1. Introduction
While zoo visitors often seek interactions with animals that they come to learn about and enjoy [
1], it is not clear if zoo animals seek and/or benefit from the presence of visitors. Zoological institutions strive to improve the well-being of animals in human care and contribute to the growing field of visitor studies. To date positive, neutral, as well as negative visitor effects have been reported [
2,
3,
4,
5]. For example, time spent playing in Diana monkeys (
Cercopithecus diana) was found to increase with a greater number of visitors [
2] and play is associated with positive welfare states [
6]. In contrast, time slender-tailed meerkats (
Suricata suricatta) spent vigilant or looking at visitors, as well as distance between meerkats and visitors, did not vary in response to reduced visitor noise and meerkat-directed visitor activity [
3] which is interpreted as a neutral effect. Soemmerring’s gazelles (
Gazella soemmerringii) kept in enclosures with greater exposure to zoo visitors had more frequent gazelle-gazelle agonistic interactions [
7], an undesirable or negative effect. Results of visitor studies clearly vary with the species observed and likely vary as a function of housing provided as well as individual histories and individual characteristics of the particular animals studied [
5]. When doing research in the zoo uncontrolled extraneous factors may influence outcomes. Numerous studies have explored the effects of visitor density on animal behavior but in many cases visitor density is associated with time of day and/or time of year. Differences attributed to visitor density may be confounded with these and other variables such as animal care routines, feeding times, and presence of enrichment. Studies of visitor effects are often carried out over relatively brief periods and differences resulting from increases in maturity of animals are rarely considered.
The presence of visitors is one of many variables that may alter the behavior of zoo animals. Additions to animals housed together, whether due to births or introductions of incoming animals, are likely to bring social changes. Observations before, during, and following births and introductions expand our knowledge of behavior and provide information that can assist in animal management. Such events were anticipated during winter of 2019/2020 in each of the two western lowland gorilla groups (Gorilla gorilla gorilla) housed at the Los Angeles Zoo. Studies to document changes in time spent in specific activities, habitat utilization, and frequency of social behavior in relation to zoo attendance, time of day, and increasing gorilla maturity were underway when COVID-19 was declared a pandemic in Los Angeles.
To reduce the spread of the virus, people were advised to restrict their outside activities. In response to pandemic conditions most zoos were closed to visitors for one or more periods of time. Zoo closures made it possible to assess differences in the behavior of zoo animals with and without visitors present. At the Los Angeles Zoo we continued gorilla observations, and, in this report, we include the presence or absence of visitors as one of the input variables to be assessed with respect to changes in gorilla behavior, including frequency of both affiliative and agonistic actions directed to gorillas and to zoo visitors.
2. Materials and Methods
Two groups of western lowland gorillas (
Gorilla gorilla gorilla) have been housed at the Los Angeles Zoo since 2007 and changes in the composition of both groups occurred during the winter of 2019/2020. A blackback male was introduced to the bachelor group of two older silverbacks and a female was born in the family group consisting of one older silverback and three adult females. Life-history parameters of gorillas are known and indicated that during the nineteen-month study the eleven-year-old male’s secondary sexual characteristics would continue to develop as he approached the age of a young silverback [
8,
9]. The infant female would be carried and nursed by her dam, but she would increasingly interact with other members of the family group [
10].
