

Predator-Proofing Avian Nestboxes: A Review of Interventions, Opportunities, and Challenges

Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
3. Predation at Nestboxes
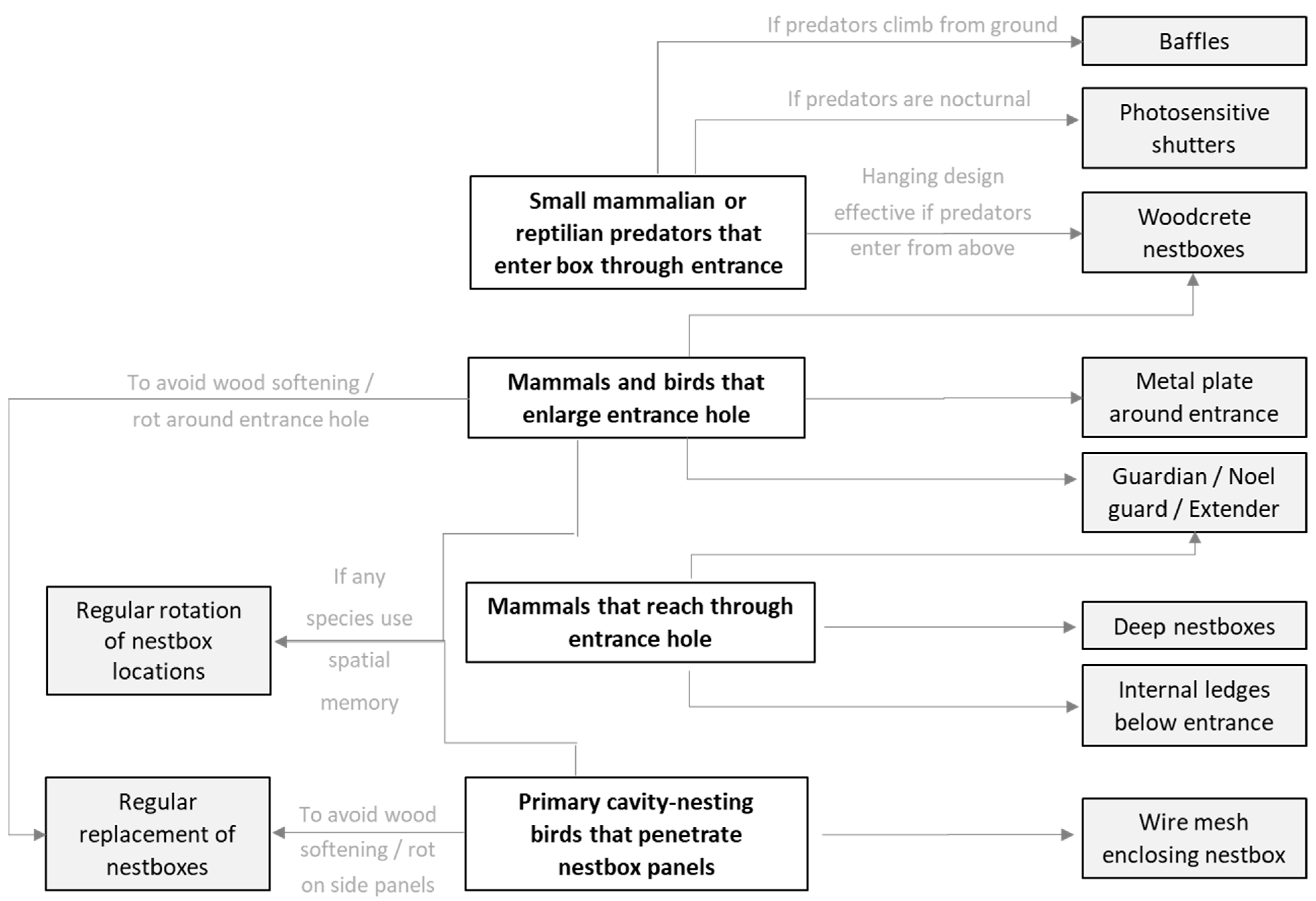
3.1. Possible Predators and Access Routes
3.2. Predation Risk
3.3. Effects of Predation
4. Nestbox Predation Prevention Strategies
4.1. Advantages and Disadvantages of Metal Plates around Entrance or Wire Mesh Panels
4.2. Advantages and Disadvantages of Deep Nestboxes
4.3. Advantages and Disadvantages of Guardian Tubes, Noel Guards, Hole Extenders, and Internal Ledges
4.4. Advantages and Disadvantages of Woodcrete Nestboxes
4.5. Advantages and Disadvantages of Photosensitive Shutters
4.6. Advantages and Disadvantages of Baffles
4.7. Advantages and Disadvantages of Regular Replacement and/or Relocation of Nestboxes
4.8. Summary of the Advantages and Disadvantages of Nestbox Predation Prevention Strategies: Weighing up the Evidence
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gill, F.B. Ornithology, 3rd ed.; W. H. Freeman and Company: New York, NY, USA, 2007. [Google Scholar]
- Dial, K.P. Evolution of avian locomotion: Correlates of flight style, locomotor modules, nesting biology, body size, development, and the origin of flapping flight. Auk 2003, 120, 941–952. [Google Scholar] [CrossRef]
- van der Hoek, Y.; Gaona, G.V.; Martin, K. The diversity, distribution and conservation status of the tree-cavity-nesting birds of the world. Divers. Distrib. 2017, 23, 1120–1131. [Google Scholar] [CrossRef]
- Martin, K.; Eadie, J.M. Nest webs: A community-wide approach to the management and conservation of cavity-nesting forest birds. For. Ecol. Manag. 1999, 115, 243–257. [Google Scholar] [CrossRef]
- Martin, K.; Aitken, K.E.H.; Wiebe, K.L. Nest-sites and nest webs for cavity-nesting communities in interior British Columbia: Nest characteristics and niche partitioning. Condor 2004, 106, 5–19. [Google Scholar] [CrossRef]
- Gibbons, P.; Lindenmayer, D. Tree Hollows and Wildlife Conservation in Australia; CSIRO Publishing: Collingwood, Australia, 2002. [Google Scholar]
- Bednarz, J.C.; Ripper, D.; Radley, P.M. Emerging concepts and research directions in the study of cavity-nesting birds: Keystone ecological processes. Condor 2004, 106, 1–4. [Google Scholar] [CrossRef]
- Newton, I. The role of nest sites in limiting the numbers of hole-nesting birds—A review. Biol. Conserv. 1994, 70, 265–276. [Google Scholar] [CrossRef]
- Newton, I. Population Limitation in Birds; Academic Press: London, UK, 1998. [Google Scholar]
- Aitken, K.E.; Martin, K. Resource selection plasticity and community responses to experimental reduction of a critical resource. Ecology 2008, 89, 971–980. [Google Scholar] [CrossRef]
- Cockle, K.L.; Martin, K.; Drever, M.C. Supply of tree-holes limits nest density of cavity-nesting birds in primary and logged subtropical Atlantic forest. Biol. Conserv. 2010, 143, 2851–2857. [Google Scholar] [CrossRef]
- Dhondt, A.A. Interspecific Competition in Birds; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Ruggera, R.A.; Schaaf, A.A.; Vivanco, C.G.; Politi, N.; Rivera, L.O. Exploring nest webs in more detail to improve forest management. For. Ecol. Manag. 2016, 372, 93–100. [Google Scholar] [CrossRef]
- Referowska-Chodak, E. Pressures and threats to nature related to human activities in european urban and suburban forests. Forests 2019, 10, 765. [Google Scholar] [CrossRef]
- Mänd, R.; Tilgar, V.; Lohmus, A.; Leivits, A. Providing nestboxes for hole-nesting birds—Does habitat matter? Biodivers. Conserv. 2005, 14, 1823–1840. [Google Scholar] [CrossRef]
- Deng, W.H.; Gao, W. Edge effects on nesting success of cavity-nesting birds in fragmented forests. Biol. Conserv. 2005, 126, 363–370. [Google Scholar] [CrossRef]
- Gregory, R.D.; Vorisek, P.; Van Strien, A.; Meyling, A.W.G.; Jiguet, F.; Fornasari, L.; Reif, J.; Chylarecki, P.; Burfield, I.J. Population trends of widespread woodland birds in Europe. Ibis 2007, 149, 78–97. [Google Scholar] [CrossRef]
- Hewson, C.M.; Amar, A.; Lindsell, J.A.; Thewlis, R.M.; Butler, S.; Smith, K.; Fuller, R.J. Recent changes in bird populations in British broadleaved woodland. Ibis 2007, 149, 14–28. [Google Scholar] [CrossRef]
- Department for Environment Food and Rural Affairs (DEFRA). Wild Bird Populations in the UK, 1970 to 2019—Updated for Wintering Waterbirds. Available online: https://www.gov.uk/government/statistics/wild-bird-populations-in-the-uk (accessed on 22 February 2023).
