
Defining Collision Risk: Lesser Flamingo Phoeniconaias minor Power Line Collision Sensitivity and Exposure for Proactive Mitigation
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
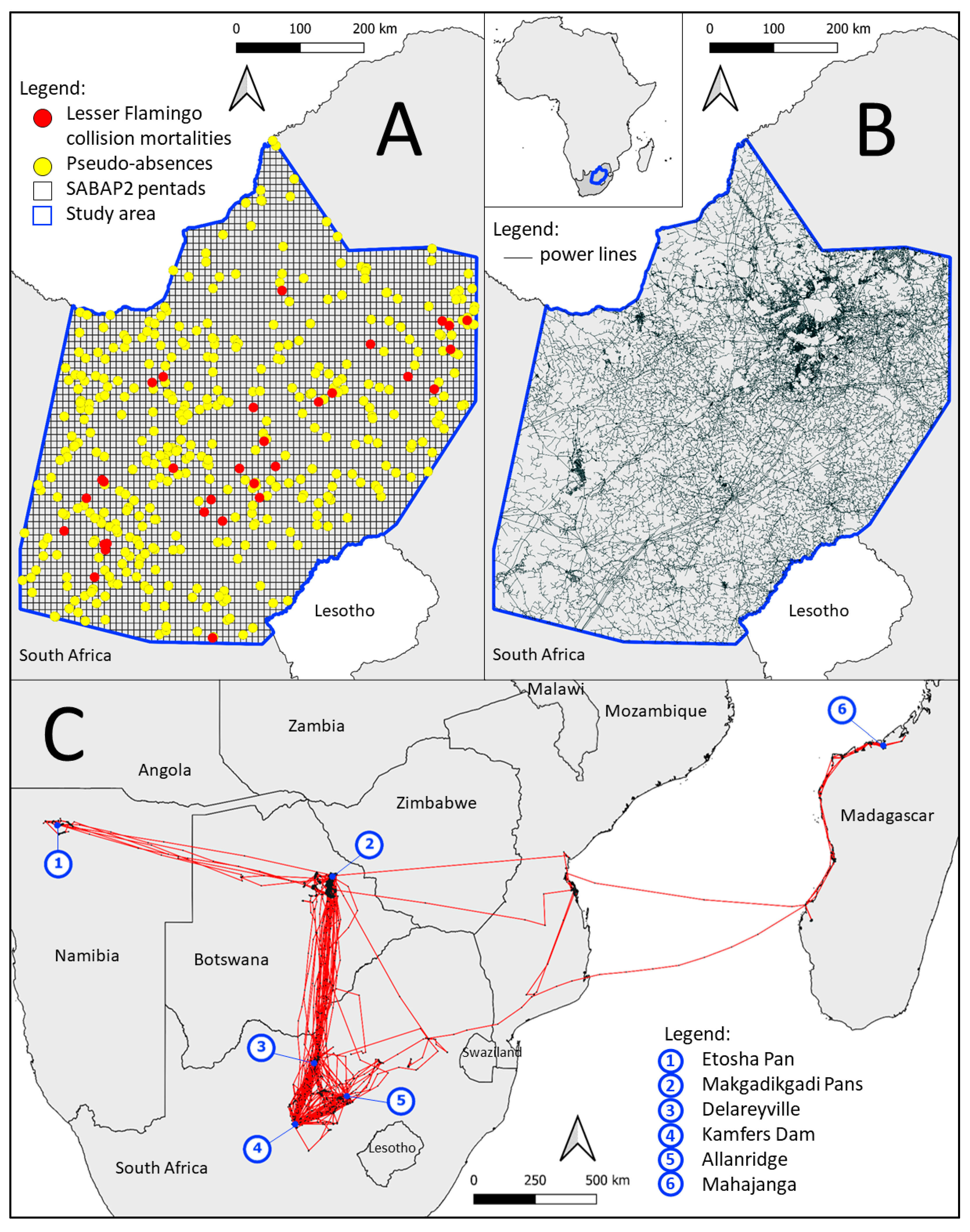
2.1. Study Area
2.2. Data Collection
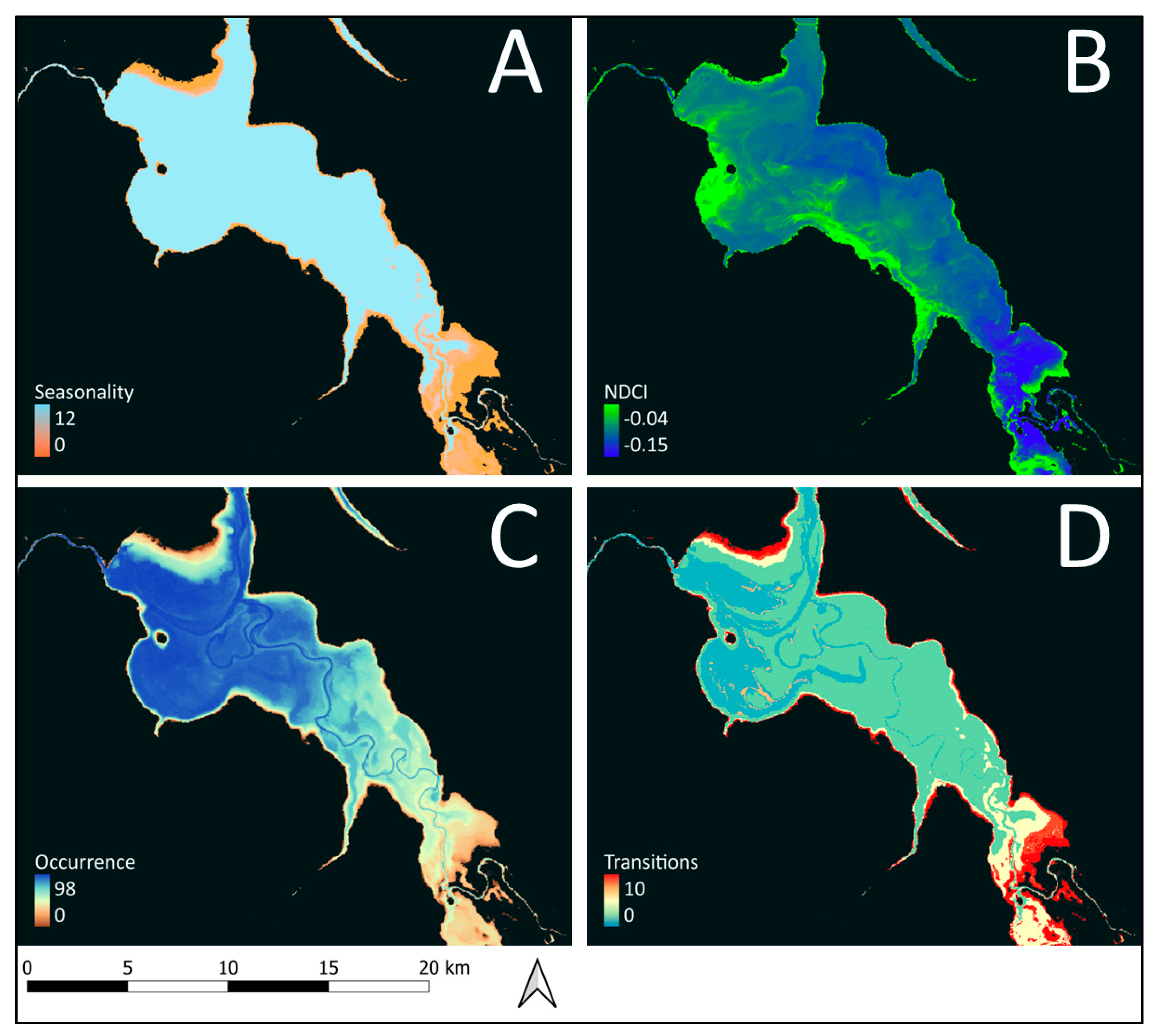
2.2.1. Habitat Suitability Variables
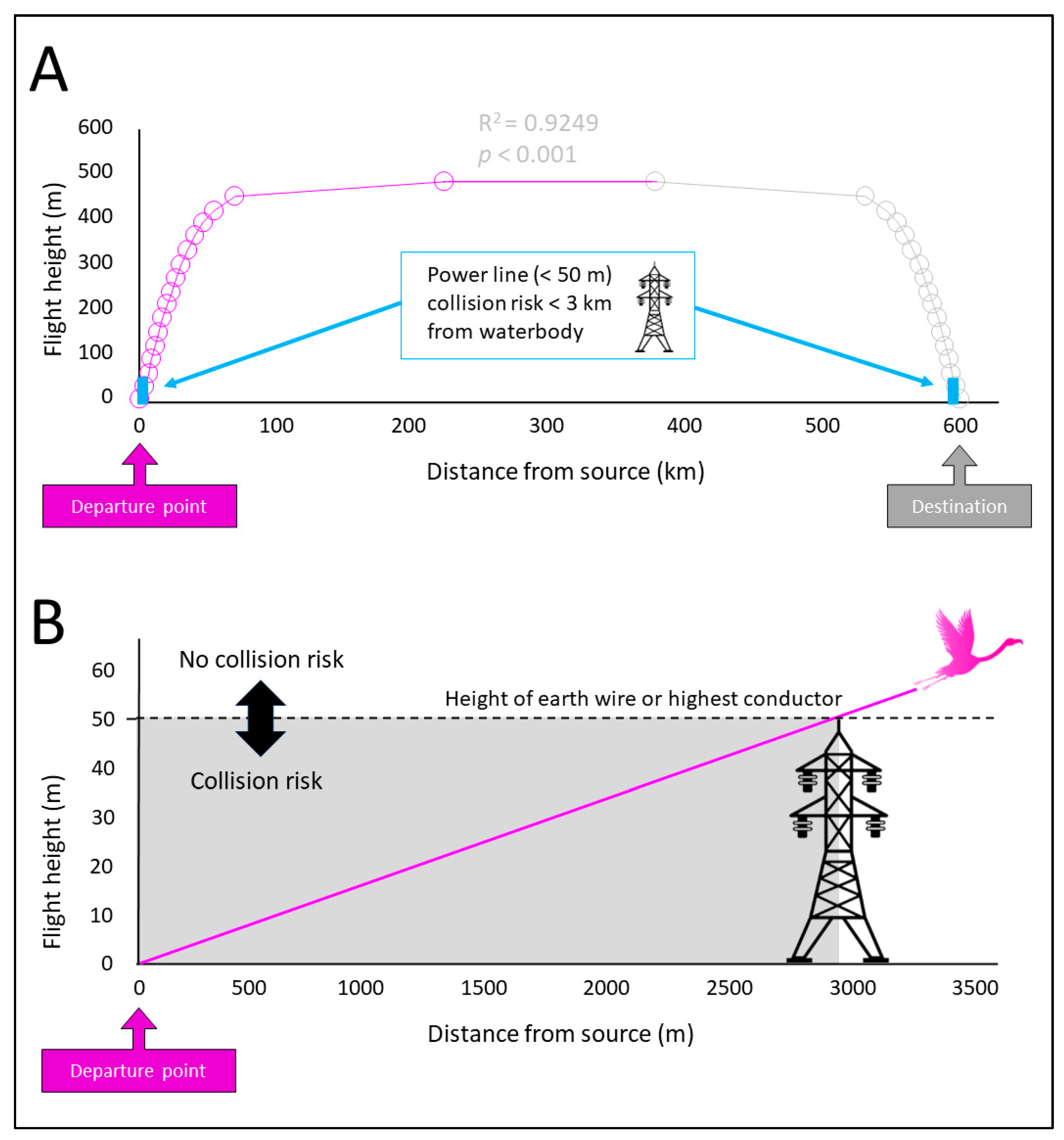
2.2.2. Species Exposure Variables
2.2.3. Threat Exposure Variables
2.3. Data Analysis
2.3.1. Habitat Suitability Index
2.3.2. Collision Risk Models
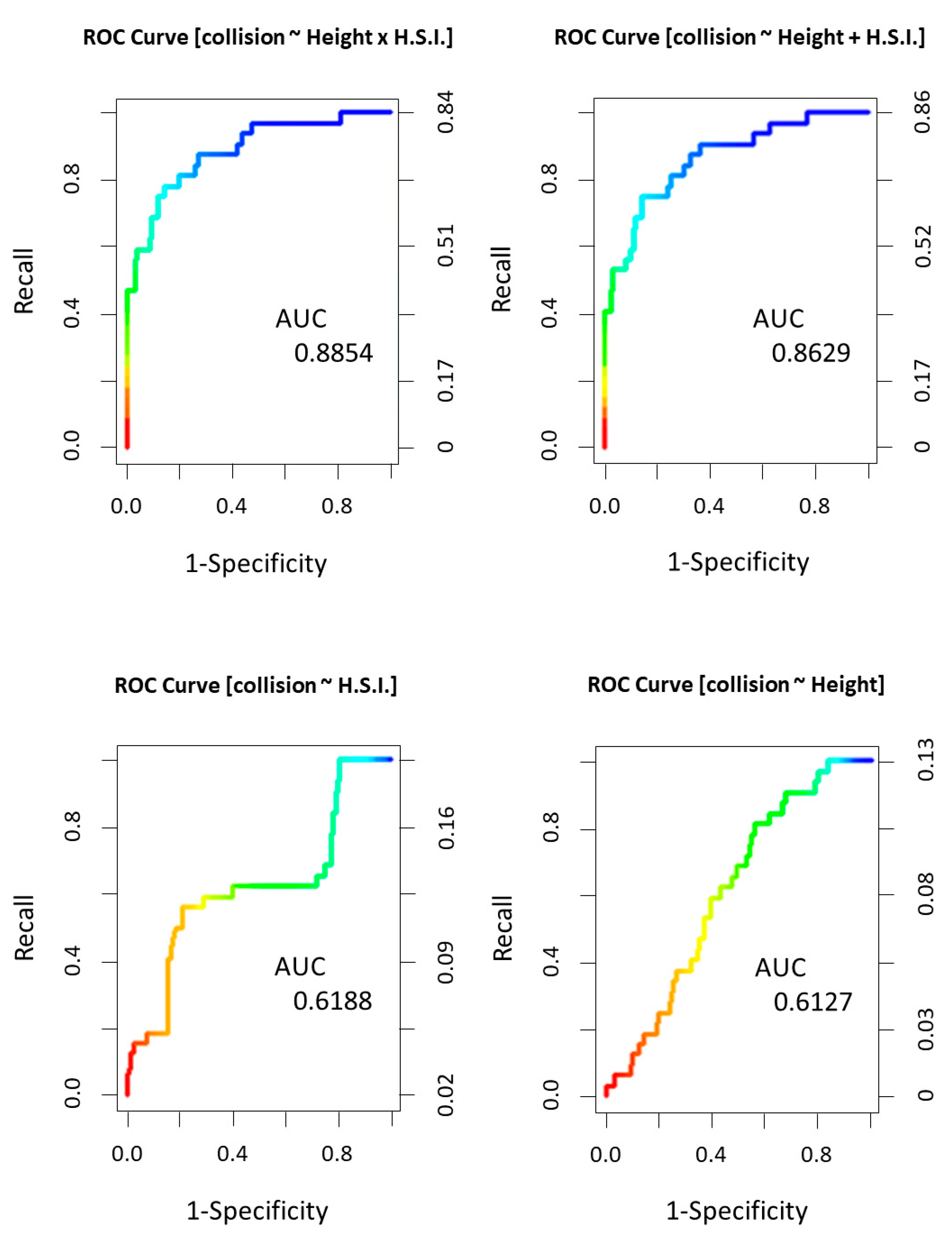
2.3.3. Model Validation
3. Results
3.1. Lesser Flamingo Habitat Suitability
3.2. Species Exposure
3.3. Multivariate Collision Risk Models
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensitivity | Exposure | ||||||
|---|---|---|---|---|---|---|---|
| Species Sensitivity | Habitat Suitability | Species Exposure | Threat Exposure | ||||
| Flight behavior | Suitability | Occurrence | Height | ||||
| Nocturnal habits 1 | Food 3 | Extent 4 | Tower height 3 | ||||
| Depth 2 | |||||||
| Flight dynamics | Abundance | Distance | |||||
| Wing-loading 1 | Availability | Reporting rate 3 | From water 3 | ||||
| Mass 1 | Water recurrence 4 | Individuals/ha 2 | From suitable habitat 3 | ||||
| Flight aspect 1 | Water seasonality 4 | ||||||
| Water transition 4 | Movements | ||||||
| Vision | Flight height 3 | ||||||
| Binocular vision 1 | Occurrence | ||||||
| Color range 1 | Water occurrence 4 | ||||||
Appendix B
| Scene Path | |||||
|---|---|---|---|---|---|
| 169 | 170 | 171 | 172 | ||
| - | - | - | - | ||
| Scene Row | 077 | - | - | LC08_L1TP_171077 | LC08_L1TP_172077 |