A total of eight gorillas were observed who ranged in age from newborn to 43 years at the start of the study; their ID codes, sex and birth years are shown in
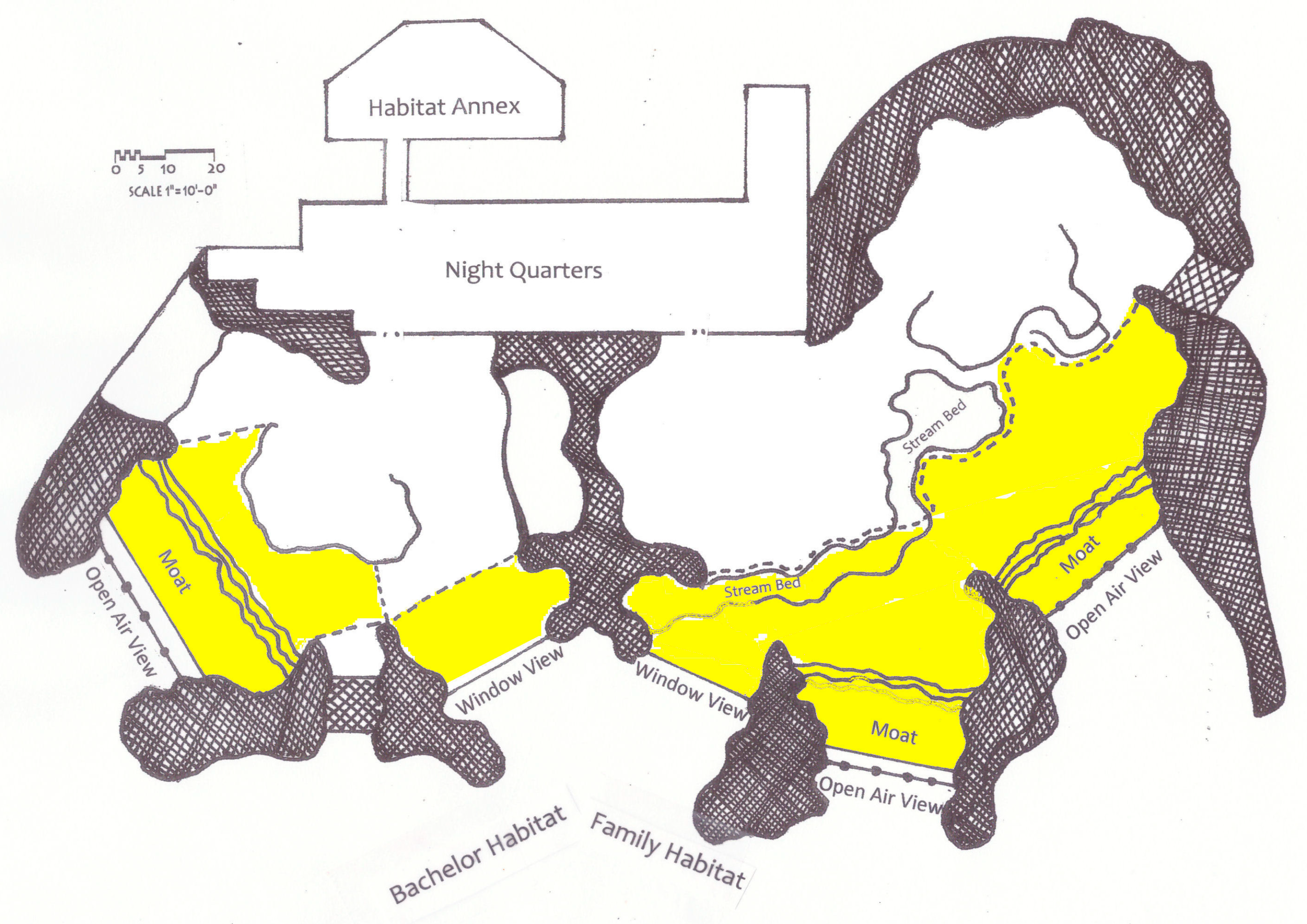
Table 1. All but the female born in 1976 were parent reared. During the day the gorillas occupy the outdoor multilevel habitat between 0945 and 1600. The bachelor and family groups reside in adjacent areas in the Campo Gorilla Reserve which is diagramed in
Figure 1. Usable outdoor gorilla space which can be viewed by visitors totals 14,963 sq. ft. The outdoor habitat utilized by the family group is approximately twice the size of that used by the bachelor group. Behind the indoor night-quarters there is an off-exhibit outdoor annex which provides an additional 858 sq. ft. of usable space for the bachelors who can access it from their outdoor multilevel habitat. At night the bachelors are housed separately; in the family group the oldest female is housed separately, other members of the family group can choose to rest together or apart.
The gorillas are fed three times each day. The morning meal, browse and enrichment items are spread across the multilevel outdoor habitat prior to 0945. The gorillas may be briefly brought into the night-quarters while the mid-day meal is scattered in their outdoor habitat. Alternatively, an extra portion may be included in the morning feed or tossed in while gorillas remain in the habitat. The evening meal is provided at 1630, after the gorillas have entered their night-quarters.
