
Behavioural Impact of Captive Management Changes in Three Species of Testudinidae



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Population
2.3. Husbandry
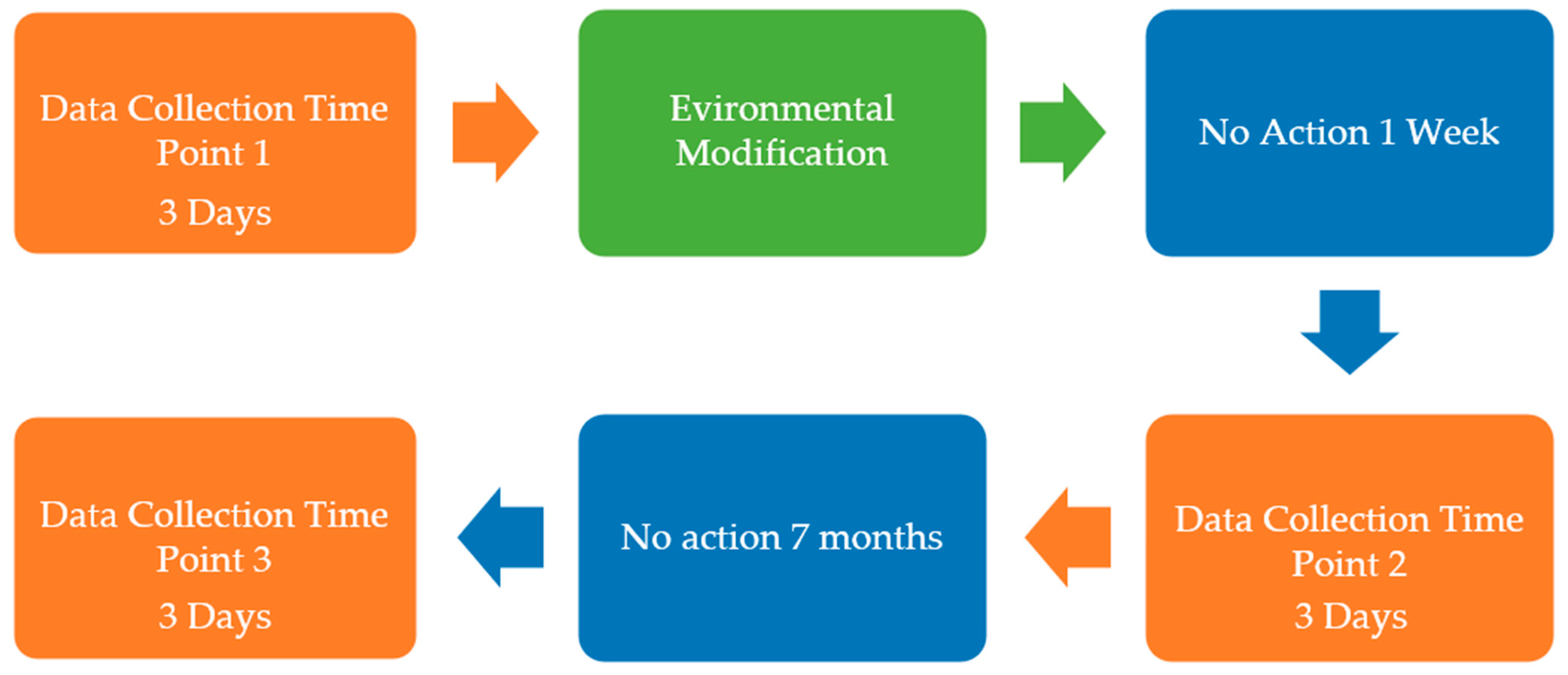
2.4. Study Design
2.5. Behavioural Data Collection
2.6. Data Analysis
3. Results