- Goodenough, A.E.; Elliot, S.L.; Hart, A.G. The challenges of conservation for declining migrants: Are reserve-based initiatives during the breeding season appropriate for the Pied Flycatcher Ficedula hypoleuca? Ibis 2009, 151, 429–439. [Google Scholar] [CrossRef]
- Dulisz, B.; Stawicka, A.M.; Knozowski, P.; Diserens, T.A.; Nowakowski, J.J. Effectiveness of using nest boxes as a form of bird protection after building modernization. Biodivers. Conserv. 2022, 31, 277–294. [Google Scholar] [CrossRef]
- Morris, F.O. History of British Birds; Groombridge and Sons: London, UK, 1857. [Google Scholar]
- Soper, T. The Bird Table Book, 6th ed.; David and Charles: Newton Abbot, UK, 1992. [Google Scholar]
- du Feu, C. The BTO Nestbox Guide; British Trust for Ornithology: Thetford, UK, 2003. [Google Scholar]
- Hickin, N. Bird Nest-Boxing; Stanley Paul: London, UK, 1971. [Google Scholar]
- Sanz, J.J. Latitudinal variation in female local return rate in the philopatric pied flycatcher (Ficedula hypoleuca). Auk 2001, 188, 539–543. [Google Scholar] [CrossRef]
- Merilä, J.; Wiggins, D.A. Interspecific competition for nest holes causes adult mortality in the collared flycatcher. Condor 1995, 97, 445–450. [Google Scholar]
- Twedt, D.J.; Henne-Kerr, J.L. Artificial cavities enhance breeding bird densities in managed cottonwood forests. Wildl. Soc. Bull. 2001, 29, 680–687. [Google Scholar]
- Mänd, R.; Leivits, A.; Leivits, M.; Rodenhouse, N.L. Provision of nestboxes raises the breeding density of Great Tits Parus major equally in coniferous and deciduous woodland. Ibis 2009, 151, 487–492. [Google Scholar] [CrossRef]
- Bock, C.E.; Fleck, D.C. Avian response to nest box addition in two forests in the Colorado front range. J. Field Ornithol. 1995, 66, 352–362. [Google Scholar]
- Lõhmus, A.; Remm, J. Nest quality limits the number of hole-nesting passerines in their natural cavity-rich habitat. Acta. Oecol. 2004, 27, 125–128. [Google Scholar] [CrossRef]
- Robles, H.; Ciudad, C.; Matthysen, E. Responses to experimental reduction and increase of cavities by a secondary cavity-nesting bird community in cavity-rich Pyrenean oak forests. For. Ecol. Manag. 2012, 277, 46–53. [Google Scholar] [CrossRef]
- Purcell, K.L.; Verner, J.; Oring, L.W. A comparison of the breeding ecology of birds nesting in boxes and tree cavities. Auk 1997, 114, 646–656. [Google Scholar] [CrossRef]
- Minot, E.O.; Perrins, C.M. Interspecific interference competition—Nest sites for blue and great tits. J. Anim. Ecol. 1986, 55, 331–350. [Google Scholar] [CrossRef]
- Nilsson, S.G. Clutch size and breeding success of the pied flycatcher Ficedula hypoleuca in natural tree-holes. Ibis 1984, 126, 407–410. [Google Scholar] [CrossRef]
- Robertson, R.J.; Rendell, W.B. A comparison of the breeding ecology of a secondary cavity nesting bird, the tree swallow (Tachycineta bicolor), in nest boxes and natural cavities. Can. J. Zool. 1990, 68, 1046–1052. [Google Scholar] [CrossRef]
- Norris, A.R.; Aitken, K.E.; Martin, K.; Pokorny, S. Nest boxes increase reproductive output for Tree Swallows in a forest grassland matrix in central British Columbia. PLoS ONE 2018, 13, e0204226. [Google Scholar] [CrossRef]
- Kuitunen, M.T.; Aleknonis, A. Nest predation and breeding success in common treecreepers nesting in boxes and natural cavities. Ornis Fenn. 1992, 69, 7–12. [Google Scholar]
- Nilsson, S.G. Evolution of hole-nesting in birds: On balancing selection pressures. Auk 1986, 103, 432–435. [Google Scholar]
- Mitrus, C. A comparison of the breeding ecology of Collared Flycatchers nesting in boxes and natural cavities. J. Field Ornithol. 2003, 74, 293–299. [Google Scholar] [CrossRef]
- Nilsson, S.G. Kullstorlek och häckingsframgång i holkar och naturliga hål [Translation: Clutch size and breeding success of birds in nest boxes and natural cavities]. Var Fagelvarld 1975, 34, 207–211. [Google Scholar]
- Brightsmith, D.J. Competition, predation and nest niche shifts among tropical cavity nesters: Ecological evidence. J. Avian Biol. 2005, 36, 74–83. [Google Scholar] [CrossRef]
- Wesołowski, T.; Tomiałojć, L. Breeding bird dynamics in a primeval temperate forest: Long-term trends in Białowieża National Park (Poland). Ecography 2005, 20, 432–453. [Google Scholar] [CrossRef]
- Crick, H.Q.P.; Baillie, S.R.; Leech, D.I. The UK Nest Record Scheme: Its value for science and conservation. Bird Study 2003, 50, 254–270. [Google Scholar] [CrossRef]
- Lambrechts, M.M.; Adriaensen, F.; Ardia, D.R.; Artemyev, A.V.; Atienzar, F.; Banbura, J.; Barba, E.; Bouvier, J.C.; Camprodon, J.; Cooper, C.B.; et al. The design of artificial nestboxes for the study of secondary hole-nesting birds: A review of methodological inconsistencies and potential biases. Acta Ornithol. 2010, 45, 1–26. [Google Scholar] [CrossRef]
