
Is There a Role of Inferior Frontal Cortex in Motor Timing? A Study of Paced Finger Tapping in Patients with Non-Fluent Aphasia

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Task Procedure
2.3. Statistical Analysis
3. Results
3.1. Descriptive Analysis of Finger Tapping Data
3.2. One-Way ANOVA
3.3. Repeated Measures MANOVA
- (a)
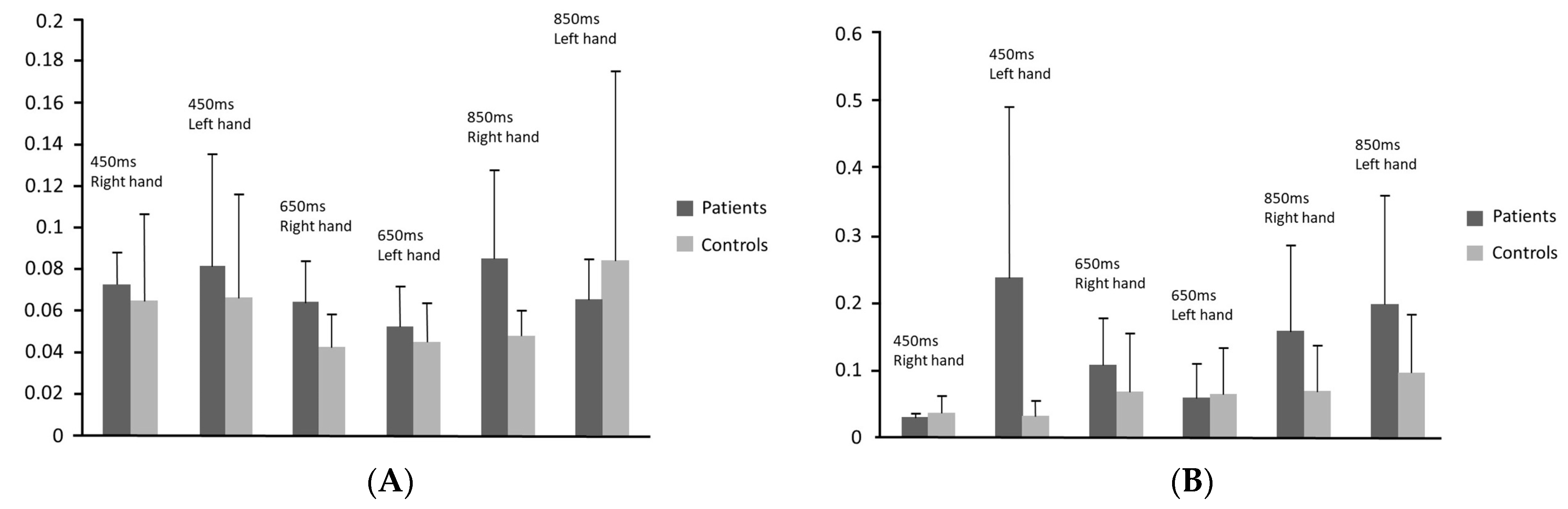
- Differences between right and left hand were detected only in patients who showed increased variability in inter-tone intervals reproduction with the right hand.
- (b)
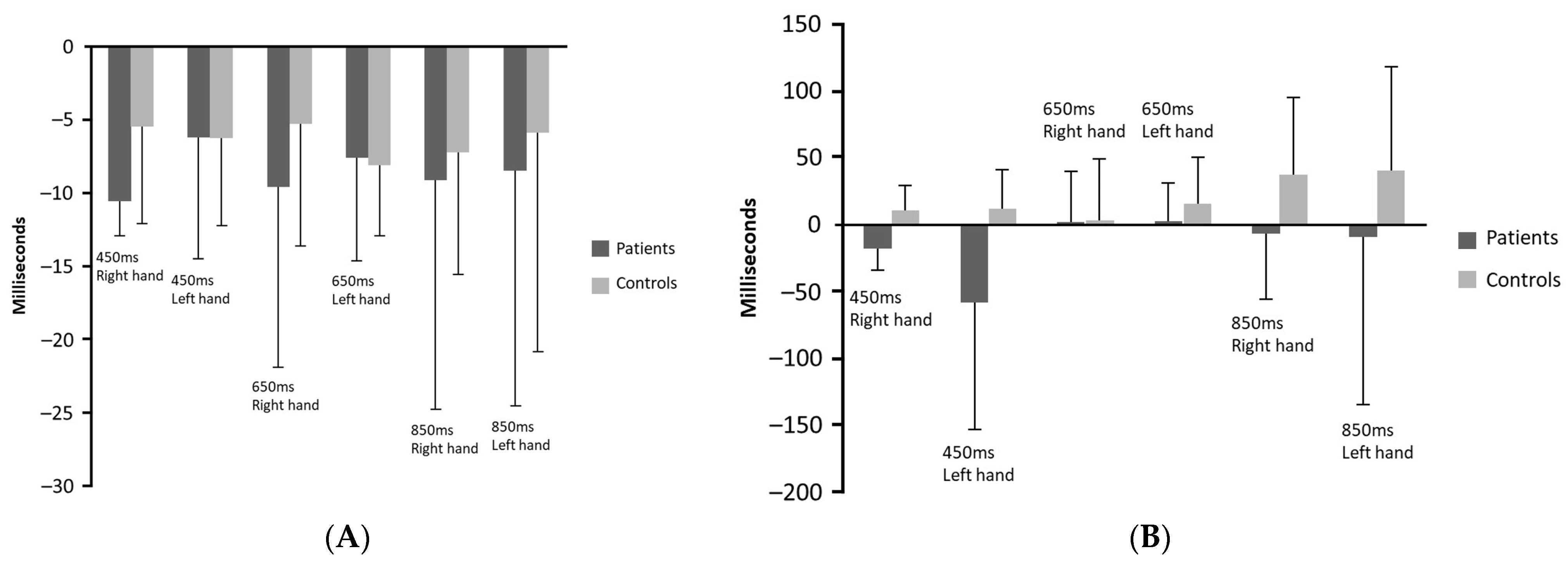
- No significant differences between the synchronisation and the continuation phase were detected in accuracy of tapping performance of the patient group. However, significantly greater variability of the tempo reproduced was detected in patients in the continuation phase. Healthy controls had significant difference in accuracy between the two phases (delayed responses to the stimuli tempo in the synchronisation phase versus accelerated responses in the continuation phase), whereas no significant differences between the synchronisation and continuation phases were found in variability of tapping performance.