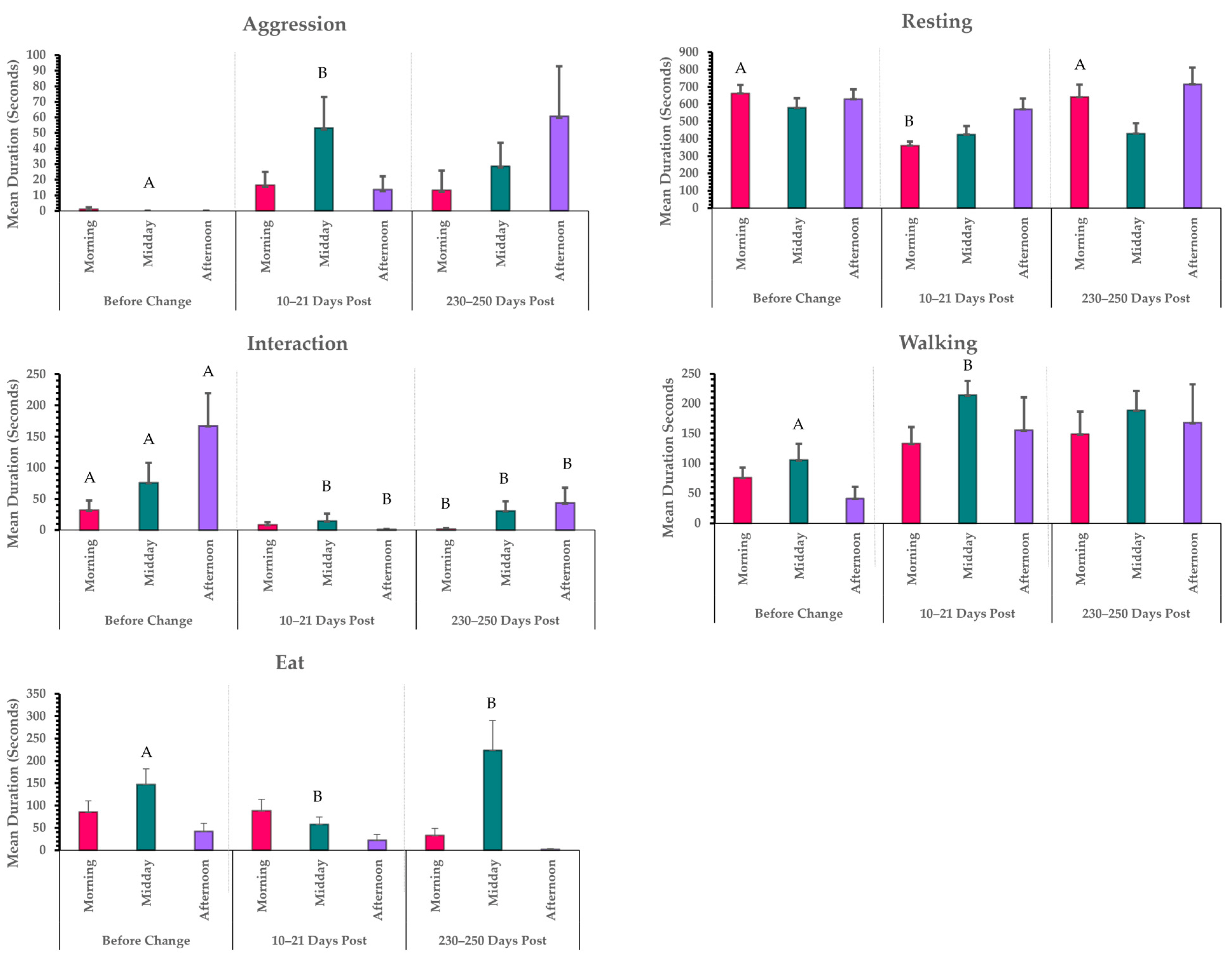
3.1. Activity Budget Analysis Results
3.1.1. Temperature and Time of Day Interactions
3.1.2. Species and Time Point Interactions
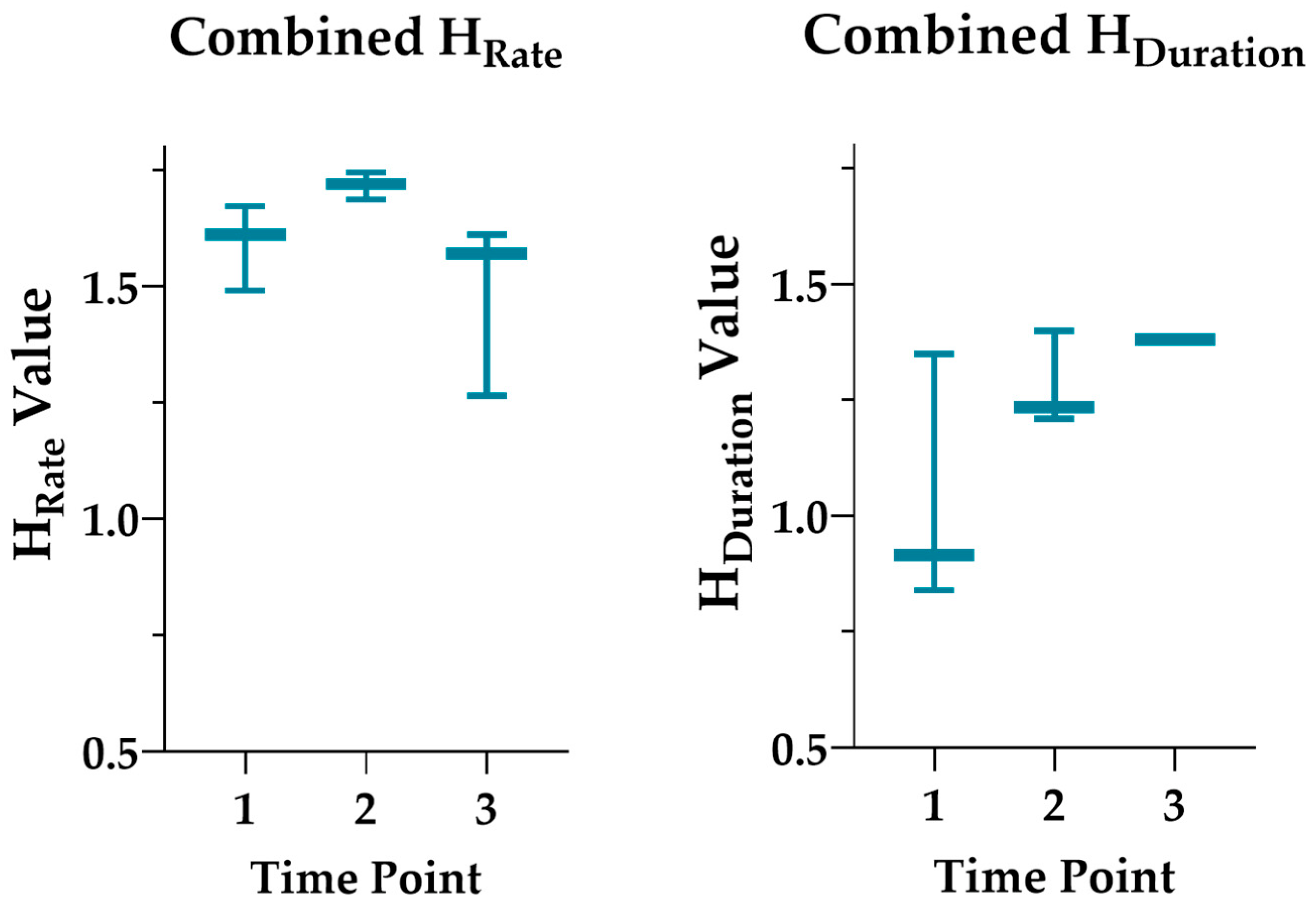
3.2. Diversity Index Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
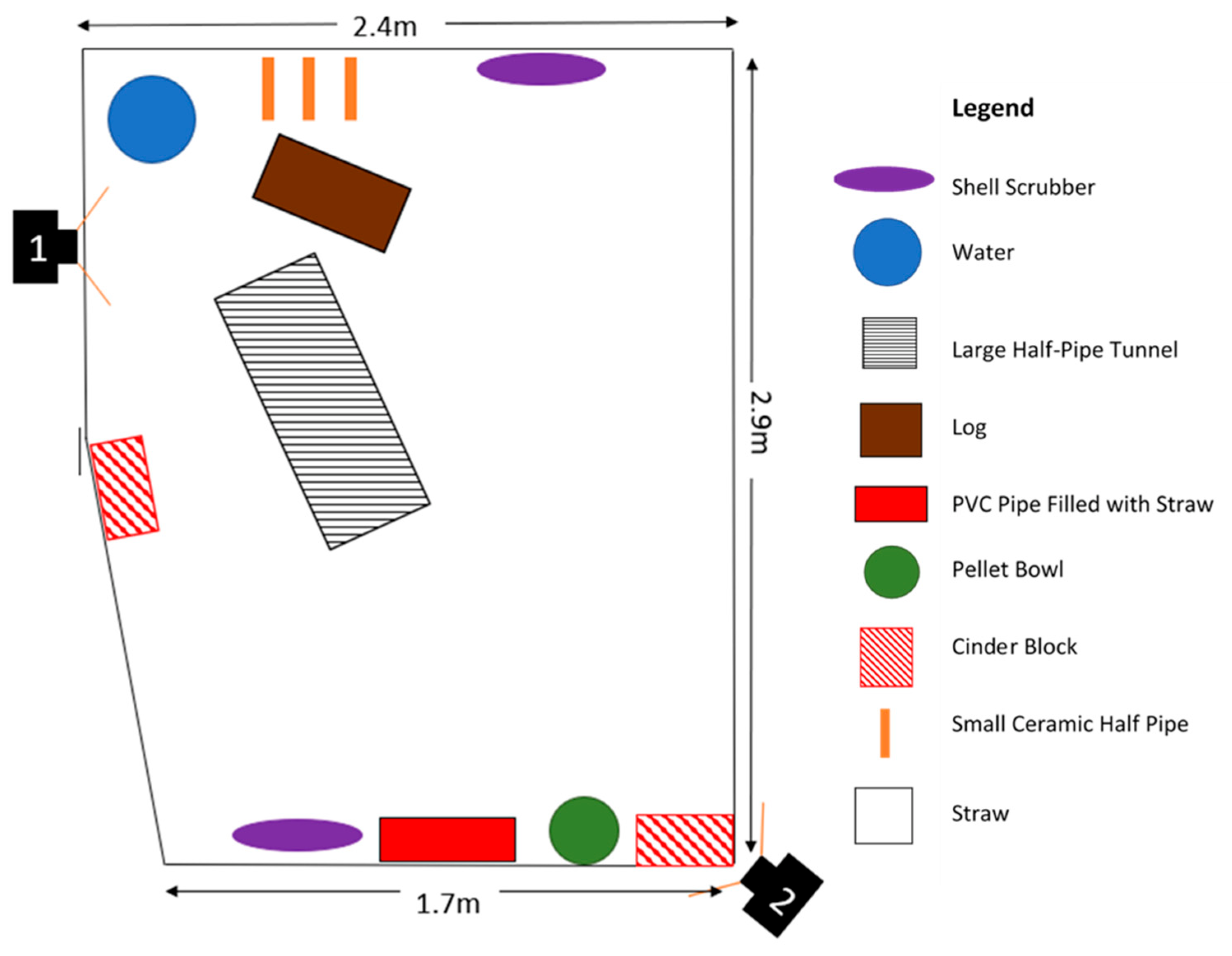
