
Baseline Behavioral Data and Behavioral Correlates of Disturbance for the Lake Oku Clawed Frog (Xenopus longipes)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Ethical Approval
2.3. Ethogram
2.4. Baseline Behavioral Data
Statistical Analysis
2.5. Behavioral Response to Stressor
Statistical Analysis
3. Results
3.1. Ethogram
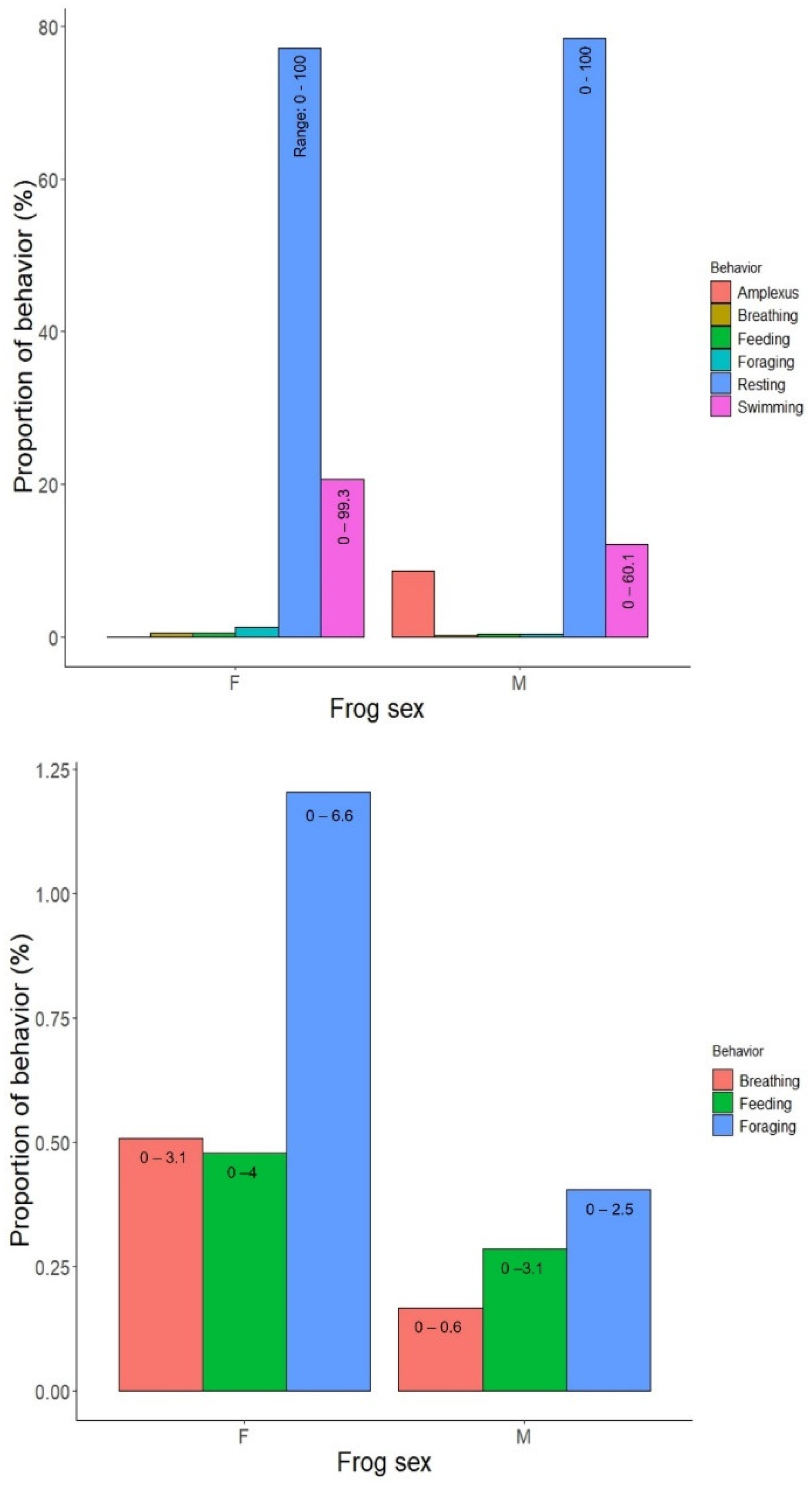
3.2. Baseline Behavioral Data
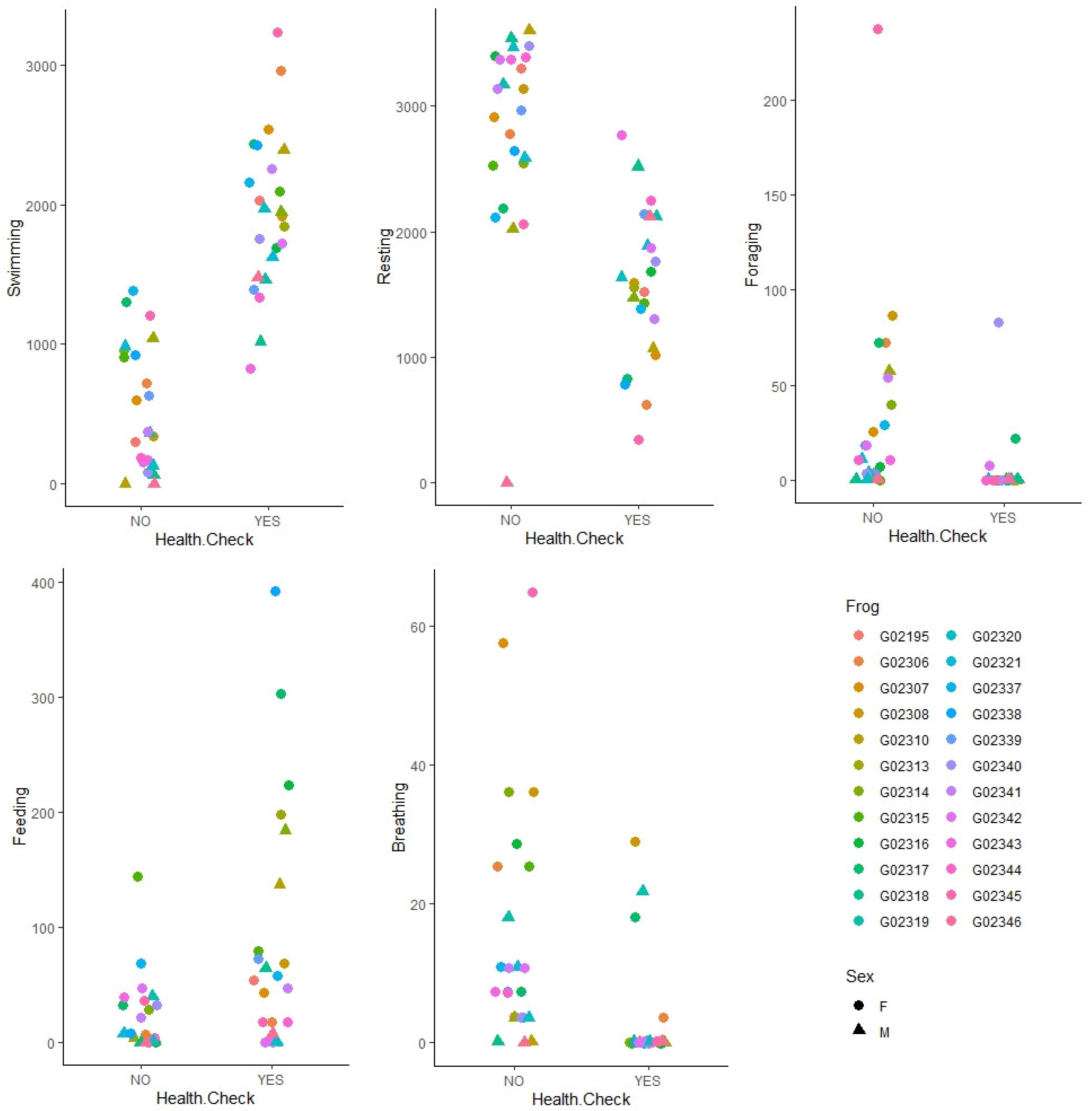
3.3. Behavioral Response to Stressor

{kind=link}
{kind=link}
| Model | Response Variable | Parameter | Estimate (SD for Random Effect) | Standard Error of Estimate | t Value | Lower 95% CI of Estimate | Upper 95% CI of Estimate |
|---|---|---|---|---|---|---|---|
| Behavior = sex + time of day + frog (tank) | Swimming | Intercept | 656.43 | 159.51 | 4.115 | 348.306 | 964.553 |
| Sex (M) | −399.70 | 250.75 | −1.594 | −889.705 | 90.303 | ||
| Time (noon) | 174.00 | 145.98 | 1.192 | −111.794 | 459.794 | ||
| Time (pm) | 365.70 | 145.98 | 2.505 | 79.906 | 651.494 | ||
| R2 Marginal | 0.104 | - | - | - | - | ||
| R2 Conditional | 0.525 | - | - | - | - | ||
| Random effect | 475.9 | - | - | - | - | ||
| Resting | Intercept | 2781.37 | 200.78 | 13.853 | 2392.367 | 3170.374 | |
| Sex (M) | 136.04 | 335.39 | 0.406 | −519.360 | 791.448 | ||
| Time (noon) | −99.15 | 150.05 | −0.661 | −392.906 | 194.606 | ||
| Time (pm) | −180.75 | 150.05 | −1.205 | −474.506 | 113.006 | ||
| R2 Marginal | 0.013 | - | - | - | - | ||
| R2 Conditional | 0.638 | - | - | - | - | ||
| Random effect | 683.9 | - | - | - | - | ||
| Foraging | Intercept | 39.59 | 10.96 | 3.611 | 18.463 | 60.725 | |
| Sex (M) | −26.72 | 13.36 | −2.000 | −52.830 | 0.6149 | ||
| Time (noon) | 3.00 | 14.30 | 0.210 | −24.934 | 30.934 | ||
| Time (pm) | 2.10 | 14.30 | 0.147 | −25.834 | 30.034 | ||
| R2 Marginal | 0.057 | - | - | - | - | ||
| R2 Conditional | 0.082 | - | - | - | - | ||
| Random effect | 8.188 | - | - | - | - | ||
| Feeding | Intercept | 23.609 | 5.718 | 4.129 | 12.592 | 34.625 | |
| Sex (M) | −6.373 | 7.184 | −0.887 | −20.412 | 7.666 | ||
| Time (noon) | −8.400 | 7.275 | −1.155 | −22.642 | 5.842 | ||
| Time (pm) | −12.450 | 7.275 | −1.711 | −26.692 | 1.792 | ||