- Winkler, D.W.; Allen, P.E. Effects of handicapping on female condition and reproduction in tree swallows (Tachycineta bicolor). Auk 1995, 112, 737–747. [Google Scholar]
- García-Navas, V.; Arroyo, L.; Sanz, J.J. Woodcrete nestboxes: Are they adequate for multibrooded species? Ardeola–Int. J. Ornithol. 2010, 57, 159–166. [Google Scholar]
- Fokkema, R.W.; Ubels, R.; Tinbergen, J.M. Experimentally quantifying the effect of nest-site depth on the predation risk and breeding success of Blue Tits. Auk 2018, 135, 919–932. [Google Scholar] [CrossRef]
- Stojanovic, D.; Owens, G.; Young, C.M.; Alves, F.; Heinsohn, R. Do nest boxes breed the target species or its competitors? A case study of a critically endangered bird. Restor. Ecol. 2021, 29, e13319. [Google Scholar] [CrossRef]
- Dunn, E. Predation by weasels (Mustela nivalis) on breeding tits (Parus spp.) in relation to the density of tits and rodents. J. Anim. Ecol. 1977, 46, 633–652. [Google Scholar] [CrossRef]
- McCleery, R.H.; Perrins, C.M. Evidence for the impact of predators on the population of great tits (Parus major) in Wytham wood. In Birds Population Studies: Their Relevance for Conservation and Management; Oxford University Press: Oxford, UK, 1991; pp. 129–147. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E. Nesting success of the great crested flycatcher in nestboxes and in tree cavities: Are nestboxes safer from nest predation? Wilson Bull. 2002, 114, 179–185. [Google Scholar] [CrossRef]
- Kaliński, A.; Wawrzyniak, J.; Banbura, M.; Skwarska, J.; Zielinski, P.; Gladalski, M.; Banbura, J. Does the threat of European Pine Marten (Martes martes) predation influence the height of nests built by Blue Tits (Cyanistes caeruleus) and Great Tits (Parus major)? Avian Biol. Res. 2014, 7, 83–90. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Hartley, I.R. Covering nestboxes with wire mesh reduces great spotted woodpecker Dendrocopos major predation of blue tit Cyanistes caeruleus nestlings, Lancashire, England. Conserv. Evid. 2008, 5, 45–46. [Google Scholar]
- Skwarska, J.A.; Kalinski, A.; Wawrzyniak, J.; Banbura, J. Opportunity makes a predator: Great Spotted Woodpecker predation on Tit broods depends on nestbox design. Ornis Fenn. 2009, 86, 109–112. [Google Scholar]
- Evans, M.R.; Lank, D.B.; Boyd, W.S.; Cooke, F. A comparison of the characteristics and fate of Barrow’s Goldeneye and Bufflehead nests in nestboxes and natural cavities. Condor 2002, 104, 610–619. [Google Scholar] [CrossRef]
- Sonerud, G. Nest hole shift in Tengmalm’s owl Aegolius funereus as defence against nest predation involving long-term memory in the predator. J. Anim. Ecol. 1985, 54, 179–192. [Google Scholar] [CrossRef]
- Sonerud, G.A. Reduced predation by pine martens on nests of Tengmalm’s owl in relocated boxes. Anim. Behav. 1989, 37, 332–334. [Google Scholar] [CrossRef]
- Sonerud, G.A.; Fjeld, P.E. Long-term-memory in egg predators—An experiment with a hooded crow. Ornis Scand. 1987, 18, 323–325. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Johnsson, K.; Tjernberg, M. Is avoidance by black woodpeckers of old nest holes due to predators? Anim. Behav. 1991, 41, 439–441. [Google Scholar] [CrossRef]
- Pelech, S.A.; Smith, J.N.M.; Boutin, S. A predator’s perspective of nest predation: Predation by red squirrels is learned, not incidental. Oikos 2010, 119, 841–851. [Google Scholar] [CrossRef]
- Fontaine, J.J.; Martin, T.E. Parent birds assess nest predation risk and adjust their reproductive strategies. Ecol. Lett. 2006, 9, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.Y.; Sieving, K.E.; Fletcher, R.J.; Wright, C.A. Increased perception of predation risk to adults and offspring alters avian reproductive strategy and performance. Behav. Ecol. 2014, 25, 509–519. [Google Scholar] [CrossRef]
- Lima, S.L. Predators and the breeding bird: Behavioral and reproductive flexibility under the risk of predation. Biol. Rev. 2009, 84, 485–513. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.N. Assessment of predation risk through referential communication in incubating birds. Sci. Rep. 2015, 5, 10239. [Google Scholar] [CrossRef]
- Chalfoun, A.D.; Martin, T.E. Parental investment decisions in response to ambient nest-predation risk versus actual predation on the prior nest. Condor 2010, 112, 701–710. [Google Scholar] [CrossRef]
- Amo, L.; Galvan, I.; Tomas, G.; Sanz, J.J. Predator odour recognition and avoidance in a songbird. Funct. Ecol. 2008, 22, 289–293. [Google Scholar] [CrossRef]
- Slagsvold, T. Clutch size variation of birds in relation to nest predation—On the cost of reproduction. J. Anim. Ecol. 1984, 53, 945–953. [Google Scholar] [CrossRef]
- Farnsworth, G.L.; Simons, T.R. How many baskets? Clutch sizes that maximize annual fecundity of multiple-brooded birds. Auk 2001, 118, 973–982. [Google Scholar] [CrossRef]