The study reported here covers a 19-month period and includes two periods of zoo closure which were preceded or followed by periods in which zoo visitors were present. In phase 1 (1 November 2019 through 12 March 2020) visitors were present; incoming male, HJ, was introduced to the bachelor group on 12 December and female A was born to N in the family group on 18 January. In phase 2 (13 March through 25 August 2020) visitors were absent. In phase 3 (26 August through 6 December 2020) visitors were present but limited to stay within the attendance range specified by the City of Los Angeles and Los Angeles County Department of Public Health who were closely monitoring the level of COVID-19. Date spans and maximum to be admitted per day in phase 3 follow: 26–30 August/1200, 31 August–31 October/1800, 1 November–6 December/2300. In phase 4 (7 December 2020 through 15 February 2021) visitors were absent. In phase 5 (16 February through 30 June 2021) visitors were present but attendance was limited to stay within the city and county specified ranges. Date spans and maximum to be admitted per day in phase 5 follow: 16 February–11 March/2400, 12 March–15 April/4400, 16 April–26 May/5000, 27 May–14 June/7700, 15–30 June/10,000. The number of focal observations of each gorilla during each of the 5 different visitor phases is shown in
Table 1.
All gorilla observations were made from visitor outdoor viewing areas between 0945 and 1545. On study days each gorilla who had access to the multi-level outdoor area was observed as the focal animal for a 15 min period. A focal observation started with sequential description of all social behavior directed to or from the focal gorilla. Potential recipients included neighboring gorillas and zoo visitors as well as other gorillas in the same habitat. Time markers sounded at 60 s intervals and the observer instantaneously recorded focal activity and habitat sector occupied, then continued with sequential recording [
11]. Definitions of behaviors recorded are given in
Table 2. Members of the bachelor group were observed for a total of 96.25 h and members of the family group for a total of 210.75 h.
Analysis of activity and location input variables included whether visitors could enter the zoo, sequential month of the study, zoo attendance on observation days, and hour at start of focal observation. Social interactions within bachelor groups and family groups are well known to differ in type and in frequency. When housed with other near-adult or adult males, bachelors show substantial non-contact and contact aggression which peaks in young silverbacks between the ages of 14 and 20 years [
12]. Occurrences of social behavior were analyzed separately for the two groups. All outcome variables were recorded as counts and analyzed using mixed-effects negative binomial regression (MENBREG). MENBREG in Stata 17.0 was utilized as it is suitable for non-normal distributions of count data, can incorporate repeated observations of the same individuals, and allows multiple input variables to be considered. In the analyses reported below repeated observations were nested in focal gorillas. To account for variation in amount of time a focal animal could be seen, visible activities instantaneously sampled were summed and used as the exposure variable for activity analyses in Stata. Number of minutes in which social interactions between an initiator and recipient could be documented was used as the exposure variable for social interaction analyses. As we would be assessing multiple input variables alpha was set at 0.01.
3. Results
3.1. Activity and Location
Regressions were done for the six instantaneously sampled measures described in
Table 2; results are shown in
Table 3. The hour at which the observation was started was significantly associated with three output variables: proportion of scans feeding and/or foraging, proportion of scans stationary or inactive, and proportion of scans in proximity to visitor viewing areas. At later start times the proportion of scans in foraging activity, as well as scans in proximity to visitor viewing areas, decreased while the proportion of scans in which the gorillas were stationary increased. In the morning, before gorillas had access to the outdoor areas, keepers scattered food throughout the habitat, including areas near visitor viewing sites. The gorillas spent the most time foraging when they first entered the outdoor habitat. As food was consumed foraging opportunities decreased, including in the areas proximate to visitor viewing. As a result, gorillas foraged less in later hours, were less likely to spend time in areas adjacent to visitor viewing, and tended to be less active.
Study month was also associated with three output variables: exploration/self-play, not visible, and time proximate to visitor viewing areas. In this study successive study months are associated with increasing maturity of the introduced bachelor and of the infant. As months progressed, the gorillas spent a greater proportion of scans in exploration/self-play, and a smaller proportion of scans not visible. They also spent a greater proportion of scans in proximity to visitor viewing areas.