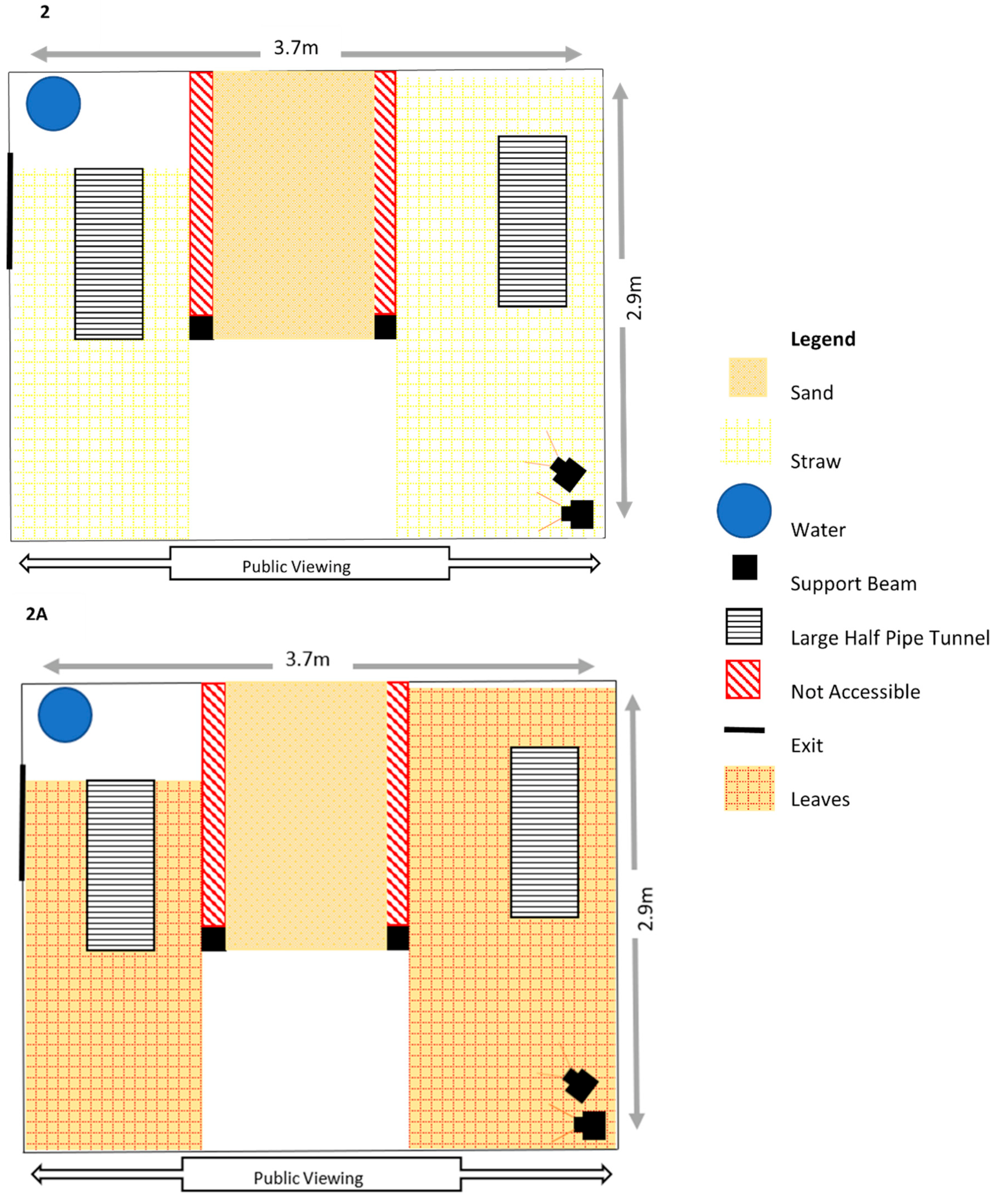
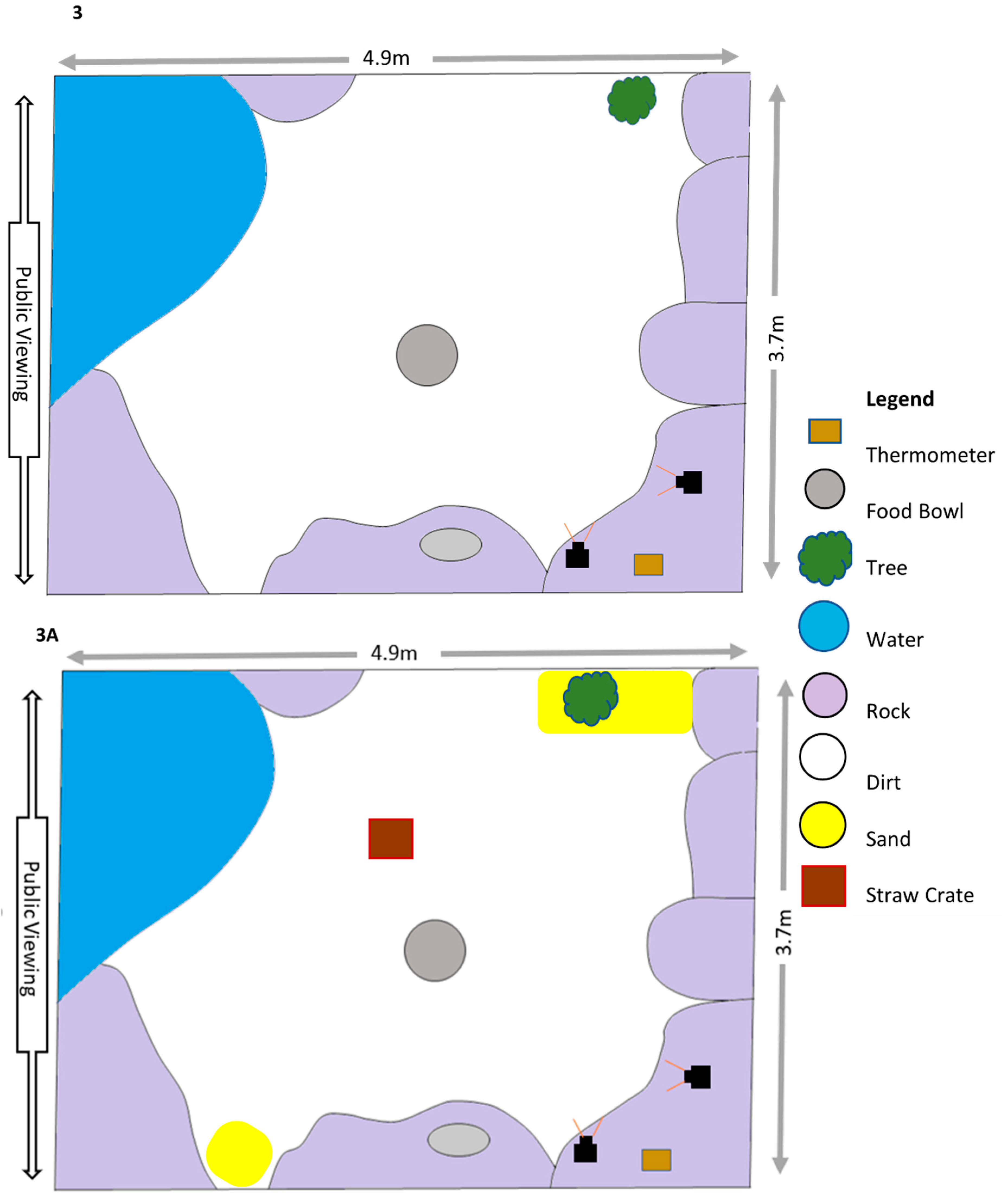
Appendix A. Enclosure Diagrams




References
- Learmonth, M.J.; Sherwen, S.; Hemsworth, P.H. Assessing preferences of two zoo-housed Aldabran giant tortoises (Aldabrachelys gigantea) for three stimuli using a novel preference test. Zoo Biol. 2020, 40, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Gray, J. Zoo Ethics: The Challenges of Compassionate Conservation; CSIRO Publishing: Clayton, Australia, 2017; p. 256. [Google Scholar]
- Sherwen, S.; Hemsworth, L.; Beausoleil, N.; Embury, A.; Mellor, D. An Animal Welfare Risk Assessment Process for Zoos. Animals 2018, 8, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoos South Australia. Animal Welfare Charter. 2018, pp. 1–12. Available online: https://www.zoossa.com.au/wp-content/uploads/2018/08/K004-Animal-Welfare-Charter-2018-1.pdf (accessed on 5 August 2022).