| R2 Marginal | 0.050 | - | - | - | - | ||
| R2 Conditional | 0.112 | - | - | - | - | ||
| Random effect | 6.649 | - | - | - | - | ||
| Breathing | Intercept | 1.930 × e01 | 4.205 | 4.589 | 11.186 | 27.408 | |
| Sex (M) | −1.165 × e01 | 5.031 | −2.315 | −21.308 | 1.986 | ||
| Time (noon) | −4.174 × e−14 | 5.558 | 0.00 | −10.757 | 10.757 | ||
| Time (pm) | −4.950 | 5.558 | −0.891 | 15.707 | 5.807 | ||
| R2 Marginal | 0.083 | - | - | - | - | ||
| R2 Conditional | 0.088 | - | - | - | - | ||
| Random effect | 1.395 | - | - | - | - | ||
| Behavior = health check status + sex + frog (tank) | Swimming | Intercept | 608.284 | 114.36 | 5.319 | 384.294 | 829.647 |
| Healthcheck (yes) | 1397.250 | 106.70 | 13.095 | 1184.948 | 1609.553 | ||
| Sex (M) | −234.632 | 177.11 | −1.325 | −578.974 | 119.898 | ||
| R2 Marginal | 0.671 | - | - | - | - | ||
| R2 Conditional | 0.820 | - | - | - | - | ||
| Random effect | 336.5 | - | - | - | - | ||
| Resting | Intercept | 608.284 | 160.25 | 17.547 | 384.294 | 829.647 | |
| Health check (yes) | 1397.250 | 183.11 | −6.839 | 1184.948 | 1609.553 | ||
| Sex (M) | −234.632 | 239.44 | 0.132 | −578.974 | 119.898 | ||
| R2 Marginal | 0.671 | - | - | - | - | ||
| R2 Conditional | 0.820 | - | - | - | - | ||
| Random effect | 336.5 | - | - | - | - | ||
| Foraging | Intercept | 37.187 | 8.444 | 4.404 | 20.836 | 53.537 | |
| Health check (yes) | −27.150 | 10.875 | −2.497 | −48.206 | −6.093 | ||
| Sex (M) | −18.469 | 11.962 | −1.544 | −41.632 | 4.694 | ||
| R2 Marginal | 0.155 | - | - | - | - | ||
| R2 Conditional | 0.155 | - | - | - | - | ||
| Random effect | 0.00 | - | - | - | - | ||
| Feeding | Intercept | 30.251 | 17.36 | 1.743 | −3.589 | 63.861 | |
| Health check (yes) | 61.050 | 22.35 | 2.731 | 17.765 | 104.334 | ||
| Sex (M) | −29.148 | 24.59 | −1.185 | −76.762 | 18.467 | ||
| R2 Marginal | 0.159 | - | - | - | - | ||
| R2 Conditional | 0.159 | - | - | - | - | ||
| Random effect | 0.00 | - | - | - | - | ||
| Breathing | Intercept | 18.164 | 3.025 | 6.004 | 12.307 | 24.024 | |
| Health check (yes) | −12.900 | 4.630 | −3.553 | −20.137 | −5.663 | ||
| Sex (M) | −7.762 | 4.439 | −1.749 | −16.541 | 0.826 | ||
