
Social Behavior Deficiencies in Captive American Alligators (Alligator mississippiensis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Wild Observation Site and Congregation
2.2. Captive Observation Site and Congregation
2.3. Behavioral and Environmental Data Collection
2.4. Data Analysis
3. Results
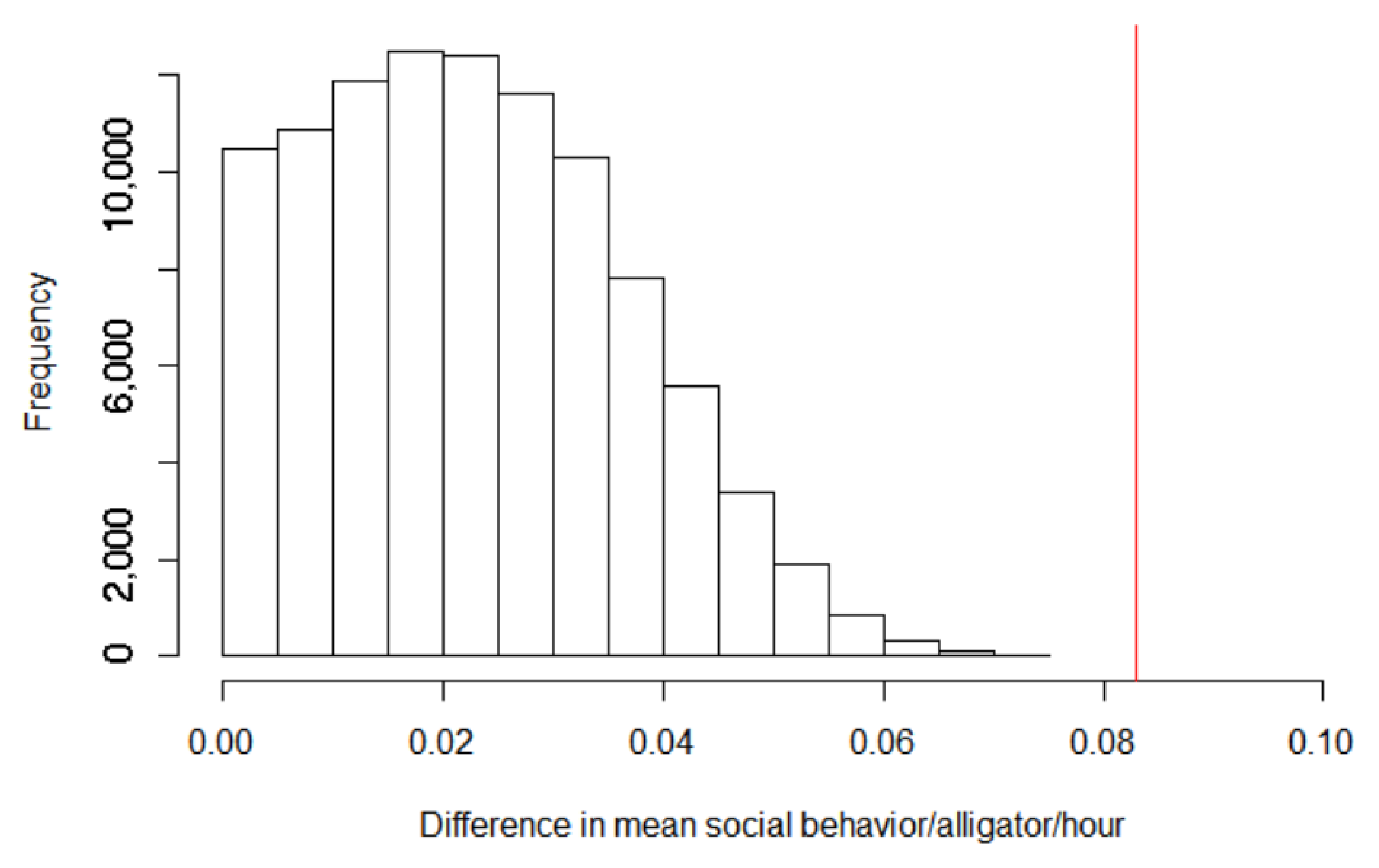
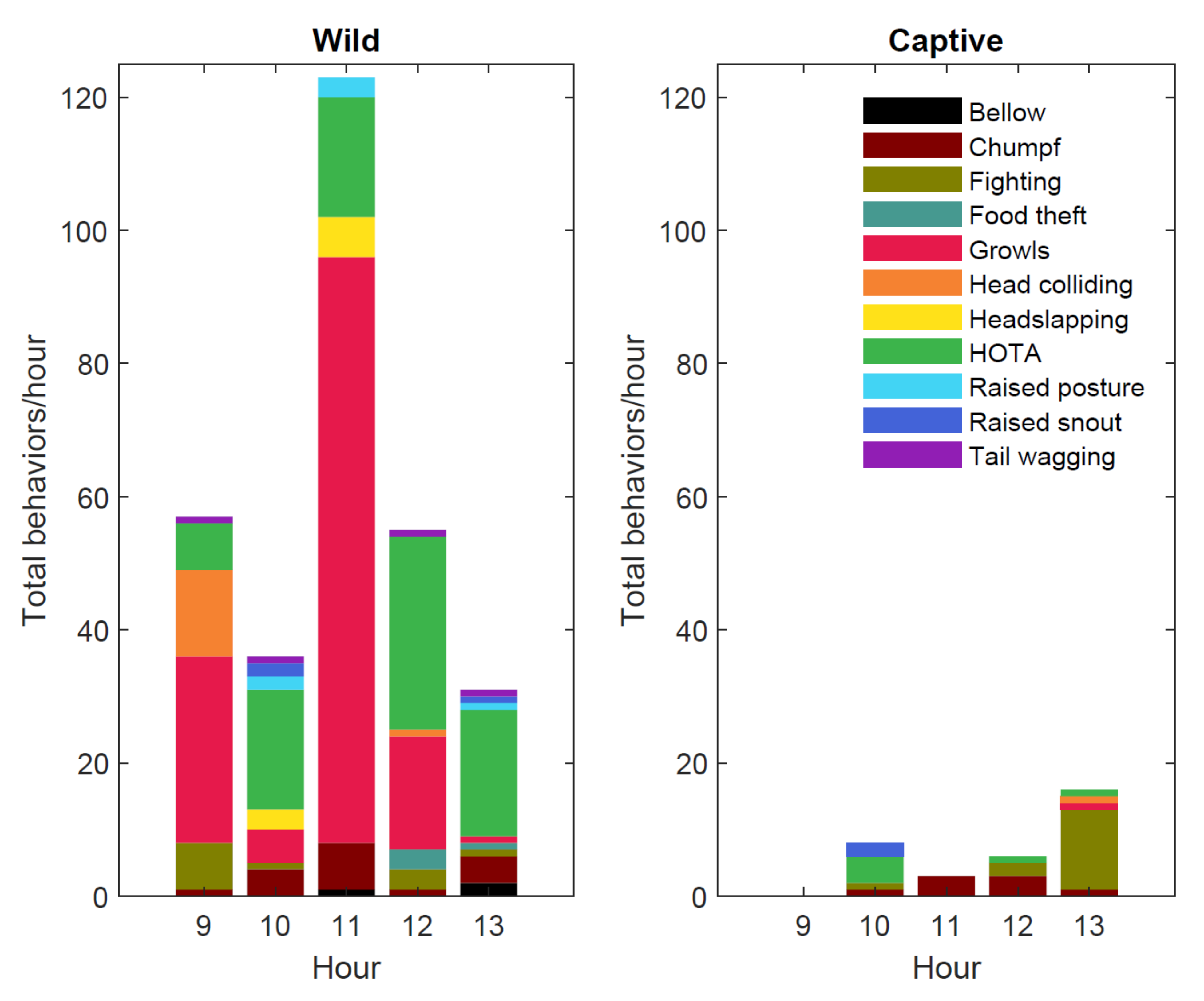
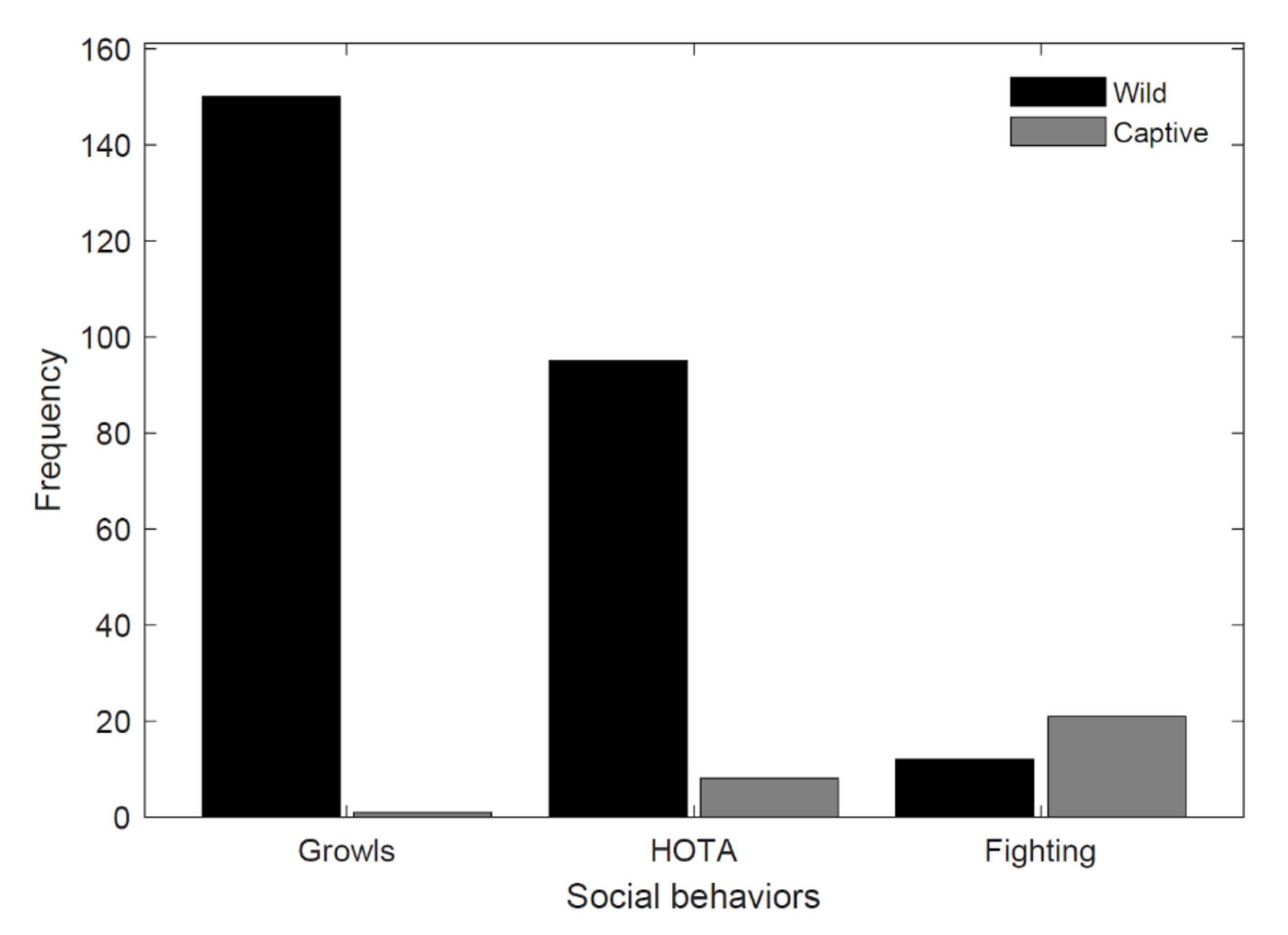
3.1. Social Behaviors
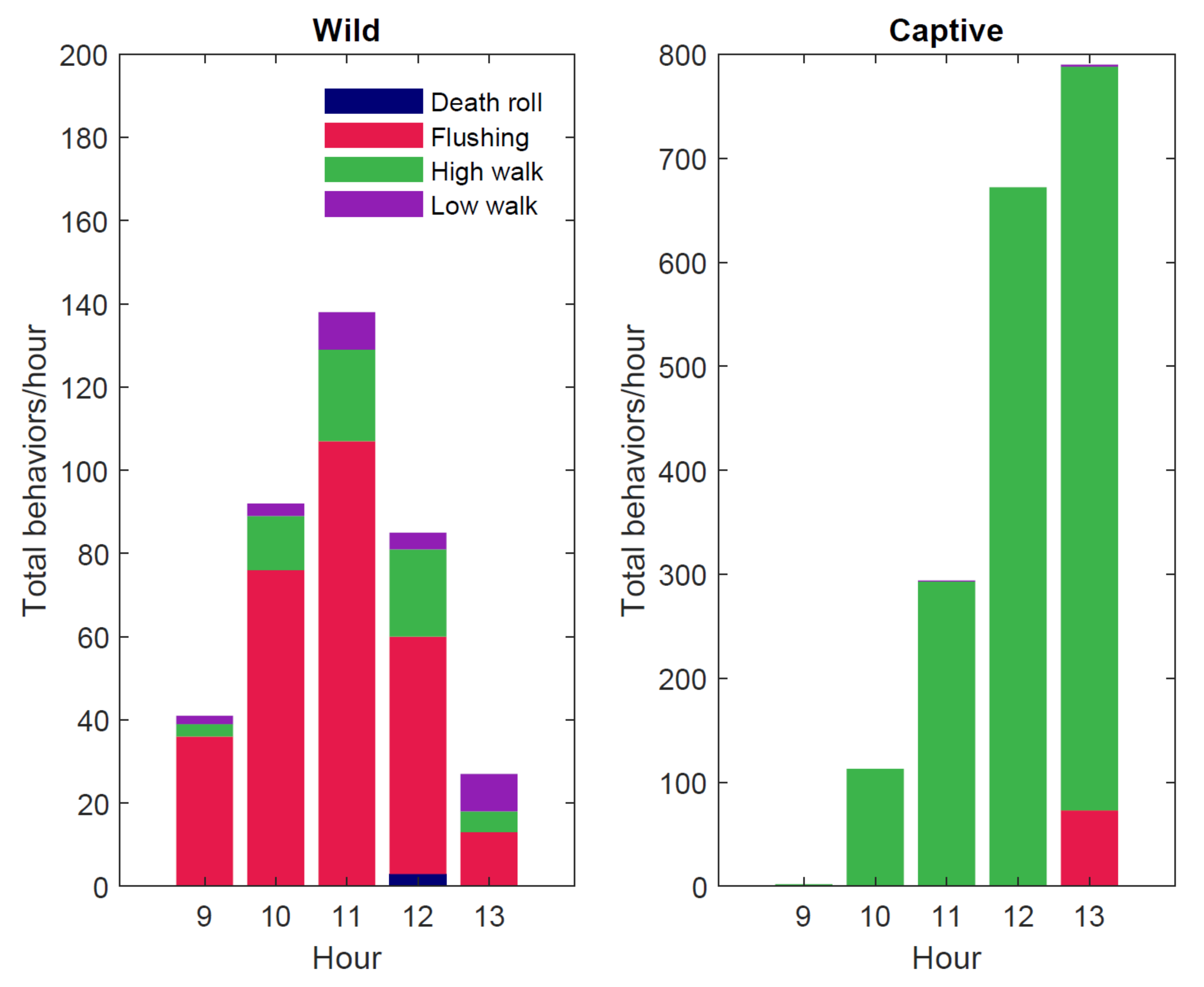
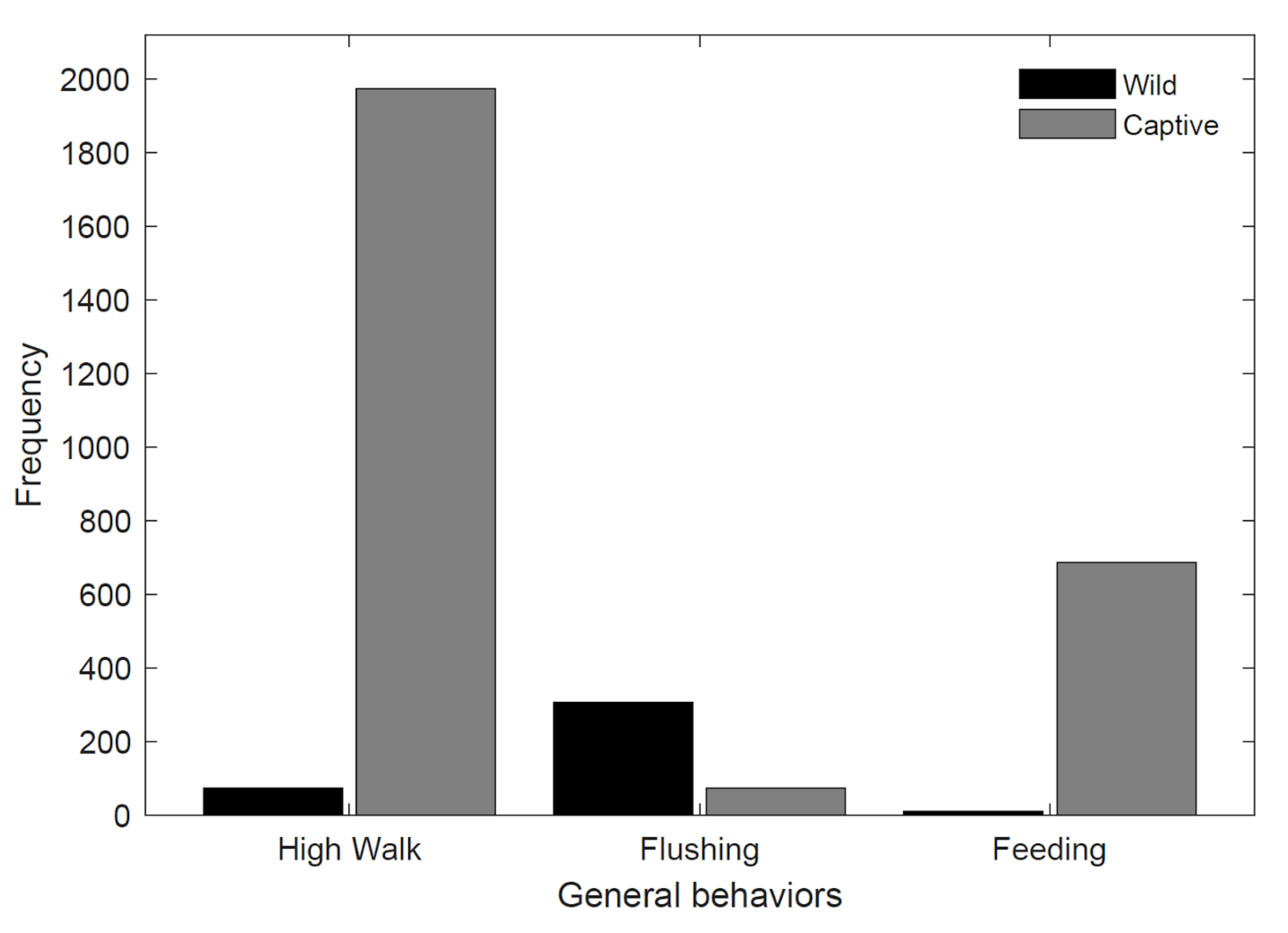
3.2. General Behaviors
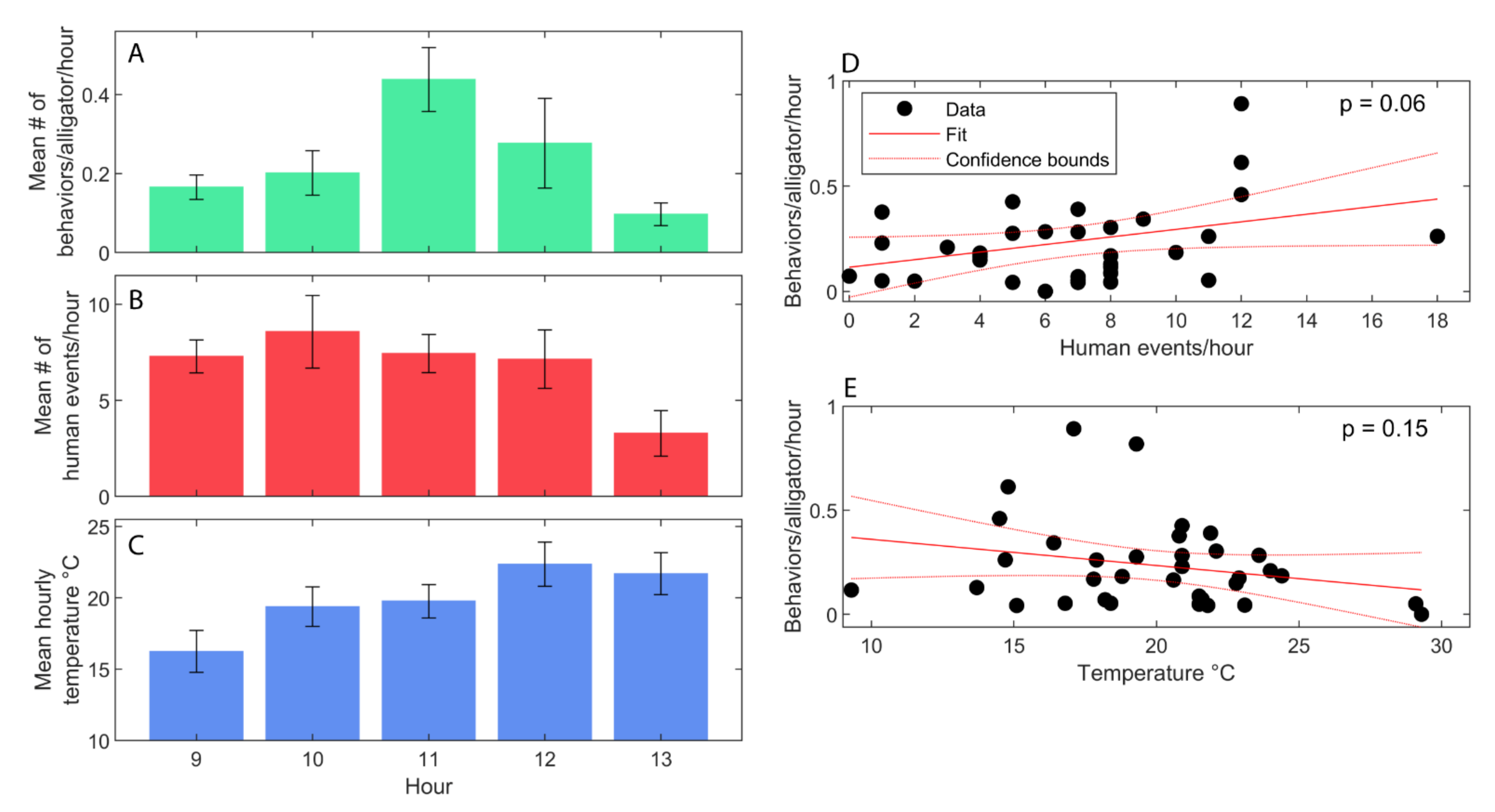