- McCleery, R.H.; Clobert, J.; Julliard, R.; Perrins, C.M. Nest predation and delayed cost of reproduction in the great tit. J. Anim. Ecol. 1996, 65, 96–104. [Google Scholar] [CrossRef]
- Nilsson, J.Ǻ.; Svensson, E. The cost of reproduction: A new link between current reproductive effort and future reproductive success. Proc. R. Soc. Lond. B Biol. Sci. 1996, 263, 711–714. [Google Scholar]
- Olofsson, H.; Ripa, J.; Jonzen, N. Bet-hedging as an evolutionary game: The trade-off between egg size and number. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 2963–2969. [Google Scholar] [CrossRef] [PubMed]
- Ekner, A.; Tryjanowski, P. Do small hole nesting passerines detect cues left by a predator? A test on winter roosting sites. Acta Ornithol. 2008, 43, 107–111. [Google Scholar] [CrossRef]
- Dhondt, A.A.; Blondel, J.; Perret, P. Why do Corsican Blue Tits Cyanistes caeruleus ogliastrae not use nest boxes for roosting? J. Ornithol. 2010, 151, 95–101. [Google Scholar] [CrossRef]
- Broughton, R.K. Current and future impacts of nest predation and nest-site competition by invasive eastern grey squirrels Sciurus carolinensis on European birds. Mamm. Rev. 2020, 50, 38–51. [Google Scholar] [CrossRef]
- Carter, J.H.; Walters, J.R.; Everhart, S.H.; Doerr, P.D. Restrictors for red-cockaded woodpecker cavities. Wildl. Soc. Bull. 1989, 17, 68–72. [Google Scholar]
- Blanc, L.A.; Walters, J.R. Cavity excavation and enlargement as mechanisms for indirect interactions in an avian community. Ecology 2008, 89, 506–514. [Google Scholar] [CrossRef]
- Charter, M.; Leshem, Y.; Halevi, S.; Izhaki, I. Nest box use by great tits in semi–arid rural residential gardens. Wilson J. Ornithol. 2010, 122, 604–608. [Google Scholar] [CrossRef]
- Orchan, Y.; Chiron, F.; Shwartz, A.; Kark, S. The complex interaction network among multiple invasive bird species in a cavity-nesting community. Biol. Invasions 2013, 15, 429–445. [Google Scholar] [CrossRef]
- Wesołowski, T. Anti-predator adaptations in nesting Marsh Tits Parus palustris: The role of nest-site security. Ibis 2002, 144, 593–601. [Google Scholar] [CrossRef]
- Maziarz, M.; Wesołowski, T.; Hebda, G.; Cholewa, M.; Broughton, R.K. Breeding success of the Great Tit Parus major in relation to attributes of natural nest cavities in a primeval forest. J. Ornithol. 2016, 157, 343–354. [Google Scholar] [CrossRef]
- van Balen, J. The relationship between nest-box size, occupation and breeding parameters of the Great Tit Parus major and some other hole-nesting species. Ardea 1984, 72, 163–175. [Google Scholar]
- Löhrl, H. Experiments on nest-site selection in great tit, Parus major. J. Ornithol. 1986, 127, 51–59. [Google Scholar] [CrossRef]
- Summers, R.; Taylor, W. Use by tits of nestboxes of different designs in pinewoods. Bird Study 1996, 43, 138–141. [Google Scholar] [CrossRef]
- Mazgajski, T.D. Nest site choice in relation to the presence of old nests and cavity depth in the starling Sturnus vulgaris. Ethol. Ecol. Evol. 2003, 15, 273–281. [Google Scholar] [CrossRef]
- Cockle, K.L.; Bodrati, A.A. Nesting of the planalto woodcreeper (Dendrocolaptes platyrostris). Wilson J. Ornithol. 2009, 121, 789–795. [Google Scholar] [CrossRef]
- Sergio, F.; Blas, J.; Baos, R.; Forero, M.G.; Donazar, J.A.; Hiraldo, F. Short- and long-term consequences of individual and territory quality in a long-lived bird. Oecologia 2009, 160, 507–514. [Google Scholar] [CrossRef]
- Major, R.E. Identification of nest predators by photography, dummy eggs, and adhesive tape. Auk 1991, 108, 190–196. [Google Scholar]
- Rowcliffe, J.M.; Carbone, C.; Jansen, P.A.; Kays, R.; Kranstauber, B. Quantifying the sensitivity of camera traps: An adapted distance sampling approach. Methods Ecol. Evol. 2011, 2, 464–476. [Google Scholar] [CrossRef]
- Surmacki, A.; Podkowa, P. The use of trail cameras to monitor species inhabiting artificial nestboxes. Ecol. Evol. 2022, 12, e8550. [Google Scholar] [CrossRef] [PubMed]
- Tomás, G.; Ruiz-Castellano, C.; Ruiz-Rodríguez, M.; Soler, J.J. Smaller distance between nest contents and cavity entrance increases risk of ectoparasitism in cavity-nesting birds. J. Avian Biol. 2020, 51, 1–11. [Google Scholar] [CrossRef]
- Blunsden, T.; Goodenough, A.E. Influence of nestbox design and nesting material on ectoparasite load for four woodland passerines. Bird Study 2023, 70, 25–36. [Google Scholar] [CrossRef]
- Wesołowski, T.; Czeszczewik, D.; Rowinski, P.; Walankiewicz, W. Nest soaking in natural holes—A serious cause of breeding failure? Ornis Fenn. 2002, 79, 132–138. [Google Scholar]
- Heenan, C.B. An overview of the factors influencing the morphology and thermal properties of avian nests. Avian Biol. Res. 2013, 6, 104–118. [Google Scholar] [CrossRef]