| R2 Marginal | 0.235 | - | - | - | - | ||
| R2 Conditional | 0.326 | - | - | - | - | ||
| Random effect | 4.612 | - | - | - | - |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melfi, V.A. There Are Big Gaps in Our Knowledge, and Thus Approach, to Zoo Animal Welfare: A Case for Evidence-Based Zoo Animal Management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Behavior as a tool in the assessment of animal welfare. Zoology 2003, 106, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Using behavior to assess animal welfare. Anim. Welf. 2004, 13, 3–7. [Google Scholar]
- Maple, T.L. Toward a Science of Welfare for Animals in the Zoo. J. Appl. Anim. Welf. Sci. 2007, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Tillbrook, A.J.; Ralph, C.R. Hormones, stress and the welfare of animals. Anim. Prod. Sci. 2017, 58, 408–415. [Google Scholar] [CrossRef]
- Mench, J.A.; Mason, G.J. Chapter 9: Behavior. In Animal Welfare; Appleby, M.C., Hughes, B.O., Eds.; CAB International: Cambridge, MA, USA, 1997; pp. 127–141. [Google Scholar]
- Moberg, G.P.; Mench, J.A. The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CABI Publishing: New York, NY, USA, 2000. [Google Scholar]
- Beebee, T.J.C.; Griffiths, R.A. The amphibian decline crisis: A watershed for conservation biology? Biol. Conserv. 2005, 125, 271–285. [Google Scholar] [CrossRef]
- Home Office. Annual Statistics of Scientific Procedures on Living Animals Great Britain 2018; The Stationery Office: London, UK, 2018; ISBN 978-1-5286-1336-1. [Google Scholar]
- Griffiths, R.A.; Pavajeau, L. Captive Breeding, Reintroduction, and the Conservation of amphibians. Conserv. Biol. 2008, 22, 852–861. [Google Scholar] [CrossRef]
- Browne, R.K.; Wolfram, K.; Garcia, G.; Bagaturov, M.F.; Pereboom, Z.J.J.M. Zoo based amphibian research and Conservation Breeding Programs (CBPs). Amphib. Rept. Conserv. 2011, 5, 1–14. [Google Scholar]
- Zippel, K.; Johnson, K.; Gargliardo, R.; Gibson, R.; McFadden, M.; Browne, R.; Martinez, C.; Townsend, E. The Amphibian Ark: A global community for ex situ conservation of amphibians. Herpetol. Conserv. Biol. 2011, 6, 340–352. [Google Scholar]