- Sayers, D.W.J. The influence of animal welfare accreditation programmes on zoo visitor perceptions of the welfare of zoo animals. J. Zoo Aquar. Res. 2020, 8, 188–193. [Google Scholar] [CrossRef]
- Cronin, K.A. Working to Supply the Demand: Recent Advances in the Science of Zoo Animal Welfare. J. Zool. Bot. Gard. 2021, 2, 349–350. [Google Scholar] [CrossRef]
- Webster, J. The assessment and implementation of animal welfare: Theory into practice. Rev. Sci. Tech. 2005, 24, 723–734. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.H.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, A.L.; Golder-Dewar, B.; Triggs, J.L.; Sherwen, S.L.; McLelland, D.J. Identification of Animal-Based Welfare Indicators in Captive Reptiles: A Delphi Consultation Survey. Animals 2021, 11, 2010. [Google Scholar] [CrossRef]
- Ahloy-Dallaire, J.; Espinosa, J.; Mason, G. Play and optimal welfare: Does play indicate the presence of positive affective states? Behav. Process. 2018, 156, 3–15. [Google Scholar] [CrossRef]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef]
- Benn, A.; McLelland, D.; Whittaker, A. A Review of Welfare Assessment Methods in Reptiles, and Preliminary Application of the Welfare Quality® Protocol to the Pygmy Blue-Tongue Skink, Tiliqua adelaidensis, Using Animal-Based Measures. Animals 2019, 9, 27. [Google Scholar] [CrossRef]
- Whittaker, A.; Marsh, L. The role of behavioural assessment in determining positive affective states in animals. CAB Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- Broom, D.M. Animal welfare: Concepts and measurement. J. Anim. Sci. 1991, 69, 4167–4175. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human-Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.J.F. Farm Animal Welfare: The Five Freedoms and the Free Market. Vet. J. 2001, 161, 229–237. [Google Scholar] [CrossRef]
- Quality, W. Assessment Protocol for Poultry (Broilers, Laying Hens); Welfare Quality Consortium: Lelystad, The Netherlands, 2009. [Google Scholar]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar]
- Burghardt, G.M. Environmental enrichment and cognitive complexity in reptiles and amphibians: Concepts, review, and implications for captive populations. Appl. Anim. Behav. Sci. 2013, 147, 286–298. [Google Scholar] [CrossRef]
- Warwick, C. Reptilian ethology in captivity: Observations of some problems and an evaluation of their aetiology. Appl. Anim. Behav. Sci. 1990, 26, 1–13. [Google Scholar] [CrossRef]
- Johansson, E. The Impact of Food Enrichment on the Behaviour of Turtles in Captivity. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2017. [Google Scholar]
- Therrien, C.L.; Gaster, L.; Cunningham-Smith, P.; Manire, C.A. Experimental evaluation of environmental enrichment of sea turtles. Zoo Biol. 2007, 26, 407–416. [Google Scholar] [CrossRef]
- Bannister, C.C.; Thomson, A.J.C.; Cuculescu-Santana, M. Can colored object enrichment reduce the escape behavior of captive freshwater turtles? Zoo Biol. 2021, 40, 160–168. [Google Scholar] [CrossRef]
- Bernheim, M.; Livne, S.; Shanas, U. Mediterranean Spur-thighed Tortoises (Testudo graeca) exhibit pre-copulatory behavior particularly under specific experimental setups. J. Ethol. 2020, 38, 355–364. [Google Scholar] [CrossRef]
- Case, B.C. Environmental Enrichment for Captive Eastern Box Turtles (Terrapene carolina carolina). Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2003. [Google Scholar]
- Case, B.C.; Lewbart, G.A.; Doerr, P.D. The physiological and behavioural impacts of and preference for an enriched environment in the eastern box turtle (Terrapene carolina carolina). Appl. Anim. Behav. Sci. 2005, 92, 353–365. [Google Scholar] [CrossRef]
- Cassola, F.M.; Henaut, Y.; Cedeño-Vázquez, J.R.; Méndez-De La Cruz, F.R.; Morales-Vela, B. Temperament and sexual behaviour in the Furrowed Wood Turtle Rhinoclemmys areolata. PLoS ONE 2020, 15, e0244561. [Google Scholar] [CrossRef] [PubMed]
- Fossette, S.; Gaspar, P.; Handrich, Y.; Maho, Y.L.; Georges, J.-Y. Dive and beak movement patterns in leatherback turtles Dermochelys coriacea during internesting intervals in French Guiana. J. Anim. Ecol. 2008, 77, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Freeland, L.; Ellis, C.; Michaels, C.J. Documenting aggression, dominance and the impacts of visitor interaction on Galápagos tortoises (Chelonoidis nigra) in a zoo setting. Animals 2020, 10, 699. [Google Scholar] [CrossRef] [PubMed]