- Mazgajski, T.D.; Rykowska, Z. Dependence of nest mass on nest hole depth in the Great Tit Parus major. Acta Ornithol. 2008, 43, 49–55. [Google Scholar] [CrossRef]
- Alabrundzinska, J.; Kalinski, A.; Slomczynski, R.; Wawrzyniak, J.; Zielinski, P.; Banbura, J. Effects of nest characteristics on breeding success of Great Tits Parus major. Acta Ornithol. 2003, 38, 151–154. [Google Scholar] [CrossRef]
- Hilton, G.M.; Hansell, M.H.; Ruxton, G.D.; Reid, J.M.; Monaghan, P. Using artificial nests to test importance of nesting material and nest shelter for incubation energetics. Auk 2004, 121, 777–787. [Google Scholar] [CrossRef]
- Vergara, P.M. Effects of nestbox size on the nesting and renesting pattern of Aphrastura spinicauda and Troglodytes aedon. Ecol. Austral 2007, 17, 133–141. [Google Scholar]
- Gold, C.S.; Dahlsten, D.L. Prevalence, habitat selection, and biology of Protocalliphora (Diptera, Calliphoridae) found in nests of mountain and chestnut-backed chickadees in California. Hilgardia 1989, 57, 1–19. [Google Scholar] [CrossRef]
- Rendell, W.B.; Verbeek, N.A.M. Are avian ectoparasites more numerous in nestboxes with old nest material? Can. J. Zool. 1996, 74, 1819–1825. [Google Scholar] [CrossRef]
- Tripet, F.; Richner, H. The coevolutionary potential of a ‘generalist’ parasite, the hen flea Ceratophyllus gallinae. Parasitology 1997, 115, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Weddle, C.B. Effects of ectoparasites on nestling body mass in the house sparrow. Condor 2000, 102, 684–687. [Google Scholar] [CrossRef]
- Puchala, P. Detrimental effects of larval blow files (Protocalliphora azurea) on nestlings and breeding success of Tree Sparrows (Passer montanus). Can. J. Zool. 2004, 82, 1285–1290. [Google Scholar] [CrossRef]
- Tomás, G.; Merino, S.; Moreno, J.; Morales, J. Consequences of nest reuse for parasite burden and female health and condition in blue tits, Cyanistes caeruleus. Anim. Behav. 2007, 73, 805–814. [Google Scholar] [CrossRef]
- Richner, H.; Oppliger, A.; Christe, P. Effect of an ectoparasite on reproduction in great tits. J. Anim. Ecol. 1993, 62, 703–710. [Google Scholar] [CrossRef]
- Merino, S.; Potti, J. Mites and blowflies decrease growth and survival in nestling pied flycatchers. Oikos 1995, 73, 95–103. [Google Scholar] [CrossRef]
- Moore, J. Parasites and the Behaviour of Animals; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Bush, S.E.; Clayton, D.H. Anti-parasite behaviour of birds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170196. [Google Scholar] [CrossRef]
- Wesołowski, T. Host–parasite interactions in natural holes: Marsh Tits (Parus palustris) and blow flies (Protocalliphora falcozi). J. Zool. 2001, 255, 495–503. [Google Scholar] [CrossRef]
- Dudek, B.M.; Henderson, M.T.; Hudon, S.F.; Hayden, E.J.; Heath, J.A. Haematophagous ectoparasites lower survival of and have detrimental physiological effects on golden eagle nestlings. Conserv. Physiol. 2021, 9, coab060. [Google Scholar] [CrossRef]
- Wesołowski, T.; Maziarz, M. Dark tree cavities—A challenge for hole nesting birds? J. Avian Biol. 2012, 43, 454–460. [Google Scholar] [CrossRef]
- Podkowa, P.; Surmacki, A. The importance of illumination in nest site choice and nest characteristics of cavity nesting birds. Sci. Rep. 2017, 7, 1329. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.B.; Voss, M.A.; Ardia, D.R.; Austin, S.H.; Robinson, W.D. Light increases the rate of embryonic development: Implications for latitudinal trends in incubation period. Funct. Ecol. 2011, 25, 769–776. [Google Scholar] [CrossRef]
- Austin, S.H.; Hau, M.; Robinson, W.D. Effect of photoperiod on incubation period in a wild passerine, Sylvia atricapilla. J. Avian Biol. 2014, 45, 359–364. [Google Scholar] [CrossRef]
- Howe, S.; Kilgore, D.L. Convective and diffusive gas exchange in nest cavities of the northern flicker (Colaptes auratus). Physiol. Zool. 1987, 60, 707–712. [Google Scholar] [CrossRef]
- Bailey, R.L.; Bonter, D.N. Predator guards on nestboxes improve nesting success of birds. Wildl. Soc. Bull. 2017, 41, 434–441. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Kawano, K.M.; Yamaguchi, Y.; Saito, T. Small protection plates against marten predation on nestboxes. Appl. Entomol. Zool. 2005, 40, 575–577. [Google Scholar] [CrossRef]
- Williams, J.J. Bluebird. J. N. Am. Bluebird Soc. 2001, 23, 12. [Google Scholar]
- Reynolds, S.J.; Martin, G.R.; Cassey, P. Is sexual selection blurring the functional significance of eggshell coloration hypotheses? Anim. Behav. 2009, 78, 209–215. [Google Scholar] [CrossRef]
- Browne, S.J. Effect of nestbox construction and colour on the occupancy and breeding success of nesting tits Parus spp. Bird Study 2006, 53, 187–192. [Google Scholar] [CrossRef]