| 078 | LC08_L1TP_169078 | LC08_L1TP_170078 | LC08_L1TP_171078 | LC08_L1TP_172078 | |
| 079 | - | LC08_L1TP_170079 | LC08_L1TP_171079 | LC08_L1TP_172079 | |
| 080 | - | - | LC08_L1TP_171080 | LC08_L1TP_172080 | |
References
- Jenkins, A.R.; Smallie, J.J.; Diamond, M. Avian collisions with power lines: A global review of causes and mitigation with a South African perspective. Bird Conserv. Int. 2010, 20, 263–278. [Google Scholar] [CrossRef]
- Bernardino, J.; Bevanger, K.; Barrientos, R.; Dwyer, J.F.; Marques, A.T.; Martins, R.C.; Shaw, J.M.; Silva, J.P.; Moreira, F. Bird collisions with power lines: State of the art and priority areas for research. Biol. Conserv. 2018, 222, 1–13. [Google Scholar] [CrossRef]
- Dwyer, J.F.; Harness, R.E.; Donohue, K. Predictive model of avian electrocution risk on overhead power lines. Conserv. Biol. 2014, 28, 159–168. [Google Scholar] [CrossRef]
- Silva, J.P.; Palmeirim, J.M.; Alcazar, R.; Correia, R.; Delgado, A.; Moreira, F. A spatially explicit approach to assess the collision risk between birds and overhead power lines: A case study with the little bustard. Biol. Conserv. 2014, 170, 256–263. [Google Scholar] [CrossRef]
- Heck, N. A Landscape-Scale Model to Predict the Risk of Bird Collisions with Electric Power Transmission Lines in Alberta. Master Thesis, University of Calgary, Alberta, Canada, 2017. [Google Scholar]
- Shaw, J.M.; Jenkins, A.R.; Smallie, J.J.; Ryan, P.G. Modelling power-line collision risk for the Blue Crane Anthropoides paradiseus in South Africa. Ibis 2010, 152, 590–599. [Google Scholar] [CrossRef]
- Pretorius, M.D.; Hoogstad, C. An Avian Red-List Sensitivity Map Relevant to Eskom Distribution Power Lines in South Africa; Report no. RES/RR/15/1785097; Eskom Holdings SOC Ltd.: Johannesburg, South Africa, 2016. [Google Scholar]