Zoo closure was associated with just one instantaneously sampled outcome variable; the gorillas spent a greater proportion of scans proximate to visitor viewing areas when there were no visitors.
When observing it was clear that female E, who had been nursery-reared, utilized the display habitat differently from others. In contrast to the other family gorillas in the same habitat, she spent a substantially greater proportion of her time in sectors proximate to visitor viewing areas. During the 19-month study, E spent an average of 0.7635 (standard deviation = 0.3585) proportion of her scans in those areas in contrast to the other four gorillas in the same habitat who spent an average of 0.3549 (standard deviation = 0.37696) proportion of their scans there.
None of the input variables, including zoo attendance, showed a significant relationship to proportion of scans in self-directed activity which is often used as an indicator of stress [
13,
14,
15].
3.2. Social Interactions
Non-contact threats including tight lips, stiff stance, chest beat, bluff charge, object display, and chase were summed with contact aggression to obtain total occurrences of aggressive behavior. Brief contact, sustained contact, sexual contact, social examine/groom, and social play were summed to obtain total occurrences of affiliative behavior. Interactions among the three bachelors were more than six times as frequent as those among the family group of five gorillas.
Among the bachelor group an average of 1.3806 occurrences of aggressive behavior (standard deviation of 3.0935) and an average of 0.0382 occurrences of affiliative behaviors (standard deviation of 0.4676) took place during focal observations. Among the family group an average of 0.0241 occurrences of aggressive behavior (standard deviation of 0.1899) and 0.1509 occurrences of affiliative behavior (standard deviation of 0.4676) took place during focal observations.
