Uchida, M.; Douglas, T. Plant Viruses as Biotemplates for Materials and Their Use in Nanotechnology. Annu. Rev. Phytopathol. 2008, 46, 361–384. [Google Scholar] [CrossRef]
- Shukla, S.; Ablack, A.L.; Wen, A.M.; Lee, K.L.; Lewis, J.D.; Steinmetz, N.F. Increased Tumor Homing and Tissue Penetration of the Filamentous Plant Viral Nanoparticle Potato virus X. Mol. Pharm. 2012, 10, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.L.; Hubbard, L.C.; Hern, S.; Yildiz, I.; Gratzl, M.; Steinmetz, N.F. Shape matters: The diffusion rates of TMV rods and CPMV icosahedrons in a spheroid model of extracellular matrix are distinct. Biomater. Sci. 2013, 1, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Eber, F.J.; Nagarajan, A.S.; DiFranco, N.A.; Schmidt, N.; Wen, A.M.; Eiben, S.; Twyman, R.M.; Wege, C.; Steinmetz, N.F. The Impact of Aspect Ratio on the Biodistribution and Tumor Homing of Rigid Soft-Matter Nanorods. Adv. Health Mater. 2015, 4, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Vijayan, K.; Cheng, D.; Lima, E.M.; Discher, D.E. Micelles of Different Morphologies—Advantages of Worm-like Filomicelles of PEO-PCL in Paclitaxel Delivery. Pharm. Res. 2007, 24, 2099–2109. [Google Scholar] [CrossRef]
- Nkanga, C.I.; Steinmetz, N.F. The pharmacology of plant virus nanoparticles. Virology 2021, 556, 39–61. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Antoniw, J.F.; Bar-Joseph, M.; Brunt, A.A.; Candresse, T.; Foster, G.D.; Martelli, G.P.; Milne, R.G.; Zavriev, S.K.; Fauquet, C.M. The new plant virus family Flexiviridae and assessment of molecular criteria for species demarcation. Arch. Virol. 2004, 149, 1672. [Google Scholar] [CrossRef] [Green Version]
- Verchot, J. Potato virus X: A global potato-infecting virus and type member of the Potexvirus genus. Mol. Plant Pathol. 2022, 23, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Hemenway, C. Mutations That Alter a Conserved Element Upstream of the Potato Virus X Triple Block and Coat Protein Genes Affect Subgenomic RNA Accumulation. Virology 1997, 232, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Huisman, M.J.; Linthorst, H.J.M.; Bol, J.F.; Cornelissen, B.J.C. The Complete Nucleotide Sequence of Potato Virus X and Its Homologies at the Amino Acid Level with Various Plus-stranded RNA Viruses. J. Gen. Virol. 1988, 69, 1789–1798. [Google Scholar] [CrossRef]
- Sober, J.; Jarvekulg, L.; Toots, I.; Radavsky, J.; Villems, R.; Saarma, M. Antigenic Characterization of Potato Virus X with Monoclonal Antibodies. J. Gen. Virol. 1988, 69, 1799–1807. [Google Scholar] [CrossRef]
- Kendall, A.; McDonald, M.; Bian, W.; Bowles, T.; Baumgarten, S.C.; Shi, J.; Stewart, P.L.; Bullitt, E.; Gore, D.; Irving, T.C.; et al. Structure of Flexible Filamentous Plant Viruses. J. Virol. 2008, 82, 9546–9554. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.L.; Uhde-Holzem, K.; Fischer, R.; Commandeur, U.; Steinmetz, N.F. Genetic Engineering and Chemical Conjugation of Potato Virus X. In Virus Hybrids as Nanomaterials; Humana Press: Totowa, NJ, USA, 2013; Volume 1108, pp. 3–21. [Google Scholar]
- Lico, C.; Mancini, C.; Italiani, P.; Betti, C.; Boraschi, D.; Benvenuto, E.; Baschieri, S. Plant-produced potato virus X chimeric particles displaying an influenza virus-derived peptide activate specific CD8+ T cells in mice. Vaccine 2009, 27, 5069–5076. [Google Scholar] [CrossRef]
- Nemykh, M.A.; Efimov, A.V.; Novikov, V.K.; Orlov, V.N.; Arutyunyan, A.M.; Drachev, V.A.; Lukashina, E.V.; Baratova, L.A.; Dobrov, E.N. One more probable structural transition in potato virus X virions and a revised model of the virus coat protein structure. Virology 2008, 373, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.L.; Murray, A.A.; Le, D.H.T.; Sheen, M.R.; Shukla, S.; Commandeur, U.; Fiering, S.; Steinmetz, N.F. Combination of Plant Virus Nanoparticle-Based in Situ Vaccination with Chemotherapy Potentiates Antitumor Response. Nano Lett. 2017, 17, 4019–4028. [Google Scholar] [CrossRef]