- Bernstein, P.L. Against the Gods: The Remarkable Story of Risk; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Hertz, D.B.; Thomas, H. Risk Analysis and its Applications; Wiley: Chichester, UK, 1983. [Google Scholar]
- Kirkland, L.-H.; Thompson, D. Environmental Management Systems. In Tools for Environmental Management: A Practical Introduction and Guide; Thompson, D., Ed.; New Society Publishers: Gabriola Island, BC, Canada, 2002; pp. 19–42. [Google Scholar]
- Holton, G.A. Defining risk. Financ. Anal. J. 2004, 60, 19–25. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Gaston, K.J. Extrinsic factors and the population sizes of threatened birds. Ecol. Lett. 2002, 5, 568–576. [Google Scholar] [CrossRef]
- Dickinson, M.G.; Orme, C.D.L.; Suttle, K.B.; Mace, G.M. Separating sensitivity from exposure in assessing extinction risk from climate change. Sci. Rep. 2014, 4, 6898. [Google Scholar] [CrossRef] [PubMed]
- Burger, J. A risk assessment for lead in birds. J. Toxicol. Env. Health 1995, 45, 369–396. [Google Scholar] [CrossRef]
- Fischer, D.O.; Blomberg, S.P.; Owens, I.P.F. Extrinsic versus intrinsic factors in the decline and extinction of Australian marsupials. Proc. R. Soc. B: Biol. Sci. 2003, 270, 1801–1808. [Google Scholar] [CrossRef]
- Gauld, J.G.; Silva, J.P.; Atkinson, P.W.; Record, P.; Acácio, M.; Arkumarev, V.; Blas, J.; Bouten, W.; Burton, N.; Catry, I.; et al. Hotspots in the grid: Avian sensitivity and vulnerability to collision risk from energy infrastructure interactions in Europe and North Africa. J. Appl. Ecol. 2022, 59, 1496–1512. [Google Scholar] [CrossRef]