There were few interactions between the two gorilla groups: During 96.25 bachelor focal observation hours we documented a total of 9 threats from a bachelor towards the family area and all occurred in the last phase of the study when bachelor aggression was markedly increasing (3 from H, the dominant male, and 6 from HJ, the introduced male). During 210.75 family focal observation hours no aggression was directed to the bachelors. No affiliative behavior was seen between the two gorilla groups.
The bachelors directed a total of 13 threats (3 from H, 6 from J, 4 from HJ) and showed no affiliative behavior to visitors. The family group gave 11 threats to visitors (5 from E, 4 from K, 2 from R) and, except for gorilla E, no affiliative behavior to visitors. In contrast to others in the family group, E directed a total of 122 affiliative behaviors to visitors.
MENBREG showed considerable differences between the two groups with respect to associations between input variables and within group social occurrence outcomes that can be viewed in
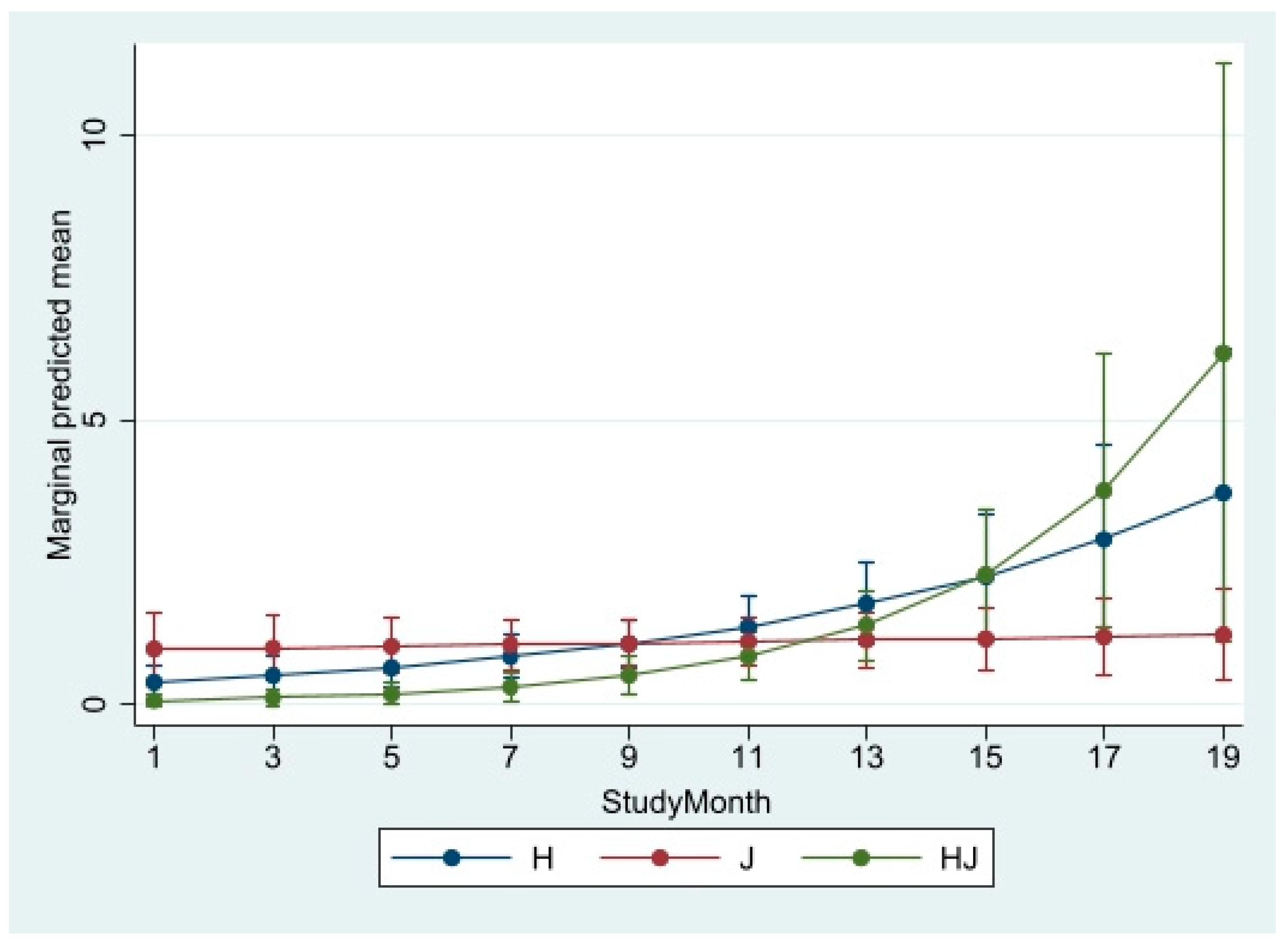
Table 4. For bachelors, progression in study months was positively associated with increased aggression (see
Figure 2). As months progressed, introduced male HJ was maturing and becoming more assertive. MENBREG also shows there was significantly more bachelor aggression when the zoo was open to visitors but less aggression when attendance was higher. The family group also showed less aggression with higher attendance and the association between affiliative behavior and attendance approached significance. That is, there was less affiliative behavior among family members when attendance was higher (
p = 0.016).
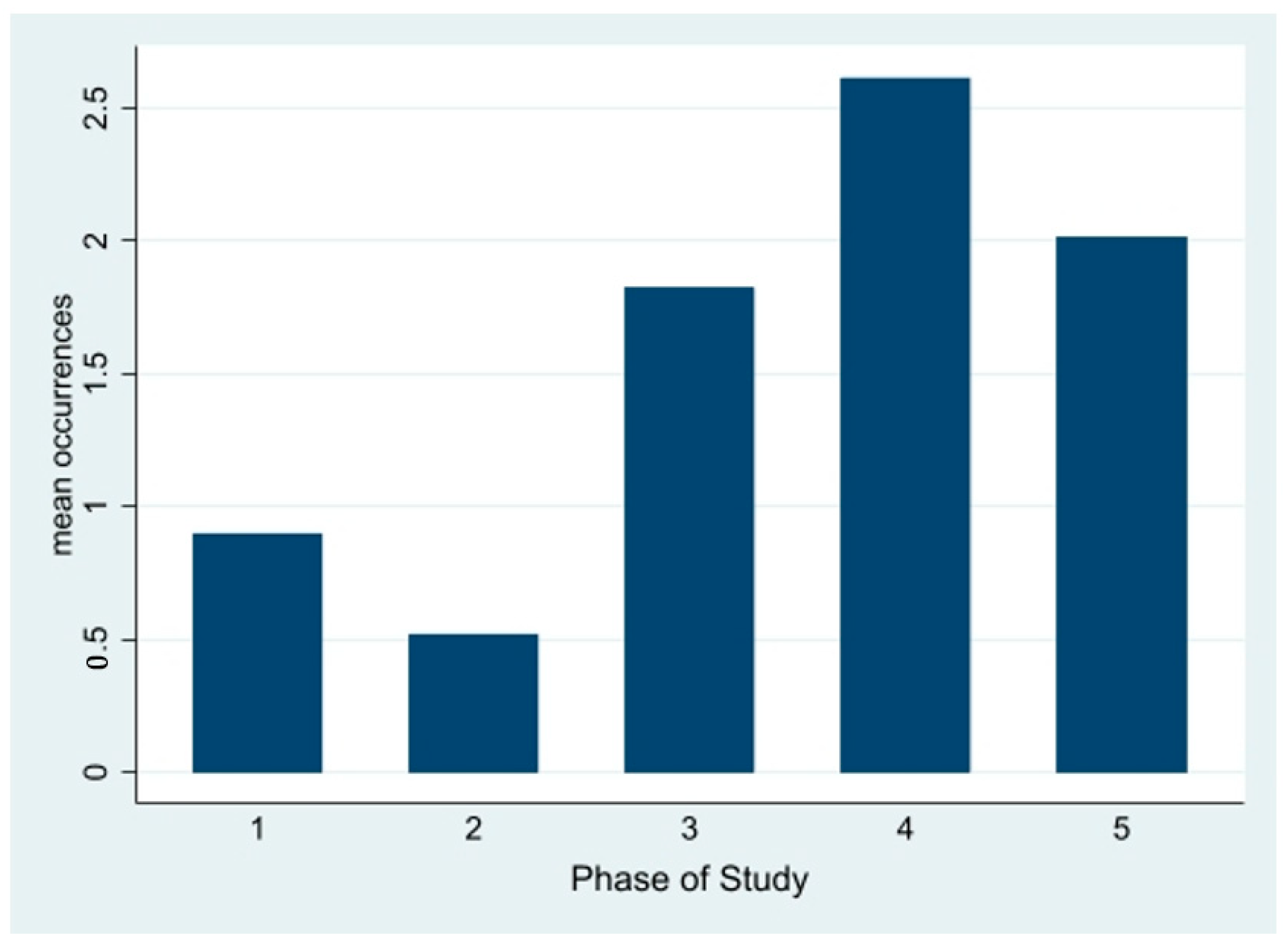
While there is a statistically significant association between zoo closure and lower bachelor aggression, looking at the level of aggression in