- Le, D.H.T.; Lee, K.L.; Shukla, S.; Commandeur, U.; Steinmetz, N.F. Potato virus X, a filamentous plant viral nanoparticle for doxorubicin delivery in cancer therapy. Nanoscale 2017, 9, 2348–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovlid, M.L.; Lau, J.L.; Breitenkamp, K.; Higginson, C.J.; Laufer, B.; Manchester, M.; Finn, M.G. Encapsidated Atom-Transfer Radical Polymerization in Qβ Virus-like Nanoparticles. ACS Nano 2014, 8, 8003–8014. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Dickmeis, C.; Nagarajan, A.S.; Fischer, R.; Commandeur, U.; Steinmetz, N.F. Molecular farming of fluorescent virus-based nanoparticles for optical imaging in plants, human cells and mouse models. Biomater. Sci. 2014, 2, 784–797. [Google Scholar] [CrossRef] [PubMed]
- Röder, J.; Dickmeis, C.; Fischer, R.; Commandeur, U. Systemic Infection of Nicotiana benthamiana with Potato virus X Nanoparticles Presenting a Fluorescent iLOV Polypeptide Fused Directly to the Coat Protein. BioMed Res. Int. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howarth, M.; Brune, K.D.; Ishizuka, A.S.; Biswas, S.; Leneghan, D.B.; Draper, S.J.; Bachmann, M.; Brian, I.J. Plug-and-Display: Decoration of Virus-Like Particles via isopeptide bonds for modular immunization. Sci. Rep. 2016, 6, 19234. [Google Scholar]
- Van Haren, J.; Charafeddine, R.A.; Ettinger, A.; Wang, H.; Hahn, K.M.; Wittmann, T. Local control of intracellular microtubule dynamics by EB1 photodissociation. Nat. Cell Biol. 2018, 20, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Röder, J.; Fischer, R.; Commandeur, U. Engineering Potato Virus X Particles for a Covalent Protein Based Attachment of Enzymes. Small 2017, 13, 1702151. [Google Scholar] [CrossRef]
- Peyret, H. A protocol for the gentle purification of virus-like particles produced in plants. J. Virol. Methods 2015, 225, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Passmore, L.; Russo, C. Specimen Preparation for High-Resolution Cryo-EM. In Methods in Enzymology; Europe PMC: Biggleswade, UK, 2016; Volume 579, pp. 51–86. [Google Scholar]
- Siow, K.; Britcher, L.; Kumar, S.; Griesser, H. Plasma Methods for the Generation of Chemically Reactive Surfaces for Biomolecule Immobilization and Cell Colonization—A Review. Plasma Process. Polym. 2006, 3, 392–418. [Google Scholar] [CrossRef]
- Zhou, X.X.; Chung, H.K.; Lam, A.J.; Lin, M.Z. Optical Control of Protein Activity by Fluorescent Protein Domains. Science 2012, 338, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Beauté, L.; McClenaghan, N.; Lecommandoux, S. Photo-triggered polymer nanomedicines: From molecular mechanisms to therapeutic applications. Adv. Drug Deliv. Rev. 2018, 138, 148–166. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Harris, C.C. Inflammation and cancer: An ancient link with novel potentials. Int. J. Cancer 2007, 121, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, M.; Krebs, R.; Anders, A.; Heinrich, U.; Tronnier, H. Wavelength-dependent penetration depths of ultraviolet radiation in human skin. J. Biomed. Opt. 2008, 13, 044030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baratova, L.; Grebenshchikov, N.; Dobrov, E.; Gedrovich, A.; Kashirin, I.; Shishkov, A.; Efimov, A.; Järvekülg, L.; Radavsky, Y.; Saarma, M. The organization of potato virus X coat proteins in virus particles studied by tritium planigraphy and model building. Virology 1992, 188, 175–180. [Google Scholar] [CrossRef]