- Desholm, M.; Kahlert, J. Avian collision risk at an offshore wind farm. Biol. Lett. 2005, 1, 296–298. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Rehfisch, M.R.; Fox, A.D.; Desholm, M.; Anthony, S.J. The effect of avoidance rates on bird mortality predictions made by wind turbine collision risk models. Ibis 2006, 148, 198–202. [Google Scholar] [CrossRef]
- Krijgsveld, K.L.; Akershoek, K.; Schenk, F.; Dijk, F.; Dirksen, S. Collision risk of birds with modern large wind turbines. Ardea 2009, 97, 357–366. [Google Scholar] [CrossRef]
- D’Amico, M.; Martins, R.C.; Álvarez-Martínez, J.M.; Porto, M.; Barrientos, R.; Moreira, F. Bird collisions with power lines: Prioritizing species and areas by estimating potential population-level impacts. Divers. Distrib. 2019, 25, 975–982. [Google Scholar] [CrossRef]
- Martin, G.R.; Shaw, J.M. Bird collisions with power lines: Failing to see the way ahead? Biol. Conserv. 2010, 143, 2695–2702. [Google Scholar] [CrossRef]
- Bevanger, K. Bird interactions with utility structures: Collision and electrocution, causes and mitigation measures. Ibis 1994, 136, 412–425. [Google Scholar] [CrossRef]
- Janss, G.F.E. Avian mortality from power lines: A morphologic approach of a species-specific mortality. Biol. Conserv. 2000, 95, 353–359. [Google Scholar] [CrossRef]
- Shaw, J.M.; Reid, T.A.; Gibbons, B.K.; Pretorius, M.D.; Jenkins, A.R.; Visagie, R.; Michael, M.D.; Ryan, P.G. A large-scale experiment demonstrates that line marking reduces power line collision mortality for large terrestrial birds, but not bustards, in the Karoo, South Africa. Condor 2021, 123, duaa067. [Google Scholar] [CrossRef]