- García-Navas, V.; Arroyo, L.; Sanz, J.J.; Diaz, M. Effect of nestbox type on occupancy and breeding biology of Tree Sparrows Passer montanus in central Spain. Ibis 2008, 150, 356–364. [Google Scholar] [CrossRef]
- Bueno-Enciso, J.; Ferrer, E.S.; Barrientos, R.; Sanz, J.J. Effect of nestbox type on the breeding performance of two secondary hole-nesting passerines. J. Ornithol. 2016, 157, 759–772. [Google Scholar] [CrossRef]
- Imlay, T.L.; Nickerson, D.; Horn, A.G. Temperature and breeding success for Cliff Swallows (Petrochelidon pyrrhonota) nesting on man-made structures: Ecological traps? Can. J. Zool. 2019, 97, 429–435. [Google Scholar] [CrossRef]
- Greño, J.L.; Belda, E.J.; Barba, E. Influence of temperatures during the nestling period on post-fledging survival of great tit Parus major in a Mediterranean habitat. J. Avian Biol. 2008, 39, 41–49. [Google Scholar] [CrossRef]
- Andreasson, F.; Nord, A.; Nilsson, J.A. Experimentally increased nest temperature affects body temperature, growth and apparent survival in blue tit nestlings. J. Avian Biol. 2018, 49, e01620. [Google Scholar] [CrossRef]
- Griebel, I.A.; Dawson, R.D.; Clark, R.G. Cavity type influences abundance of nest-dwelling avian blow flies: An experiment with tree swallows. Ecol. Entomol. 2020, 45, 434–443. [Google Scholar] [CrossRef]
- Mennerat, A.; Charmantier, A.; Perret, P.; Hurtrez-Boussès, S.; Lambrechts, M.M. Parasite intensity is driven by temperature in a wild bird. Peer Community J. 2021, 1, e60. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Elliot, S.L.; Hart, A.G. Do orientation-based differences in nest box temperature cause differential ectoparasite load and explain patterns of nest-site selection and offspring condition in great tits? Int. J. Zool. 2011, 2011, 514913. [Google Scholar] [CrossRef]
- Castaño-Vázquez, F.; Schumm, Y.R.; Bentele, A.; Quillfeldt, P.; Merino, S. Experimental manipulation of cavity temperature produces differential effects on parasite abundances in blue tit nests at two different latitudes. Int. J. Parasitol. Parasites Wildl. 2021, 14, 287–297. [Google Scholar] [CrossRef]
- Albert, L.; Rumschlag, S.; Parker, A.; Vaziri, G.; Knutie, S.A. Elevated nest temperature has opposing effects on host species infested with parasitic nest flies. Oecologia 2023, 201, 877–886. [Google Scholar] [CrossRef]
- Belda, E.J.; Ferrandis, P.; Gil-Delgado, J.A. Clutch size variation and nest failure of the Serin Serinus serinus in orange groves. Ardeola 1995, 42, 1–10. [Google Scholar]
- Goodenough, A.E.; Stallwood, B. Differences in culturable microbial communities in bird nestboxes according to orientation and influences on offspring quality in great tits (Parus major). Microb. ecol. 2012, 63, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Deeming, D.C. Importance of nest type on the regulation of humidity in bird nests. Avian Biol. Res. 2011, 4, 23–31. [Google Scholar] [CrossRef]
- Deeming, D.C.; Mainwaring, M.C.; Hartley, I.R.; Reynolds, S.J. Local temperature and not latitude determines the design of Blue Tit and Great Tit nests. Avian Biol. Res. 2012, 5, 203–208. [Google Scholar] [CrossRef]
- Cunningham, S.J.; Martin, R.O.; Hojem, C.L.; Hockey, P.A.R. Temperatures in excess of critical thresholds threaten nestling growth and survival in a rapidly-warming arid savanna: A Study of Common Fiscals. PLoS ONE 2013, 8, e74613. [Google Scholar] [CrossRef] [PubMed]
- Mainwaring, M.C.; Hartley, I.R.; Bearhop, S.; Brulez, K.; du Feu, C.R.; Murphy, G.; Plummer, K.E.; Webber, S.L.; Reynolds, S.J.; Deeming, D.C. Latitudinal variation in blue tit and great tit nest characteristics indicates environmental adjustment. J. Biogeogr. 2012, 39, 1669–1677. [Google Scholar] [CrossRef]
- Kasprzyk, K.; Ruczynski, I. The structure of bat communities roosting in bird nest boxes in two pine monocultures in Poland. Folia Zool. 2001, 50, 107–116. [Google Scholar]
- Stojanovic, D.; Eyles, S.; Cook, H.; Alves, F.; Webb, M.; Heinsohn, R. Photosensitive automated doors to exclude small nocturnal predators from nestboxes. Anim. Conserv. 2019, 22, 297–301. [Google Scholar] [CrossRef]
- Cates, D.; Allen, N. Build Nestboxes for Wild Birds; Oregon State University, Extension Service: Corvallis, OR, USA, 2002. [Google Scholar]
- Bolen, E.G. Nesting boxes for black-bellied tree ducks. J. Wildl. Manage. 1967, 31, 794–797. [Google Scholar] [CrossRef]
- Lacki, M.J.; George, S.P.; Viscosi, P.J. Evaluation of site variables affecting nestbox use by wood ducks. Wildl. Soc. Bull. 1987, 15, 196–200. [Google Scholar]
- Laubergs, A.R.; Viksne, J. Elevated artificial nest sites for Mallard Anas platyrhynchos in Latvia. Acta Univ. Latv. 2004, 676, 107–118. [Google Scholar]
- Keo, O.; Collar, N.J.; Sutherland, W.J. Nest protectors provide a cost-effective means of increasing breeding success in Giant Ibis Thaumatibis gigantea. Bird Conserv. Int. 2009, 19, 77–82. [Google Scholar] [CrossRef]