Figure 3 during each of the two closures separately makes it clear that closure does not cause the reduction in bachelor aggression. Over the course of the 19-month study, the lowest level of bachelor aggression that occurred was during the first closure while the highest level of aggression that occurred was during the second closure.
4. Discussion
In this study we focused on assessing possible effects of the closure on changes in gorilla behavior while looking at other factors that could also be contributing to changes. Just two significant relationships were found with zoo closure: more time was spent in areas proximate to visitor viewing when the zoo was closed to visitors and the level of aggression among the bachelors was lower. In fact, the level of bachelor aggression during the two closures was quite different, with low levels of aggression during the first closure but high levels of aggression during the second closure which was much briefer. Increases in aggression are characteristic of maturing blackbacks [
12,
16] and consistent with the difference in level of aggression among the bachelors during the successive closures. HJ had been introduced at 11 years of age and by the time of the second closure he had reached 12 years of age. Both
Figure 2 and
Figure 3 clearly show that aggression is increasing as HJ is maturing, a finding consistent with the literature.
A further indication that lack of visitors is unlikely to result in differences in gorilla social behavior is that, with the exception of nursery-reared gorilla E, few social behaviors were directed to visitors when the zoo was open. In short, we found just one change that is likely to have resulted from zoo closure, increased time in proximity to visitor viewing areas. This does suggest that viewing visitors is of interest to the gorillas and for this reason we have initiated a follow-up study in which visual attention is being recorded. A long-term study exploring the effects of zoo closure on the behavior of gorillas is underway at the Buffalo Zoo where researchers also report minimal effects [
17,
18].