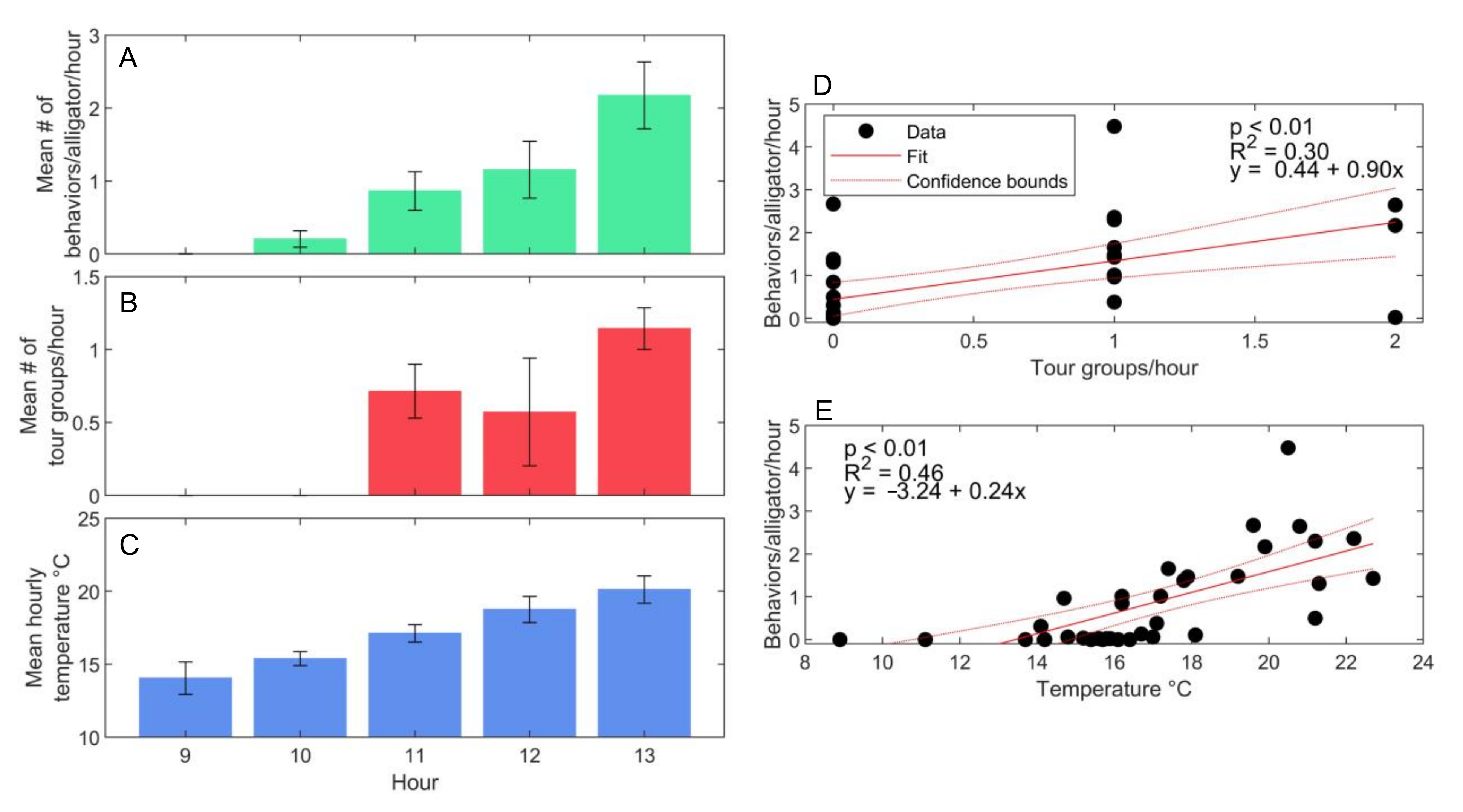
3.3. Environmental and Human Impacts on Behavior
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powell, R.; Conant, R.; Collins, J.T. Peterson Field Guide to Reptiles and Amphibians of Eastern and Central North America, 4th ed.; Houghton Mifflin Harcourt: Boston, MA, USA, 2016; ISBN 978-0-544-66249-0. [Google Scholar]
- Mazzotti, F.J.; Best, G.R.; Brandt, L.A.; Cherkiss, M.S.; Jeffery, B.M.; Rice, K.G. Alligators and Crocodiles as Indicators for Restoration of Everglades Ecosystems. Ecol. Indic. 2009, 9, S137–S149. [Google Scholar] [CrossRef]