- Cornell, K.L.; Kight, C.R.; Burdge, R.B.; Gunderson, A.R.; Hubbard, J.K.; Jackson, A.K.; LeClerc, J.E.; Pitts, M.L.; Swaddle, J.P.; Cristol, D.A. Reproductive success of eastern bluebirds (Siala sialis) on suburban golf courses. Auk 2011, 128, 577–586. [Google Scholar] [CrossRef]
- Briskie, J.V.; Shorey, L.; Massaro, M. Nest-boxes increase fledging success in the declining rifleman Acanthisitta chloris, New Zealand. Conserv. Evid. 2014, 11, 12–15. [Google Scholar]
- Mueller, A.J.; Twedt, D.J.; Bowers, E.K. Rapid adoption of nestboxes by Prothonotary Warblers (Protonotaria citrea) in mesic deciduous forest. Can. J. Zool. 2019, 97, 1109–1115. [Google Scholar] [CrossRef]
- Greene, T.C.; Jones, A. Observed responses of captive stoats (Mustela erminea) to nestboxes and metal collars used to protect kaka (Nestor meridionalis) nest cavities. N. Z. J. Ecol. 2003, 27, 139–145. [Google Scholar]
- Garnett, S.T.; Pedler, L.P.; Crowley, G.M. The breeding biology of the Glossy Black-Cockatoo Calyptorhynchus lathami on Kangaroo Island, South Australia. Emu 1999, 99, 262–279. [Google Scholar] [CrossRef]
- Williams, R.L.; Goodenough, A.E.; Hart, A.G.; Stafford, R. Using long-term volunteer records to examine dormouse (Muscardinus avellanarius) nestbox selection. PLoS ONE 2013, 8, e67986. [Google Scholar] [CrossRef]
- Navalpotro, H.; Mazzoni, D.; Senar, J.C. A plastic device fixed around trees can deter snakes from predating bird nestboxes. Anim. Biodivers. Conserv. 2021, 44, 103–108. [Google Scholar] [CrossRef]
- Fontaine, J.J.; Martin, T.E. Habitat selection responses of parents to offspring predation risk: An experimental test. Am. Nat. 2006, 168, 811–818. [Google Scholar] [CrossRef]
- Hoover, J.P. Decision rules for site fidelity in a migratory bird, the prothonotary warbler. Ecology 2003, 84, 416–430. [Google Scholar] [CrossRef]
- Schmidt, K.A.; Ostfeld, R.S.; Smyth, K.N. Spatial heterogeneity in predator activity, nest survivorship, and nest-site selection in two forest thrushes. Oecologia 2006, 148, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Chalfoun, A.D.; Martin, T.E. Facultative nest patch shifts in response to nest predation risk in the Brewer’s sparrow: A “win-stay, lose-switch” strategy? Oecologia 2010, 163, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Karell, P.; Ericsson, P.; Ericsson, D.; Haglund, K.; Morosinotto, C.; Nilsson, L.O. Mammalian nest predation induces small-scale nest site switching in territorial tawny owl (Strix aluco) females. Ornis Fenn. 2020, 97, 45–52. [Google Scholar] [CrossRef]
- Sonerud, G.A. Win—And stay, but not too long: Cavity selection by Boreal Owls to minimize nest predation by Pine Marten. J. Ornithol. 2021, 162, 839–855. [Google Scholar] [CrossRef]
- Wesołowski, T. Natural nest sites of Marsh Tit (Parus palustris) in a primaeval forest (Bialowieza National Park, Poland). Vogelwarte 1996, 38, 235–249. [Google Scholar]
- Langowska, A.; Ekner, A.; Skórka, P.; Tobolka, M.; Tryjanowski, P. Nest-site tenacity and dispersal patterns of Vespa crabro colonies located in bird nest-boxes. Sociobiology 2010, 56, 375. [Google Scholar]
- Weidinger, K.; Kocvara, R. Repeatability of nest predation in passerines depends on predator species and time scale. Oikos 2010, 119, 138–146. [Google Scholar] [CrossRef]
- Sonerud, G.A. Reduced predation by nestbox relocation—Differential effect on tengmalm owl nests and artificial nests. Ornis Scand. 1993, 24, 249–253. [Google Scholar] [CrossRef]
- Sorace, A.; Petrassi, F.; Consiglio, C. Long-distance relocation of nestboxes reduces nest predation by Pine Marten Martes martes. Bird Study 2004, 51, 119–124. [Google Scholar] [CrossRef]
- Enoksson, B. Local movements in the Nuthatch (Sitta europaea). Acta Regiae Soc. Sci. Litt. Gothob. Zool. 1987, 14, 36–47. [Google Scholar]
- Matthysen, E.; Schmidt, K.H. Natal dispersal in the nuthatch. Ornis Scand. 1987, 18, 313–316. [Google Scholar] [CrossRef]
- Tieleman, B.I.; van Noordwijk, H.J.; Williams, J.B. Nest site selection in a hot desert: Trade-off between microclimate and predation risk? Condor 2008, 110, 116–124. [Google Scholar] [CrossRef]
- Mayer, P.M.; Smith, L.M.; Ford, R.G.; Watterson, D.C.; McCutchen, M.D.; Ryan, M.R. Nest construction by a ground-nesting bird represents a potential trade-off between egg crypticity and thermoregulation. Oecologia 2009, 159, 893–901. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modification Category | Specific Modification | Details |
|---|---|---|
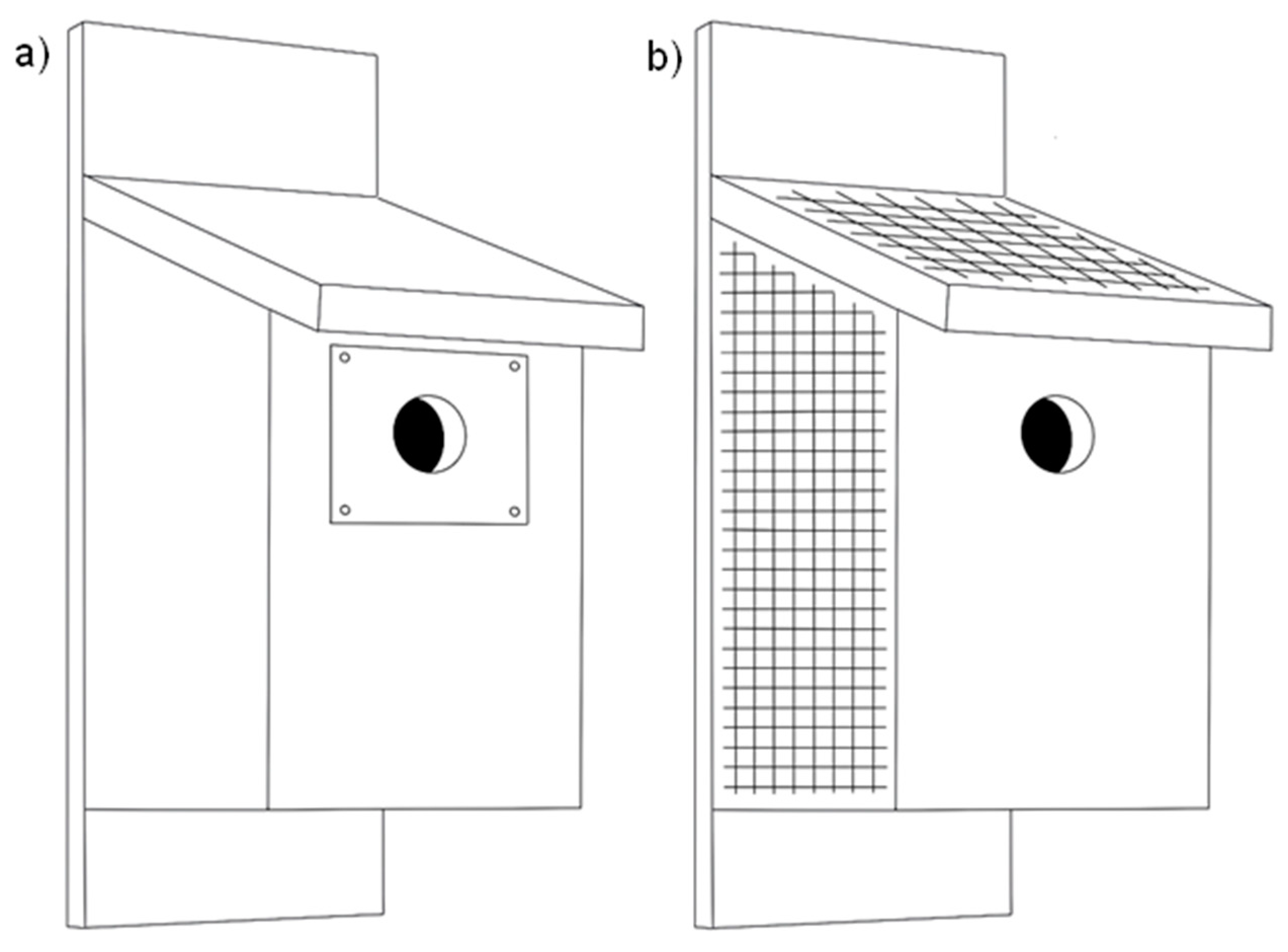
| Changes to wooden nestboxes | Metal plate around entrance | A thin metal plate is affixed to a wooden nestbox around the entrance hole to prevent mammalian predators using their teeth to enlarge the hole. |