Effects of zoo attendance or visitor density on behavior of gorillas residing at zoological institutions has been explored in a number of studies. Wells reports high visitor density encouraged more aggression, stereotypies and autogrooming [
19]. Lewis et al. report high visitor density resulted in increased locomotion and increased visitor vigilance [
20]. Stoinski et al., who assessed possible visitor effects in four gorilla groups, report that with high crowds, two groups showed increased stereotypies while the other two groups expressed fewer stereotypies. The investigators also found that only males showed more contact aggression [
21]. Kuhar, who assessed two groups, reports that with high crowds, gorillas were less visible and that the bachelor group showed increased aggression [
22]. Hashmi and Sullivan report high visitor density resulted in less time spent inactive, more time feeding, and no change in stereotypy [
23]. In this study less aggression among the bachelors as well as among the family group was associated with higher zoo attendance which had not previously been reported. However, a significant association does not imply causality. Prior to the pandemic the Los Angeles Zoo would host as many as 12,000 visitors on a single high attendance Saturday or Sunday; the average number of visitors admitted during phase 1 observation days was 3694. In order to maximize distance between visitors and minimize risk of disease transmission, when the zoo re-opened attendance was intentionally limited to 1200 visitors. Over the next four months the number of visitors who could enter was gradually raised to 2300. Average number of visitors during study days in this period, phase 3, was 1515. In December 2020, COVID-19 surged in Los Angeles and the zoo again closed to visitors. When the zoo reopened following the second closure, admission was initially limited to 2400 visitors. The number admitted was raised in increments to 10,000 by the end of June, 2021, when observations for this study came to a close. Average attendance during study days in phase 5 was 3405 which was still lower than the average attendance during study days in phase 1. Over the course of the study HJ was maturing and becoming more aggressive in the bachelor group. At the same time the infant in the family group was maturing and the adults were becoming less protective, engaging in more aggressive behavior than they had when the infant was younger. However, the number of visitors admitted had been restricted due to COVID-19 policies. Gorilla aggression and zoo attendance are confounded in phases 2–5 and the higher attendance in phase 1 should not be construed as resulting in lower levels of aggression.
The one gorilla, E, who behaved somewhat differently during the study, was the only gorilla in the group who had been nursery-reared subsequent to her 1976 birth. E spent more time in proximity to visitor viewing areas throughout the study and she was the only gorilla to direct affiliative behaviors towards visitors. Her interest in visitors can be attributed to nursery rearing as we know that early care-takers can influence later relationships [
24]. This finding serves to illustrate how the same input variables may have different associations with output variables in members of the same species housed at the same institution as has been reported by both Stoinski et al. and Hashmi and Sullivan [
21,
23].
Despite lack of opportunity to interact with visitors when the zoo was closed, the proportion of time E spent in self-directed activity, a measure of stress, did not vary significantly during the course of the study. Reviewing other studies shows variable results. As mentioned above, Wells did find more self-grooming with higher visitor density [
19]. Carder and Semple explored this relationship in two gorilla groups housed at different institutions and found increased scratching with increased visitor density in one of the groups [
25]. However, in a well-controlled study, Kuhar reports no relationship was found between large crowds and proportion of scans spent self-grooming in two gorilla groups housed at the same institution [
22].
Including multiple input variables in studies is useful in interpreting outcomes and in this study, we found that zoo closure had minimal effects. While a general variable such as the presence or absence of visitors or attendance has the potential to affect the well-being of animals, we need to identify what aspects of visitation are mediating the effects. Is it the behavior of the visitors? Is it exposure to visual and/or auditory and/or olfactory stimuli associated with visitors that influences animals? In the case of the pandemic, could the behavior of animals vary in response to their keepers’ emotions? Such questions should stimulate further research which can be usefully applied to increasing our ability to provide optimal care for the animals we manage.





{kind=link}
{kind=link}
{kind=link}