- Purvis, A.; Gittleman, J.L.; Cowlishaw, G.; Mace, G.M. Predicting extinction risk in declining species. Proc. R. Soc. B Biol. Sci. 2000, 267, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, J.; Moreira, F.; Franco, A.M.; Soriano-Redondo, A.; Acacio, M.; Gauld, J.; Rego, F.C.; Silva, J.P.; Catry, I. Flight altitudes of a soaring bird suggest landfill sites as power line collision hotspots. J. Environ. Manag. 2021, 294, 113–149. [Google Scholar] [CrossRef]
- Gustin, M.; Giglio, G.; Pellegrino, S.C.; Frassanito, A.; Ferrarini, A. Nocturnal flights lead to collision risk with power lines and wind farms in Lesser Kestrels: A preliminary assessment through GPS tracking. Comput. Ecol. Softw. 2018, 8, 15–22. [Google Scholar]
- Rollan, À.; Real, J.; Bosch, R.; Tintó, A.; Hernández-Matías, A. Modelling the risk of collision with power lines in Bonelli’s Eagle Hieraaetus fasciatus and its conservation implications. Bird Conserv. Int. 2010, 20, 279–294. [Google Scholar] [CrossRef]
- Reid, T.; Kruger, S.; Whitfield, D.P.; Amar, A. Using spatial analyses of bearded vulture movements in southern Africa to inform wind turbine placement. J. Appl. Ecol. 2015, 52, 881–892. [Google Scholar] [CrossRef]
- Anderson, M.D. Lesser Flamingo Phoeniconaias minor. In The 2015 Eskom Red Data Book of Birds of South Africa, Lesotho and Swaziland; Taylor, M.R., Peacock, F., Wanless, R.M., Eds.; BirdLife South Africa: Johannesburg, South Africa, 2015; pp. 305–307. [Google Scholar]
- Childress, B.; Harper, D.; Hughes, B.; Van der Bossche, W.; Berthold, P.; Querner, U. Satellite tracking Lesser Flamingo movements in the Rift Valley, East Africa: Pilot study report. Ostrich 2004, 75, 57–65. [Google Scholar] [CrossRef]