- Pavajeau, L.; Zippel, K.C.; Gibson, R.; Johnson, K. Amphibian Ark and the 2008 Year of the Frog Campaign. Int. Zoo Yearb. 2008, 42, 24–29. [Google Scholar] [CrossRef]
- Mohanty, N.P.; Measey, J. The global pet trade in amphibians: Species traits, taxonomic bias, and future directions. Biodiv. Conserv. 2019, 28, 3915–3923. [Google Scholar] [CrossRef]
- Brod, S.; Brookes, L.; Garner, T.W.J. Discussing the future of amphibians in research. Lab. Anim. 2019, 48, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Michaels, C.J.; Gini, B.F.; Preziosi, R.F. The importance of natural history and species-specific approaches in amphibian ex-situ conservation. Herpetol. J. 2014, 24, 135–145. [Google Scholar]
- Harding, G.; Griffiths, R.A.; Pavajeau, L. Developments in amphibian captive breeding and reintroduction programs. Conserv. Biol. 2015, 30, 340–349. [Google Scholar] [CrossRef]
- Tapley, B.; Bradfield, K.S.; Michaels, C.; Bungard, M. Amphibians and conservation breeding programmes: Do all threatened amphibians belong on the ark? Biodiv. Conserv. 2015, 24, 2625–2646. [Google Scholar] [CrossRef]
- Hurme, K.; Gonzalez, K.; Halvorsen, M.; Foster, B.; Moore, D.; Chepko-Sade, B.D. Environmental enrichment for dendrobatid frogs. J. Appl. Anim. Welf. Sci. 2003, 6, 285–299. [Google Scholar] [CrossRef]
- Archard, G.A. Refuge use affects daily activity patterns in female Xenopus laevis. Appl. Anim. Behav. Sci. 2013, 145, 123–128. [Google Scholar] [CrossRef]
- Holmes, A.M.; Emmans, C.J.; Jones, N.; Coleman, R.; Smith, T.E.; Hosie, C.A. Impact of tank background on the welfare of the African clawed frog, Xenopus laevis (Daudin). Appl. Anim. Behav. Sci. 2016, 185, 131–136. [Google Scholar] [CrossRef]
- Passos, L.F.; Garcia, G.; Young, R. Neglecting the call of the wild: Captive frogs like the sound of their own voice. PLoS ONE 2017, 12, e0181931. [Google Scholar] [CrossRef] [Green Version]
- Passos, L.F.; Garcia, G.; Young, R. Do captive golden mantella frogs recognise wild conspecifics calls? Responses to the playback of captive and wild calls. J. Zoo Aquar. Res. 2020, 9, 49–54. [Google Scholar] [CrossRef]