- Parker, A.L.; Nicklin, S.A.; Baker, A.H. Interactions of adenovirus vectors with blood: Implications for intravascular gene therapy applications. Curr. Opin. Mol. Ther. 2008, 10, 439–448. [Google Scholar] [PubMed]
- O’Brien, G.J.; Bryant, C.J.; Voogd, C.; Greenberg, H.; Gardner, R.C.; Bellamy, A. Rotavirus VP6 Expressed by PVX Vectors in Nicotiana benthamiana Coats PVX Rods and also Assembles into Viruslike Particles. Virology 2000, 270, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Cruz, S.S.; Chapman, S.; Roberts, A.G.; Roberts, I.M.; Prior, D.A.; Oparka, K.J. Assembly and movement of a plant virus carrying a green fluorescent protein overcoat. Proc. Natl. Acad. Sci. USA 1996, 93, 6286–6290. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, M.L.; Luke, G.; Mehrotra, A.; Li, X.; Hughes, L.E.; Gani, D.; Ryan, M.D. Analysis of the aphthovirus 2A/2B polyprotein ‘cleavage’ mechanism indicates not a proteolytic reaction, but a novel translational effect: A putative ribosomal ‘skip’. J. Gen. Virol. 2001, 82, 1013–1025. [Google Scholar] [CrossRef]
- Bruckman, M.A.; Steinmetz, N.F. Chemical modification of the inner and outer surfaces of Tobacco Mosaic Virus (TMV). Methods Mol. Biol. 2014, 1108, 173–185. [Google Scholar]
- Pokorski, J.; Steinmetz, N.F. The Art of Engineering Viral Nanoparticles. Mol. Pharm. 2010, 8, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Koenig, R.; Torrance, L. Antigenic Analysis of Potato Virus X by Means of Monoclonal Antibodies. J. Gen. Virol. 1986, 67, 2145–2151. [Google Scholar] [CrossRef]
- Tozzini, A.C.; Ek, B.; Palva, E.; Hopp, H.E. Potato Virus X Coat Protein: A Glycoprotein. Virology 1994, 202, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Thrane, S.; Janitzek, C.M.; Matondo, S.; Resende, M.; Gustavsson, T.; De Jongh, W.A.; Clemmensen, S.; Roeffen, W.; Van De Vegte-Bolmer, M.; Van Gemert, G.J.; et al. Bacterial superglue enables easy development of efficient virus-like particle based vaccines. J. Nanobiotechnol. 2016, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, N.F.; Mertens, M.E.; Taurog, R.E.; Johnson, J.E.; Commandeur, U.; Fischer, R.; Manchester, M. Potato Virus X as a Novel Platform for Potential Biomedical Applications. Nano Lett. 2009, 10, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Le, D.H.; Hu, H.; Commandeur, U.; Steinmetz, N.F. Chemical addressability of potato virus X for its applications in bio/nanotechnology. J. Struct. Biol. 2017, 200, 360–368. [Google Scholar] [CrossRef]
- Halavaty, A.S.; Moffat, K. Coiled-coil dimerization of the LOV2 domain of the blue-light photoreceptor phototropin 1 from Arabidopsis thaliana. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 1316–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarabak, J. Human placental 15-hydroxyprostaglandin dehydrogenase. Proc. Natl. Acad. Sci. USA 1972, 69, 533–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongdusit, A.; Liechty, E.T.; Fox, J.M. Analysis of Three Architectures for Controlling PTP1B with Light. ACS Synth. Biol. 2021, 11, 61–68. [Google Scholar] [CrossRef]
- Kałafut, J.; Czapiński, J.; Przybyszewska-Podstawka, A.; Czerwonka, A.; Odrzywolski, A.; Sahlgren, C.; Rivero-Müller, A. Optogenetic control of NOTCH1 signalling. Cell Commun. Signal. 2022, 20, 67. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Q.; Gu, Y.; Wu, Y.; Wang, Y.; Wang, L.; Feng, S.; Hu, Y.; Zheng, Y.; Li, Y.; et al. Efficient photoactivatable Dre recombinase for cell type-specific spatiotemporal control of genome engineering in the mouse. Proc. Natl. Acad. Sci. USA 2020, 117, 33426–33435. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Huang, W.; Wu, W.-H.; Xue, B.; Xiang, D.; Li, Y.; Qin, M.; Sun, F.; Wang, W.; Zhang, W.-B.; et al. Reversible hydrogels with tunable mechanical properties for optically controlling cell migration. Nano Res. 2017, 11, 5556–5565. [Google Scholar] [CrossRef]