- Childress, B.; Harper, D.; Ferris, C. Sex determination in the Lesser Flamingo (Phoenicopterus minor) using morphological measurements. Ostrich 2005, 76, 148–153. [Google Scholar] [CrossRef]
- Pretorius, M.D.; Leeuwner, L.; Tate, G.J.; Botha, A.; Michael, M.D.; Durgapersad, K.; Chetty, K. Movement patterns of lesser flamingos Phoeniconaias minor: Nomadism or partial migration? Wildl. Biol. 2020, 2020, wlb.00728. [Google Scholar] [CrossRef]
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library. R Package Version 1.4-6. 2019. Available online: https://CRAN.R-project.org/package=rgdal (accessed on 31 January 2020).
- Kellner, C.J.; Brawn, J.D.; Karr, J.R. What is habitat suitability and how should it be measured? In Wildlife 2001: Populations; McCullough, D.R., Barrett, R.H., Eds.; Elsevier Applied Science: London, UK; New York, NY, USA, 1992; pp. 476–488. [Google Scholar]
- Verner, J.; Morrison, M.L.; Ralph, C.J. Wildlife 2000: Modeling Habitat Relationships of Terrestrial Vertebrates; University of Wisconsin Press: Madison, WI, USA, 1986. [Google Scholar]
- Tebbs, E.J.; Remedios, J.J.; Harper, D.M. Remote sensing of chlorophyll-a as a measure of cyanobacterial biomass in Lake Bogoria, a hypertrophic, saline–alkaline, flamingo lake, using Landsat ETM+. Remote Sens. Environ. 2013, 135, 92–106. [Google Scholar] [CrossRef]
- Tebbs, E.J.; Remedios, J.J.; Avery, S.T.; Harper, D.M. Remote sensing the hydrological variability of Tanzania’s Lake Natron, a vital Lesser Flamingo breeding site under threat. Ecohydrol. Hydrobiol. 2013, 13, 148–158. [Google Scholar] [CrossRef]