- Boultwood, J.; O’Brien, M.; Rose, P. Bold Frogs or Shy Toads? How Did the COVID-19 Closure of Zoological Organisations Affect Amphibian Activity? Animals 2021, 11, 1982. [Google Scholar] [CrossRef]
- Swaisgood, R.R. Current status and future directions of applied behavioral research for animal welfare and conservation. Appl. Anim. Behav. Sci. 2007, 102, 139–162. [Google Scholar] [CrossRef]
- Browne, R.K.; Blackburn, D.C.; Doherty-Bone, T. Species Profile: Lake Oku Clawed Frog (Xenopus longipes), Amphibian Ark, 2009. Available online: http://www.amphibianark.org/wp-content/uploads/2018/08/Species-Profile-Xenopus-longipes-July-09.pdf (accessed on 25 November 2021).
- IUCN SSC Amphibian Specialist Group. Xenopus longipes (Amended Version of 2017 Assessment). The IUCN Red List of Threatened Species, 2020. Available online: https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T58176A177346697.en (accessed on 16 April 2022). [CrossRef]
- Blackburn, D.C.; Evans, B.J.; Pessier, A.P.; Vredenburg, V.T. An enigmatic mortality event in the only population of the Critically Endangered Cameroonian frog Xenopus longipes. Afr. J. Herpetol. 2010, 59, 111–122. [Google Scholar] [CrossRef]
- Doherty-Bone, T.M.; Ndifon, R.K.; Nyingchia, O.N.; Landrie, F.E.; Yonghabi, F.T.; Duffus, A.L.J.; Price, S.; Perkins, M.; Bielby, J.; Kome, N.B.; et al. Morbidity and mortality of the Critically Endangered Lake Oku clawed frog (Xenopus longipes). Endanger. Species Res. 2013, 21, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Michaels, C.J.; Tapley, B.; Harding, L.; Bryant, Z.; Grant, S.; Gill, I.; Nyingchia, O.; Doherty-Bone, T. Breeding and rearing the Critically Endangered Lake Oku Clawed Frog (Xenopus longipes Loumont and Kobel 1991). Amphib. Rept. Conserv. 2015, 9, 100–110. [Google Scholar]
- Tapley, B.; Michaels, C.; Harding, L.; Bryant, Z.; Gill, I.; Grant, S.; Chaney, N.; Dunker, F.; Freiermuth, B.; Willis, J.; et al. Lake Oku Frog—EAZA Best Practice Guidelines; EAZA: Amsterdam, The Netherlands, 2016; Available online: https://www.eaza.net/assets/Uploads/CCC/2016-Lake-Oku-frog-EAZA-Best-Practice-Guidelines-Approved.pdf (accessed on 25 November 2021).