| Wire mesh enclosing nestbox | Panels of a wooden nestbox are covered in galvanised wire mesh to reduce predation from predators penetrating the sides of a box. | |
| Deep nestbox | Design is very similar to a typical wooden nestbox but vertically elongated, increasing the distance between the entrance hole and nest cup; reduces risk of a predator reaching through the entrance to access eggs or chicks. | |
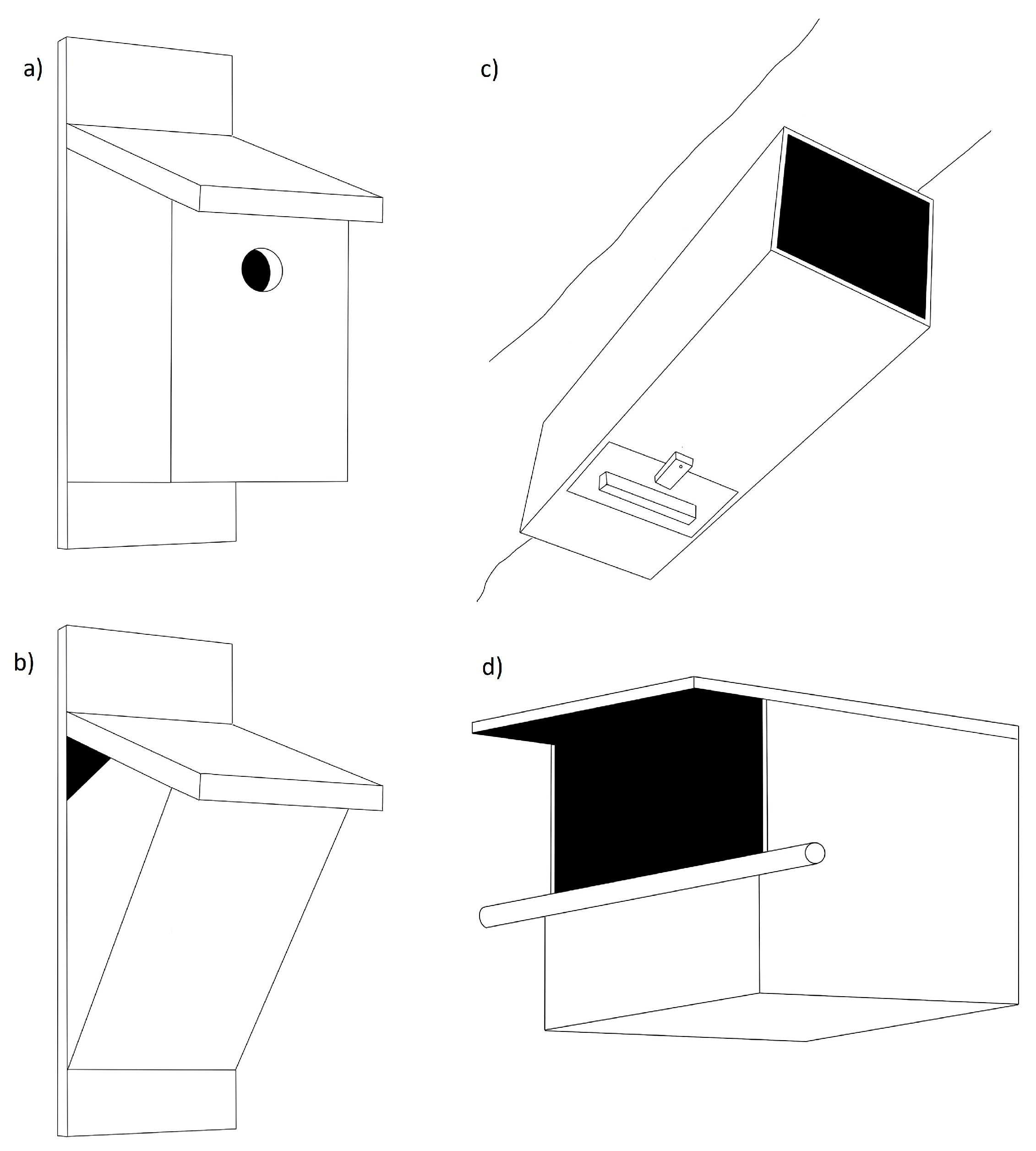
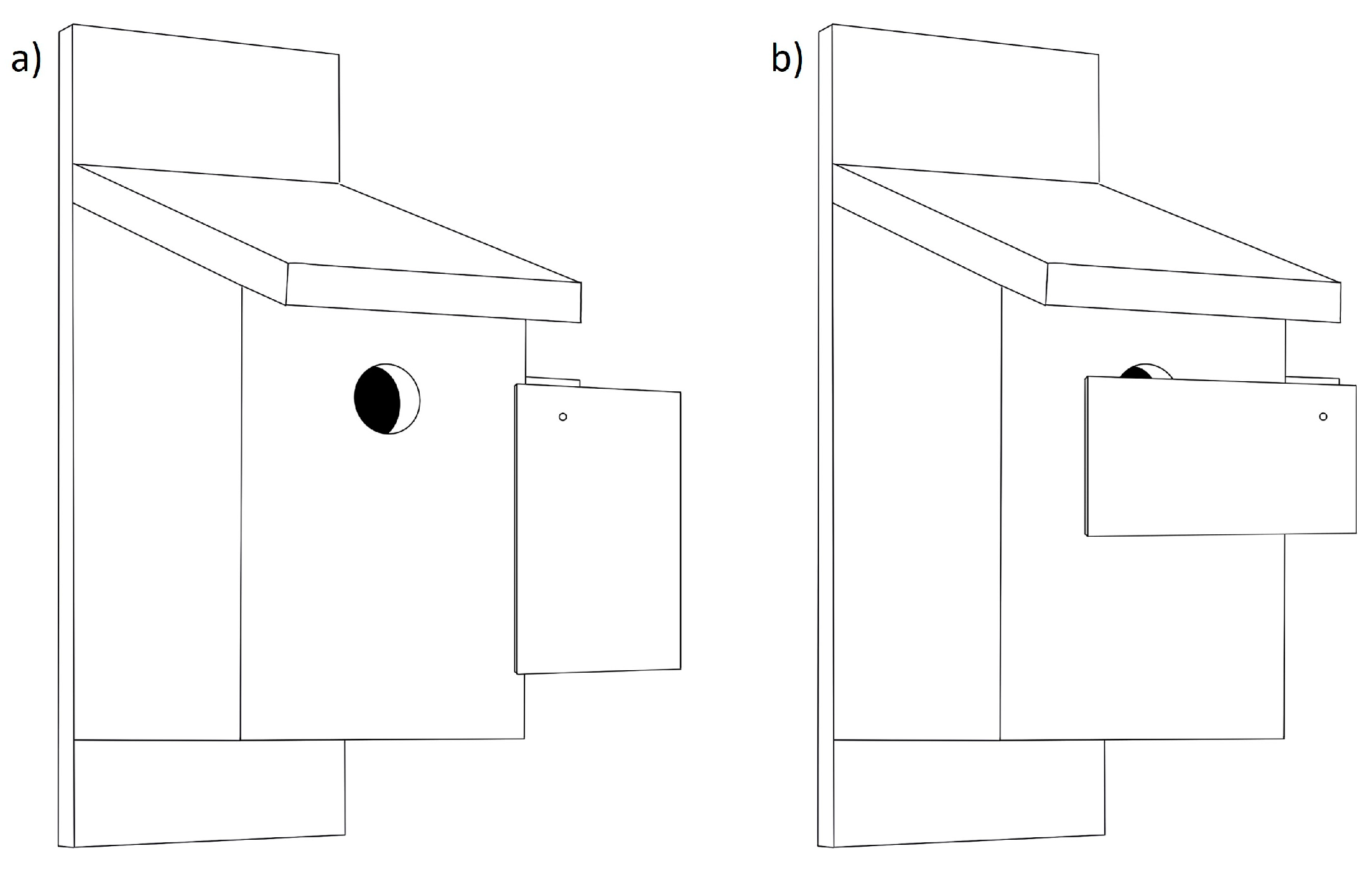
| Guardian, hole extender, Noel guard | A plastic, wooden, or wire structure attached to the outside of the nestbox to create an entrance tunnel, thereby increasing the distance from entrance to nest cup. | |
| Internal ledges below entrance | An internal modification of a small ledge inside the nestbox below the entrance hole to make it harder for mammalian predators to reach nestbox contents. | |
| Changes to nestbox materials | Woodcrete nestboxes | A mixture of cement and sawdust (“woodcrete”) that is easily moulded to produce nestboxes of various shapes and sizes—these tough nestboxes are much less likely to be penetrated or suffer hole enlargement by predators. |
| Technological solutions | Photosensitive shutters | A novel anti-predator strategy to prevent nocturnal predators entering nestboxes via a shutter over the entrance hole that closes in low light conditions. |
| Changes to nestbox positioning or replacement schedule | Baffles | Passive barriers placed below a nestbox to prevent climbing predators from reaching the nestbox; baffles are usually a stovepipe or a cone design. |
| Regular replacement of nestboxes | Wooden nestboxes decay over time and become more susceptible to penetration or hole enlargement as they age. Regular replacement can reduce this risk. | |
| Regular movement of nestboxes | Predators can use spatial memory to locate nestboxes that they have successfully depredated previously. Periodic movement of nestboxes can reduce this risk. |
| Intervention | Hidden Costs | Unanswered Questions | |
|---|---|---|---|
| Guardians, Noel guards, hole extenders | Potential decreases in provisioning rates as adults always need to enter the nest chamber rather than feeding older chicks from the entrance. | → | Based on ad-hoc observations, so extent and significance of this is unclear. |
| Changes in the ventilation and light within the nest cavity. | → | Seemingly not investigated. | |
| Deep nestboxes | Additional material being added to nestbox, with possible implications on nest building costs and first egg date. Also, relationships between nest volume and ectoparasite load. | → | Cost of extra material largely speculative and needs empirical quantification. Optimal box depth for ensuring that any extra nesting material will not decrease the (theoretically larger) distance between nest cup and entrance required. |
| Changes in the ventilation and light within the nest cavity. | → | Seemingly not investigated. | |
| Woodcrete nestboxes | Increase in nestbox internal temperature can have negative impacts on chick growth, especially when ambient temperatures are high or nestbox is exposed to sunlight for extended periods (including in gardens or open habitats). | → | Trade-offs between predation prevention and microclimate studied in ground-nesting species such as Piping Plover (Charadrius melodus) and Hoopoe Lark (Alaemon alaudipes) [165,166], but not in secondary cavity-nesting species. Critical temperature thresholds are thus unclear, as is the magnitude of benefits and costs. |
| Increase in humidity with possible implications for nest microbial load. | → | Seemingly not investigated, despite links between nestbox temperature, pathogenic microbial loads, and chick health in wooden nestboxes (e.g., [133]). | |
| Potentially more intense smell could increase predation from species that can enter cavity directly. | → | Investigated for snake predation of Blue and Great Tit nests in central Spain [123]; unclear if risk was site- or predator- specific or indicative of widespread issue. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcus, J.M.; Hart, A.G.; Goodenough, A.E. Predator-Proofing Avian Nestboxes: A Review of Interventions, Opportunities, and Challenges. Birds 2024, 5, 1-23. https://doi.org/10.3390/birds5010001
Marcus JM, Hart AG, Goodenough AE. Predator-Proofing Avian Nestboxes: A Review of Interventions, Opportunities, and Challenges. Birds. 2024; 5(1):1-23. https://doi.org/10.3390/birds5010001
Chicago/Turabian StyleMarcus, Joseph M., Adam G. Hart, and Anne E. Goodenough. 2024. "Predator-Proofing Avian Nestboxes: A Review of Interventions, Opportunities, and Challenges" Birds 5, no. 1: 1-23. https://doi.org/10.3390/birds5010001