- Passos, L.F.; Mello, H.E.S.; Young, R.J. Enriching Tortoises: Assessing Color Preference. J. Appl. Anim. Welf. Sci. 2014, 17, 274–281. [Google Scholar] [CrossRef]
- Delfour, F.; Vaicekauskaite, R.; García-Párraga, D.; Pilenga, C.; Serres, A.; Brasseur, I.; Pascaud, A.; Perlado-Campos, E.; Sánchez-Contreras, G.J.; Baumgartner, K.; et al. Behavioural Diversity Study in Bottlenose Dolphin (Tursiops truncatus) Groups and Its Implications for Welfare Assessments. Animals 2021, 11, 1715. [Google Scholar] [CrossRef]
- Spain, M.; Fuller, G.; Allard, S. Effects of Habitat Modifications on Behavioral Indicators of Welfare for Madagascar Giant Hognose Snakes (Leioheterodon madagascariensis). Anim. Behav. Cogn. 2020, 7, 70–81. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does enrichment improve reptile welfare? Leopard geckos (Eublepharis macularius) respond to five types of environmental enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar] [CrossRef]
- Hall, K.; Bryant, J.; Staley, M.; Whitham, J.C.; Miller, L.J. Behavioural diversity as a potential welfare indicator for professionally managed chimpanzees (Pan troglodytes): Exploring variations in calculating diversity using species-specific behaviours. Anim. Welf. 2021, 30, 381–392. [Google Scholar] [CrossRef]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef]
- National Health and Medical Research Council (NHMRC). Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; NHMRC: Canberra, Australia, 2013.
- ZooMonitor, version 4.1; Lincoln Park Zoo: Chicago, IL, USA, 2022.
- Yon, L.; Williams, E.; Harvey, N.D.; Asher, L. Development of a behavioural welfare assessment tool for routine use with captive elephants. PLoS ONE 2019, 14, e0210783. [Google Scholar] [CrossRef]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing reptile welfare using behavioural criteria. Practice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- SPSS, 24.0; IBM Corporation: Armok, NY, USA, 2021.
- Henderson, P.A.; Southwood, T.R.E. Ecological Methods, 5th ed.; Wiley: Hoboken, NJ, USA, 2016; pp. 579–580. [Google Scholar]
- Wisconsin, S. Measuring Biodiversity: Google Sheets Tutorial. Available online: https://dnr.wisconsin.gov (accessed on 15 September 2021).
- Rose, F.L.; Judd, F.W. Activity and home range size of the Texas tortoise, Gopherus berlandieri, in south Texas. Herpetologica 1975, 31, 448–456. [Google Scholar]
- Agha, M.; Augustine, B.; Lovich, J.E.; Delaney, D.; Sinervo, B.; Murphy, M.O.; Ennen, J.R.; Briggs, J.R.; Cooper, R.; Price, S.J. Using motion-sensor camera technology to infer seasonal activity and thermal niche of the desert tortoise (Gopherus agassizii). J. Therm. Biol. 2015, 49–50, 119–126. [Google Scholar] [CrossRef] [PubMed]
- McMaster, M.K.; Downs, C.T. Thermoregulation in leopard tortoises in the Nama-Karoo: The importance of behaviour and core body temperatures. J. Therm. Biol. 2013, 38, 178–185. [Google Scholar] [CrossRef]
- Judd, F.W.; Rose, F.L. Aspects of the Thermal Biology of the Texas Tortoise, Gopherus berlandieri (Reptilia, Testudines, Testudinidae). J. Herpetol. 1977, 11, 147. [Google Scholar] [CrossRef]
- Falcón, W.; Hansen, D.M. Island rewilding with giant tortoises in an era of climate change. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terespolsky, A.; Brereton, J.E. Investigating the Thermal Biology and Behaviour of Captive Radiated Tortoises. J. Vet. Med. Anim. Sci. 2021, 4, 1–6. [Google Scholar]
- Wheler, C.L.; Fa, J.E. Enclosure utilization and activity of Round Island geckos (Phelsuma guentheri). Zoo Biol. 1995, 14, 361–369. [Google Scholar] [CrossRef]
- Hoehfurtner, T.; Wilkinson, A.; Walker, M.; Burman, O.H.P. Does enclosure size influence the behaviour & welfare of captive snakes (Pantherophis guttatus)? Appl. Anim. Behav. Sci. 2021, 243, 105435. [Google Scholar] [CrossRef]
- Calvert, S.; Haskell, M.; Wemelsfelder, F.; Lawrence, A.B.; Mendl, M.T. The Effect of Substrate-Enriched and Substrate-Impoverished Housing Environments On the Diversity of Behaviour in Pigs. Behaviour 1996, 133, 741–761. [Google Scholar] [CrossRef] [Green Version]