- Elsey, R.; Woodward, A.; Balaguera-Reina, S.A. Alligator Mississippiensis. In The IUCN Red List of Threatened Species; IUCN Global Species programme, IUCN SSC, and IUCN Red List Partnership: Gland, Switzerland, 2019. [Google Scholar]
- American Alligator. Available online: http://myfwc.com/wildlifehabitats/profiles/reptiles/alligator/ (accessed on 9 March 2021).
- American Crocodile. Available online: http://myfwc.com/wildlifehabitats/profiles/reptiles/american-crocodile/ (accessed on 22 March 2021).
- Joanen, T.; Life, L.W.; Grand Chenier, L. Propagation of the American Alligator in Captivity. J. Wildl. Manag. 1971, 30, 50–56. [Google Scholar]
- Veasey, J.S.; Waran, N.K.; Young, R.J. On Comparing the Behaviour of Zoo Housed Animals with Wild Conspecifics as a Welfare Indicator. Anim. Welf.-Potters Bar 1996, 5, 13–24. [Google Scholar]
- Inoue, N.; Shimada, M. Comparisons of Activity Budgets, Interactions, and Social Structures in Captive and Wild Chimpanzees (Pan troglodytes). Animals 2020, 10, 1063. [Google Scholar] [CrossRef] [PubMed]
- Glatston, A.R.; Geilvoet-Soeteman, E.; Hora-Pecek, E.; Van Hooff, J.A.R.A.M. The Influence of the Zoo Environment on Social Behavior of Groups of Cotton-Topped Tamarins, Saguinus oedipus oedipus. Zoo Biol. 1984, 3, 241–253. [Google Scholar] [CrossRef]
- Vliet, K.A. Social Displays of the American Alligator (Alligator mississippiensis). Integr. Comp. Biol. 1989, 29, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Garrick, L.D.; Lang, J.W. Social Signals and Behaviors of Adult Alligators and Crocodiles. Integr. Comp. Biol. 1977, 17, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Culter, J.K.; Bowen, C.; Ryan, J.; Perry, J.; Janneman, R.; Lin, W. Exploration of Deep Hole, Myakka River State Park, Florida. In Proceedings of the Joint International Scientific Diving Symposium, Dauphin Island, AL, USA, 24–27 October 2013; pp. 49–60. [Google Scholar]
- Blumstein, D.T.; Anthony, L.L.; Harcourt, R.; Ross, G. Testing a Key Assumption of Wildlife Buffer Zones: Is Flight Initiation Distance a Species-Specific Trait? Biol. Conserv. 2003, 110, 97–100. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fay, M.P.; Shaw, P.A. Exact and Asymptotic Weighted Logrank Tests for Interval Censored Data: The Interval R Package. J. Stat. Softw. 2010, 36, i02. [Google Scholar] [CrossRef] [Green Version]
- MATLAB, Version 9.10 (R2021a); The MathWorks Inc.: Natick, MA, USA, 2021.