- Krienitz, L.; Krienitz, D.; Dadheech, P.K.; Hübener, T.; Kotut, K.; Luo, W.; Teubner, K.; Versfeld, W.D. Food algae for Lesser Flamingos: A stocktaking. Hydrobiologia 2016, 775, 21. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.R. Normalized difference Chlorophyll index: A novel model for remote estimation of Chlorophyll-a concentration in turbid productive waters. Remote Sens. Environ. 2012, 117, 394–406. [Google Scholar] [CrossRef]
- Pekel, J.-F.; Cottam, A.; Gorelick, N.; Belward, A. Global Surface Water Explorer Dataset. European Commission, Joint Research Centre (JRC). 2017. Available online: http://data.europa.eu/89h/jrc-gswe-global-surface-water-explorer-v1 (accessed on 20 February 2019).
- Pekel, J.-F.; Cottam, A.; Gorelick, N.; Belward, A. High-resolution mapping of global surface water and its long-term changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.T.; Altwegg, R.E.S.; Barnard, P. Estimating conservation metrics from atlas data: The case of southern African endemic birds. Bird Conserv. Int. 2017, 27, 323–336. [Google Scholar] [CrossRef]
- Lee, A.T.; Fleming, C.; Wright, D.R. Modelling bird atlas reporting rate as a function of density in the southern Karoo, South Africa. Ostrich 2018, 89, 363–372. [Google Scholar] [CrossRef]
- Peterson, B.G.; Carl, P. Performance Analytics: Econometric Tools for Performance and Risk Analysis. R Package Version 1.5.2. 2018. Available online: https://CRAN.R-project.org/package=PerformanceAnalytics (accessed on 30 November 2018).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.3-2. 2017. Available online: https://cran.pau.edu.tr/web/packages/AICcmodavg/AICcmodavg.pdf (accessed on 31 January 2020).