- Tapley, B.; Michaels, C.J.; Doherty-Bone, T.M. The tadpole of the Lake Oku clawed frog Xenopus longipes (Anura; Pipidae). Zootaxa 2015, 3981, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Michaels, C.J.; Das, S.; Chang, Y.M.; Tapley, B. Modulation of foraging strategy in response to distinct prey items and their scents in the aquatic frog Xenopus longipes (Anura: Pipidae). Herpetol. Bull. 2018, 143, 1–6. [Google Scholar]
- Aevarsson, U.A.E.; Graves, A.; Carter, K.C.; Doherty-Bone, T.M.; Kane, D.; Servini, F.; Tapley, B.; Michaels, C.J. Individual identification of the Lake Oku clawed frog (Xenopus longipes) using a photographic identification technique. Herpetol. Conserv. Biol. 2022, in press. [Google Scholar]
- Holmes, A.M.; Emmans, C.J.; Smith, T.; Hosie, L. Unpublished data. Available from the authors (contact LH).
- Oishi, T.; Nagai, K.; Harada, Y.; Narus, M.; Ohtani, M.; Kawano, E.; Tamotsu, S. Circadian Rhythms in Amphibians and Reptiles: Ecological Implications. Biol. Rhythm Res. 2004, 35, 105–120. [Google Scholar] [CrossRef]
- Doherty-Bone, T.M.; Nyingchia, O.N.; Tapley, B. Cannibalism in the Critically Endangered Lake Oku Clawed Frog: A possible cause of morbidities and mortalities? Herpetol. Notes 2018, 11, 667–669. [Google Scholar]
- Bates, D. Fitting linear mixed models in R. R News 2005, 5, 27–30. [Google Scholar]
- De Boeck, P.; Bakker, M.; Zwister, R.; Nivard, M.; Hofman, A.; Tuerlinckx, F.; Partchev, I. The Estimation of Item Response Models with the lmer Function from the lme4 Package in R. J. Stat. Softw. 2011, 39, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.4.7. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 7 January 2022).
- Goode, K.; Rey, K. ggResidpanel Tutorial and User Manual. 2019. Available online: https://goodekat.github.io/ggResidpanel-tutorial/tutorial.html (accessed on 2 October 2021).
- Rose, P.; Evans, C.; Coffin, R.; Miller, R.; Nash, S. Using student-centred research to evidence-base exhibition of reptiles and amphibians: Three species-specific case studies. J. Zoo Aquar. Res. 2014, 2, 25–32. [Google Scholar] [CrossRef]
- Loumont, C.; Kobel, H.R. Xenopus longipes sp. nov., a new polyploid pipid from western Cameroon. Rev. Suisse Zool. 1991, 98, 731–738. [Google Scholar] [CrossRef]
- Preece, D.J. The captive management and breeding of poison-dart frogs, family Dendrobatidae, at Jersey Wildlife Preservation Trust. Dodo 1998, 34, 103–114. [Google Scholar]
- Lisboa, C.S.; Vaz, R.I.; Brasileiro, C.A. Captive breeding program for Scinax alcatraz (Anura: Hylidae): Introducing amphibian ex situ conservation in Brazil. Amphib. Rept. Conserv. 2021, 15, 279–288. [Google Scholar]
- Michaels, C.J.; Fahrbach, M.; Harding, L.; Bryant, Z.; Capon-Doyle, J.S.; Grant, S.; Gill, I.; Tapley, B. Relating natural climate and phenology to captive husbandry in two midwife toads (Alytes obstetricans and A. cisternasii) from different climatic zones. Alytes 2016, 33, 2–11. [Google Scholar]
- Hosie, C.A.; Smith, T.E. Behavioral Biology of Amphibians. In Behavioral Biology of Laboratory Animals; Coleman, K., Schaprio, S.J., Eds.; Routledge: Oxfordshire, UK, 2021; pp. 345–359. [Google Scholar]