- Vaz, J.; Narayan, E.J.; Dileep Kumar, R.; Thenmozhi, K.; Thiyagesan, K.; Baskaran, N. Prevalence and Determinants of Stereotypic Behaviours and Physiological Stress among Tigers and Leopards in Indian Zoos. PLoS ONE 2017, 12, e0174711. [Google Scholar] [CrossRef] [Green Version]
- Emshwiller, M.G.; Gleeson, T.T. Temperature Effects on Aerobic Metabolism and Terrestrial Locomotion in American Alligators. J. Herpetol. 1997, 31, 142–147. [Google Scholar] [CrossRef]
- Elsey, R.M.; Joanen, T.; McNease, L.; Lance, V. Stress and Plasma Corticosterone Levels in the American Alligator—Relationships with Stocking Density and Nesting Success. Comp. Biochem. Physiol. Part A Physiol. 1990, 95, 55–63. [Google Scholar] [CrossRef]
- Fujisaki, I.; Hart, K.M.; Mazzotti, F.J.; Cherkiss, M.S.; Sartain, A.R.; Jeffery, B.M.; Beauchamp, J.S.; Denton, M. Home Range and Movements of American Alligators (Alligator mississippiensis) in an Estuary Habitat. Anim. Biotelemetry 2014, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.; Gochefeld, M.; Murray, B.G., Jr. Role of a Predator’s Eye Size in Risk Perception by Basking Black Iguana, Ctenosaura similis. Anim. Behav. 1991, 42, 471–476. [Google Scholar] [CrossRef]
- Martín, J. When Hiding from Predators Is Costly: Optimization of Refuge Use in Lizards. Etologia 2001, 9, 9–13. [Google Scholar]
- Bolles, R.C. Species-Specific Defense Reactions and Avoidance Learning. Psychol. Rev. 1970, 77, 32. [Google Scholar] [CrossRef]
- Rial, R.V.; Akaârir, M.; Gamundí, A.; Nicolau, C.; Garau, C.; Aparicio, S.; Tejada, S.; Gené, L.; González, J.; De Vera, L.M. Evolution of Wakefulness, Sleep and Hibernation: From Reptiles to Mammals. Neurosci. Biobehav. Rev. 2010, 34, 1144–1160. [Google Scholar] [CrossRef]
- Smith, E.N. Behavioral and Physiological Thermoregulation of Crocodilians. Am. Zool. 1979, 19, 239–247. [Google Scholar] [CrossRef]
- Boucher, M.; Tellez, M.; Anderson, J.T. Activity Budget and Behavioral Patterns of American Crocodiles (Crocodylus acutus) in Belize. Herpetol. Conserv. Biol. 2021, 16, 86–94. [Google Scholar]
- Howell, C.P.; Cheyne, S.M. Complexities of Using Wild versus Captive Activity Budget Comparisons for Assessing Captive Primate Welfare. J. Appl. Anim. Welf. Sci. 2019, 22, 78–96. [Google Scholar] [CrossRef]
- Mendl, M. Assessing the Welfare State. Nature 2001, 410, 31–32. [Google Scholar] [CrossRef] [PubMed]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing Reptile Welfare Using Behavioural Criteria. Practice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Honess, P.E.; Marin, C.M. Behavioural and Physiological Aspects of Stress and Aggression in Nonhuman Primates. Neurosci. Biobehav. Rev. 2006, 30, 390–412. [Google Scholar] [CrossRef] [PubMed]
- Alligood, C.; Leighty, K. Putting the “E” in SPIDER: Evolving Trends in the Evaluation of Environmental Enrichment Efficacy in Zoological Settings. Anim. Behav. Cogn. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Cipreste, C.F.; Young, R.J. Environmental Enrichment: A GAP Analysis. Appl. Anim. Behav. Sci. 2007, 102, 329–343. [Google Scholar] [CrossRef]
- Eagan, T. Evaluation of Enrichment for Reptiles in Zoos. J. Appl. Anim. Welf. Sci. 2019, 22, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Manrod, J.D.; Hartdegen, R.; Burghardt, G.M. Rapid Solving of a Problem Apparatus by Juvenile Black-Throated Monitor Lizards (Varanus albigularis albigularis). Anim. Cogn. 2008, 11, 267–273. [Google Scholar] [CrossRef]
- Rose, P.; Evans, C.; Coffin, R.; Miller, R.; Nash, S. Using Student-Centred Research to Evidence-Base Exhibition of Reptiles and Amphibians: Three Species-Specific Case Studies. J. Zoo Aquar. Res. 2014, 2, 25–32. [Google Scholar] [CrossRef]
- Warwick, C. Important Ethological and Other Considerations of the Study and Maintenance of Reptiles in Captivity. Appl. Anim. Behav. Sci. 1990, 27, 363–366. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Foraging Devices as Enrichment in Captive Walruses (Odobenus rosmarus). Behav. Processes 2019, 168, 103943. [Google Scholar] [CrossRef]
- Fernandez, E.J. Appetitive Search Behaviors and Stereotypies in Polar Bears (Ursus maritimus). Behav. Processes 2021, 182, 104299. [Google Scholar] [CrossRef] [PubMed]