- Dickmeis, C.; Kauth, L.; Commandeur, U. From infection to healing: The use of plant viruses in bioactive hydrogels. WIREs Nanomed. Nanobiotechnol. 2020, 13, e1662. [Google Scholar] [CrossRef]
- Lauria, I.; Dickmeis, C.; Röder, J.; Beckers, M.; Rütten, S.; Lin, L.L.; Commandeur, U.; Fischer, H. Engineered Potato virus X nanoparticles support hydroxyapatite nucleation for improved bone tissue replacement. Acta Biomater. 2017, 62, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Schuphan, J.; Dickmeis, C.; Buhl, E.M.; Commandeur, U.; Fischer, H. Attachment of Ultralow Amount of Engineered Plant Viral Nanoparticles to Mesenchymal Stem Cells Enhances Osteogenesis and Mineralization. Adv. Healthc. Mater. 2020, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Zhang, J.; Zhang, C.; Men, Y.; Sun, H.; Li, L.-Y.; Yi, L.; Xi, Z. A highly efficient dual-diazonium reagent for protein crosslinking and construction of a virus-based gel. Org. Biomol. Chem. 2018, 16, 3353–3357. [Google Scholar] [CrossRef]
- Alexis, F.; Pridgen, E.; Molnar, L.K.; Farokhzad, O.C. Factors Affecting the Clearance and Biodistribution of Polymeric Nanoparticles. Mol. Pharm. 2008, 5, 505–515. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Time [h] | Lighting Conditions | Temperature [°C] |
|---|---|---|---|
| His6-Zdk1-mCherry | 16–18 | Daylight | 26 |
| GST-SC-LOV2 | 16–18 | Dark | 26 |
| GST-SC-QPAS1 | 16–18 | Daylight | 16 |
| BphP1-mCherry-His6 § | 16–18 | Dark | 26 |
| His6-SC-G4S-Dronpa145N | 20–22 | Dark | 20 |
| His6-mCherry-G4S-Dronpa145N | 20–22 | Dark | 20 |
| Lysis Buffer | Wash Buffer | Elution Buffer | |
|---|---|---|---|
| GST-SC-LOV2 and His6-Zdk1-mCherry | |||
| 50 mM Tris-HCl pH 8.5 150 mM NaCl 0.5% (v/v) Triton X-100 1 mM PMSF § | Low salt: 50 mM Tris-HCl pH 8.5 150 mM NaCl 0.1% (v/v) Triton X-100 5 mM 2-mercaptoethanol § | High salt: 50 mM Tris-HCl pH 8.5 650 mM NaCl 0.1% (v/v) Triton X-100 5 mM 2-mercaptoethanol § | IMAC: 50 mM Tris-HCl pH 8.5 150 mM NaCl 150 mM imidazole Glutathione Sepharose: 25 mM reduced glutathione in low-salt wash buffer |
| BphP1-mCherry-His6 | |||
| 50 mM Tris-HCl pH 8.5 300 mM NaCl 10% (v/v) glycerol | 50 mM Tris-HCl pH 8.5 300 mM NaCl 10 mM imidazole 10% (v/v) glycerol | 50 mM Tris-HCl pH 8.5 300 mM NaCl 300 mM imidazole 10% (v/v) glycerol | |
| GST-SC-QPAS1 | |||
| PBS | PBS | 50 mM Tris-HCl pH 8.5 25 mM reduced glutathione | |
| His6-SC-G4S-Dronpa145N and His6-mCherry-G4S-Dronp145N | |||
| 25 mM Tris-HCl pH 7.8 300 mM NaCl 10 mM imidazole 10% (v/v) glycerol | 25 mM Tris-HCl pH 7.8 300 mM NaCl 30 mM imidazole 10% (v/v) glycerol | 25 mM Tris-HCl pH 7.8 300 mM NaCl 300 mM imidazole 10% (v/v) glycerol | |
| Optogenetic System | Direction | Incubation | Native PAGE |
|---|---|---|---|
| LOVTRAP | Binding | Dark | Dark |
| Separation | Dark | 455 nm (blue) | |
| BphP1/QPAS1 | Binding | 780 nm (NIR) | 780 nm (NIR) |
| Separation | 630 nm (red)/dark | 630 nm (red)/dark |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauth, L.; Buhl, E.M.; Luka, J.; Richter, K.; Commandeur, U.; Dickmeis, C. Plug-and-Display Photo-Switchable Systems on Plant Virus Nanoparticles. BioTech 2022, 11, 49. https://doi.org/10.3390/biotech11040049
Kauth L, Buhl EM, Luka J, Richter K, Commandeur U, Dickmeis C. Plug-and-Display Photo-Switchable Systems on Plant Virus Nanoparticles. BioTech. 2022; 11(4):49. https://doi.org/10.3390/biotech11040049
Chicago/Turabian StyleKauth, Louisa, Eva Miriam Buhl, Julian Luka, Karolin Richter, Ulrich Commandeur, and Christina Dickmeis. 2022. "Plug-and-Display Photo-Switchable Systems on Plant Virus Nanoparticles" BioTech 11, no. 4: 49. https://doi.org/10.3390/biotech11040049