- Holmes, A.M.; Emmans, C.J.; Coleman, R.; Smith, T.E.; Hosie, C.A. Effects of transportation, transport medium and re-housing on Xenopus laevis (Daudin). Gen. Comp. Endocrinol. 2018, 266, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Emmans, C. Using Behaviour and Glucocorticoids as Non-Invasive Measures of Stress and Welfare in Captive Ambystoma andersoni, a Critically Endangered Species of Salamander Found in Lake Zacapu, Mexico. Master’s Thesis, University of Chester, Chester, UK, 2015. [Google Scholar]
- Cooke, G.M. Use of a translucent refuge for Xenopus tropicalis with the aim of improving welfare. Lab. Anim. 2018, 52, 304–307. [Google Scholar] [CrossRef]


| Behavior | Definition |
|---|---|
| Swimming | Subject is moving from one location to another through the water, exercising front limbs, back limbs or both to travel. This may be horizontally or vertically. |
| Resting | Subject is stationary. None of the subject’s limbs are being exercised to actively travel in any direction. This may be in the water or resting on a substrate. |
| Foraging | Subject is actively searching for food through a substrate using the forelimbs. This may be followed by feeding, for which a separate event is recorded. |
| Feeding | Subject is consuming a food item, rapidly wafting the item towards the face and mouth with forelimbs and often tilting body forwards following. |
| Breathing | Subject is breathing at the surface of the water with the nares breaching the surface. |
| Sloughing | Subject forces out limbs in order to removed shed skin. Swimming will likely become rapid and uncontrolled. The slough is often consumed. |
| Amplex | A male frog grips a female around her inguinal region as part of reproductive behaviour. |
| Behavior | Effect of Sex | Effect of Session |
|---|---|---|
| Swimming | F1,22 = 2.54, p = 0.13 | F2,46 = 3.14, p = 0.053 |
| Resting | F1,22 = 0.16, p = 0.69 | F2,46 = 0.73, p = 0.49 |
| Foraging | F1,22 = 4.0, p = 0.06 | F2,46 = 0.02, p = 0.98 |
| Feeding | F1,22 = 0.79, p = 0.38 | F2,46 = 1.5, p = 0.2286 |
| Breathing | F1,22.003 = 5.36, p = 0.03 | F2,46.003 = 0.53, p = 0.59 |
| Behavior | Effect of Health Check | Effect of Sex |
|---|---|---|
| Swimming | F1,22.58 = 171.5, p < 0.001 | F1,34.3 = 1.755, p = 0.19 |
| Resting | F1,22.58 = 171.5, p < 0.001 | F1,34.3 = 1.756, p = 0.19 |
| Foraging | F1,45 = 6.2, p = 0.016 | F1,45 = 2.38, p = 0.13 |
| Feeding | F1,45 = 7.46, p = 0.009 | F1,45 = 1.40, p = 0.24 |
| Breathing | F1,22.9 = 12.62, p = 0.002 | F1,28.3 = 3.06 p = 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, J.E.; Ellis, C.; Smith, T.E.; Hosie, C.A.; Tapley, B.; Michaels, C.J. Baseline Behavioral Data and Behavioral Correlates of Disturbance for the Lake Oku Clawed Frog (Xenopus longipes). J. Zool. Bot. Gard. 2022, 3, 184-197. https://doi.org/10.3390/jzbg3020016
Dias JE, Ellis C, Smith TE, Hosie CA, Tapley B, Michaels CJ. Baseline Behavioral Data and Behavioral Correlates of Disturbance for the Lake Oku Clawed Frog (Xenopus longipes). Journal of Zoological and Botanical Gardens. 2022; 3(2):184-197. https://doi.org/10.3390/jzbg3020016
Chicago/Turabian StyleDias, Jemma E., Charlotte Ellis, Tessa E. Smith, Charlotte A. Hosie, Benjamin Tapley, and Christopher J. Michaels. 2022. "Baseline Behavioral Data and Behavioral Correlates of Disturbance for the Lake Oku Clawed Frog (Xenopus longipes)" Journal of Zoological and Botanical Gardens 3, no. 2: 184-197. https://doi.org/10.3390/jzbg3020016