- Kanghae, H.; Thongprajukaew, K.; Inphrom, S.; Malawa, S.; Sandos, P.; Sotong, P.; Boonsuk, K. Enrichment Devices for Green Turtles (Chelonia mydas) Reared in Captivity Programs. Zoo Biol. 2021, 40, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.; Weary, D.M.; Pajor, E.A.; Milligan, B.N. A Scientific Conception of Animal Welfare That Reflects Ethical Concerns. Ethics Anim. 1997, 6, 187–205. [Google Scholar]
- Eversol, C.B.; Henke, S.E.; Ogdee, J.L.; Wester, D.B.; Cooper, A. Nuisance American Alligators: An Investigation into Trends and Public Opinion. Hum.–Wildl. Interact. 2014, 8, 2. [Google Scholar] [CrossRef]
- Janes, D. A Review of Nuisance Alligator Management in the Southeastern United States. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation, Tucson, AZ, USA, 1–5 May 1999; University of Arizona: Tucson, AZ, USA, 1999; pp. 182–185. [Google Scholar]
- Clippinger, T.L.; Bennett, R.A.; Johnson, C.M.; Vliet, K.A.; Deem, S.L.; Orós, J.; Jacobson, E.R.; Schumacher, I.M.; Brown, D.R.; Brown, M.B. Morbidity and Mortality Associated with a New Mycoplasma Species from Captive American Alligators (Alligator mississippiensis). J. Zoo Wildl. Med. 2000, 31, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Pearson, E.L.; Lowry, R.; Dorrian, J.; Litchfield, C.A. Evaluating the Conservation Impact of an Innovative Zoo-Based Educational Campaign: ‘Don’t Palm Us Off’ for Orangutan Conservation. Zoo Biol. 2014, 33, 184–196. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.; Corkery, I.; McKeown, S.; McSweeney, L.; Flannery, K.; Kennedy, D.; O’Riordan, R. An Educational Intervention Maximizes Children’s Learning during a Zoo or Aquarium Visit. J. Environ. Educ. 2020, 51, 361–380. [Google Scholar] [CrossRef]
- Collins, C.; Corkery, I.; McKeown, S.; McSweeney, L.; Flannery, K.; Kennedy, D.; O’Riordan, R. Quantifying the Long-Term Impact of Zoological Education: A Study of Learning in a Zoo and an Aquarium. Environ. Educ. Res. 2020, 26, 1008–1026. [Google Scholar] [CrossRef]
- Kleespies, M.W.; Montes, N.Á.; Bambach, A.M.; Gricar, E.; Wenzel, V.; Dierkes, P.W. Identifying Factors Influencing Attitudes towards Species Conservation—A Transnational Study in the Context of Zoos. Environ. Educ. Res. 2021, 27, 1421–1439. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior Performed | Wild Site | Captive Site |
|---|---|---|
| Bellow | 0.94% (n = 3) | 0.00% (n = 0) |
| Chumpf | 5.31% (n = 17) | 21.43% (n = 9) |
| Fighting | 3.13% (n = 10) | 50.00% (n = 21) |
| Food theft | 1.25% (n = 4) | 0.00% (n = 0) |
| Growl | 46.56% (n = 149) | 2.38% (n = 1) |
| Head colliding | 3.75% (n = 12) | 2.38% (n = 1) |
| Headslapping | 2.81% (n = 9) | 0.00% (n = 0) |
| HOTA | 29.69% (n = 95) | 19.05% (n = 8) |
| Raised posture | 1.88% (n = 6) | 0.00% (n = 0) |
| Raised snout | 3.44 % (n = 11) | 4.76% (n = 2) |
| Roar | 0.00% (n = 0) | 0.00% (n = 0) |
| Tail wagging | 1.25% (n = 4) | 0.00% (n = 0) |
| Water dance | 0.00% (n = 0) | 0.00% (n = 0) |
| Behavior Performed | Wild Site | Captive Site |
|---|---|---|
| Deathroll | 0.71% (n = 3) | 0.00% (n = 0) |
| Feeding | 2.61% (n = 11) | 25.07% (n = 686) |
| Flushing | 72.27% (n = 305) | 2.70% (n = 74) |
| High walk | 18.01% (n = 76) | 72.11% (n = 1973) |
| Low walk | 6.40% (n = 27) | 0.11% (n = 3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walsh, Z.C.; Olson, H.; Clendening, M.; Rycyk, A. Social Behavior Deficiencies in Captive American Alligators (Alligator mississippiensis). J. Zool. Bot. Gard. 2022, 3, 131-146. https://doi.org/10.3390/jzbg3010011
Walsh ZC, Olson H, Clendening M, Rycyk A. Social Behavior Deficiencies in Captive American Alligators (Alligator mississippiensis). Journal of Zoological and Botanical Gardens. 2022; 3(1):131-146. https://doi.org/10.3390/jzbg3010011
Chicago/Turabian StyleWalsh, Zane Cullinane, Hannah Olson, Miranda Clendening, and Athena Rycyk. 2022. "Social Behavior Deficiencies in Captive American Alligators (Alligator mississippiensis)" Journal of Zoological and Botanical Gardens 3, no. 1: 131-146. https://doi.org/10.3390/jzbg3010011