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Kuhn, M. Caret: Classification and Regression Training. R Package Version 6.0-86. 2020. Available online: https://CRAN.R-project.org/package=caret (accessed on 1 April 2020).
- Martin, G.R. Understanding bird collisions with man-made objects: A sensory ecology approach. Ibis 2011, 153, 239–254. [Google Scholar] [CrossRef]
- Martin, G.R.; Jarrett, N.; Tovey, P.; White, C.R. Visual fields in Flamingos: Chick-feeding versus filter-feeding. Naturwissenschaften 2005, 92, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Pannucio, M.; Dell-Omo, G.; Bogliani, G.; Sapir, N. Migrating birds avoid flying through fog and low clouds. Int. J. Biometeorol. 2019, 63, 231–239. [Google Scholar] [CrossRef]
- Pretorius, M.D.; Hoogstad, C. Evaluating the Effectiveness of the ‘OWL Device’, a Bird Flight Diverter For Night-Flying Birds; Report no. RES/RR/16/1841861; Eskom Holdings SOC Ltd.: Johannesburg, South Africa, 2017. [Google Scholar]




| Model | K | AICc | ∆AICc | w | cum.w |
|---|---|---|---|---|---|
| Visited ~ ∆SWO + NDCI + NDCImax | 4 | 213.990 | 0.000 | 0.533 | 0.533 |
| Visited ~ Water recurrence + NDCI + NDCImax | 4 | 214.540 | 0.540 | 0.407 | 0.939 |
| Visited ~ NDCI + NDCImax | 3 | 218.440 | 4.450 | 0.058 | 0.997 |
| Coefficients | Estimate | SE | z Value | p |
|---|---|---|---|---|
| (Intercept) | −3.923 | 1.331 | −2.947 | 0.003 |
| Flight height | −0.014 | 0.005 | −2.743 | 0.006 |
| Habitat suitability | 1.763 | 0.643 | 2.740 | 0.006 |
| Reporting rate | 0.013 | 0.009 | 1.36106 | 0.173 |
| Cable height | −0.061 | 0.055 | −1.11236 | 0.266 |
| Model | K | AICc | ∆AICc | w | cum.w |
|---|---|---|---|---|---|
| Collision ~ flight height x habitat suitability | 4 | 209.08 | 0 | 0.73 | 0.73 |
| Collision ~ flight height + habitat suitability | 3 | 211.12 | 2.04 | 0.26 | 1 |
| Collision ~ habitat suitability | 2 | 220.13 | 11.05 | 0 | 1 |
| Collision ~ flight height | 2 | 221.34 | 12.26 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pretorius, M.D.; Galloway-Griesel, T.L.; Leeuwner, L.; Michael, M.D.; Durgapersad, K.; Chetty, K. Defining Collision Risk: Lesser Flamingo Phoeniconaias minor Power Line Collision Sensitivity and Exposure for Proactive Mitigation. Birds 2023, 4, 315-329. https://doi.org/10.3390/birds4040027
Pretorius MD, Galloway-Griesel TL, Leeuwner L, Michael MD, Durgapersad K, Chetty K. Defining Collision Risk: Lesser Flamingo Phoeniconaias minor Power Line Collision Sensitivity and Exposure for Proactive Mitigation. Birds. 2023; 4(4):315-329. https://doi.org/10.3390/birds4040027
Chicago/Turabian StylePretorius, Mattheuns D., Tamsyn L. Galloway-Griesel, Lourens Leeuwner, Michael D. Michael, Kaajial Durgapersad, and Kishaylin Chetty. 2023. "Defining Collision Risk: Lesser Flamingo Phoeniconaias minor Power Line Collision Sensitivity and Exposure for Proactive Mitigation" Birds 4, no. 4: 315-329. https://doi.org/10.3390/birds4040027