- Frézard, A.; Pape, G.L. Contribution to the welfare of captive wolves (Canis lupus lupus): A behavioral comparison of six wolf packs. Zoo Biol. 2003, 22, 33–44. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Bryant, J.L.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Behavioral diversity as a potential positive indicator of animal welfare in bottlenose dolphins. PLoS ONE 2021, 16, e0253113. [Google Scholar] [CrossRef] [PubMed]
- Watters, J.V.; Krebs, B.L.; Eschmann, C.L. Assessing Animal Welfare with Behavior: Onward with Caution. J. Zool. Bot. Gard. 2021, 2, 75–87. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour | Description |
|---|---|
| Resting | Includes basking, sleeping, or resting on ground with no weight on limbs, for 3 s or more. No other activities being performed. All instances of resting are included whether awake or asleep. |
| Walking | Two or more steps in one direction. One foot removed from ground at a time. |
| Digging | One or more limbs (front or hind legs) moving substrate. Motion must be repeated twice or more to be counted as digging. |
| Standing still | Tortoise must be bearing its weight on 1 or more legs for 2 s or more |
| Bathing | Includes submerging whole or part of body for more than 3 s |
| Enrichment and object interaction | Touching or playing with any object provided for play and or enrichment. This includes interaction with food dispensers (ball or similar, NOT a stationary food bowl and NOT the act of eating), using or touching tunnels/hides/ramps/logs/etc. If tortoise is inside a tunnel when set observation period starts and they cannot be reasonably seen, this is counted as individual not observed. However, if the tortoise moves into tunnel during the observation period and remains in there, this is counted as use of tunnel. |
| Eating/Drinking | Any eating or drinking activity where food/water is consumed, or food is chewed. |
| Vocalisations | Any audible noises made by the tortoises by nose or throat. This excludes defaecation and/or digestion noises. |
| Co-occupant interaction | A positive or neutral interaction with another tortoise. This includes climbing, leaning, touching, non-aggressive approach, head bobbing, etc. |
| Co-occupant aggression | Any negative aggressive action, or attempted action, towards another occupant including shell ramming, charging, displacement, hooking, aggressive social posturing, scratching, biting. |
| Stereotypies/Abnormal behaviour | Includes pacing and other repetitive behaviour and interaction with transparent boundaries. Behaviour must be completed three times consecutively. |
| Not Observable | For use only when focal animal is not visible. |
| Factor | Behaviour | Df (df1, df2) | F-Value | Significance |
|---|---|---|---|---|
| Species | Aggression | 1, 107 | 8.300 | 0.01 |
| Object Interaction | 1, 107 | 11.704 | 0.01 | |
| Eating/Drinking | 1, 107 | 5.649 | 0.02 | |
| Standing | 1, 107 | 11.136 | 0.01 | |
| Co-occupant interaction | 1, 107 | 18.758 | <0.001 | |
| Abnormal Behaviours | 1, 107 | 5.024 | 0.03 | |
| Time Point | Aggression | 2, 107 | 4.703 | 0.01 |
| Object Interaction | 2, 107 | 4.868 | 0.01 | |
| Resting | 2, 107 | 4.486 | 0.01 | |
| Standing | 2, 107 | 4.655 | 0.01 | |
| Co-occupant interaction | 2, 107 | 20.696 | <0.001 | |
| Walking | 2, 107 | 6.169 | 0.003 | |
| Temperature | Resting | 1, 75 | 8.223 | 0.005 |
| Co-occupant Interaction | 1, 75 | 21.700 | <0.001 | |
| Time of Day | Aggression | 2, 75 | 31.000 | <0.001 |
| Eating/Drinking | 2, 75 | 6.410 | 0.003 | |
| Resting | 2, 75 | 6.808 | 0.002 | |
| Co-occupant Interaction | 2, 75 | 6.522 | 0.002 | |
| Walking | 2, 75 | 4.390 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turner, J.T.; Whittaker, A.L.; McLelland, D. Behavioural Impact of Captive Management Changes in Three Species of Testudinidae. J. Zool. Bot. Gard. 2022, 3, 555-572. https://doi.org/10.3390/jzbg3040041
Turner JT, Whittaker AL, McLelland D. Behavioural Impact of Captive Management Changes in Three Species of Testudinidae. Journal of Zoological and Botanical Gardens. 2022; 3(4):555-572. https://doi.org/10.3390/jzbg3040041
Chicago/Turabian StyleTurner, Jessica T., Alexandra L. Whittaker, and David McLelland. 2022. "Behavioural Impact of Captive Management Changes in Three Species of Testudinidae" Journal of Zoological and Botanical Gardens 3, no. 4: 555-572. https://doi.org/10.3390/jzbg3040041