- (a)
- The variability of tapping performance was greater in patients in both the synchronisation [F(5, 34) = 10.07, p < 0.001] and the continuation phase [F(5, 34) = 8.78, p < 0.001].
- (b)
- Patients had delayed responses compared to the control group particularly in the continuation phase [F(5, 34) = 5.23, p = 0.001]. It is noteworthy that in this phase, the healthy participants reproduced accelerated responses in comparison to the stimuli inter-tone intervals.
- (c)
- More precisely, patients delayed significantly compared to healthy participants when tapping with the left hand in the 450 ms tempo in the continuation phase and when tapping with the right hand in 450 ms tempo in both phases and in 850 ms tempo in the continuation phase. Patients had significantly greater variability of performance compared to control group when tapping with the right hand in 650 ms inter-tone interval in the synchronisation phase and in the 850 ms inter-tone interval in both phases as well as when tapping with the left hand in 450 ms in both phases and in 850 ms inter-tone interval in the continuation phase.
4. Discussion
- Greater variability and poorer accuracy than healthy individuals in overall tapping performance;
- Greater variability of their responses in both synchronisation and continuation phases compared to healthy participants;
- Lagged tapping rates only in the continuation phase compared to healthy participants (patients reproduced slower tapping rates, whereas healthy participants faster ITIs than the stimuli tempo);
- Significantly poorer performance in terms of variability in the continuation than the synchronisation phase, whereas there was no significant difference in healthy participants’ performance between the two phases of the task;
- No significant difference in the accuracy between the two phases of the task, whereas healthy participants had significantly accelerated responses in the continuation phase.
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fontes, R.; Ribeiro, J.; Gupta, D.S.; Machado, D.; Lopes-Júnior, F.; Magalhães, F.; Bastos, V.H.; Rocha, K.; Marinho, V.; Lima, G.; et al. Time Perception Mechanisms at Central Nervous System. Neurol. Int. 2016, 8, 5939. [Google Scholar] [CrossRef]
- Coull, J.; Nobre, A. Dissociating explicit timing from temporal expectation with fMRI. Curr. Opin. Neurobiol. 2008, 18, 137–144. [Google Scholar] [CrossRef]
- Coull, J.T.; Cheng, R.K.; Meck, W.H. Neuroanatomical and neurochemical substrates of timing. Neuropsychopharmacology 2011, 36, 3–25. [Google Scholar]
- Bégel, V.; Benoit, C.E.; Correa, A.; Cutanda, D.; Kotz, S.A.; Dalla Bella, S. “Lost in time” but still moving to the beat. Neuropsychologia 2017, 94, 129–138. [Google Scholar] [CrossRef]
- Koch, G.; Oliveri, M.; Torriero, S.; Salerno, S.; Lo Gerfo, E.; Caltagirone, C. Repetitive TMS of cerebellum interferes with millisecond time processing. Expir. Brain Res. 2007, 179, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Schwartze, M.; Tavano, A.; Schröger, E.; Kotz, S. Temporal aspects of prediction in audition: Cortical and subcortical neural mechanisms. Int. J. Psychophysiol. 2012, 83, 200–207. [Google Scholar]
- Rammsayer, T.H.; Borter, N.; Troche, S.J. Visual-auditory differences in duration discrimination of intervals in the subsecond and second range. Front. Psychol. 2015, 26, 1626. [Google Scholar] [CrossRef]
- Ferrill, M.; Love, T.; Walenski, M.; Shapiro, L.P. The time-course of lexical activation during sentence comprehension in people with aphasia. Am. J. Speech-Lang. Pathol. 2012, 21, S179–S189. [Google Scholar] [CrossRef]
- Blumstein, S.E.; Cooper, W.E.; Zurif, E.B.; Caramazza, A. The perception and production of voice-onset time in aphasia. Neuropsychologia 1977, 15, 371–383. [Google Scholar] [CrossRef]
- Van Lancker Sidtis, D.; Kempler, D.; Jackson, C.; Metter, E.M. Prosodic changes in aphasic speech: Timing. Clin. Linguist. Phon. 2010, 24, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Matchin, W.; Hickok, G. The Cortical Organization of Syntax. Cereb. Cortex 2020, 30, 1481–1498. [Google Scholar] [CrossRef] [PubMed]
- Sahin, N.T.; Pinker, S.; Cash, S.S.; Schomer, D.; Halgren, E. Sequential processing of lexical, grammatical, and phonological information within Broca’s area. Science 2009, 326, 445–449. [Google Scholar] [CrossRef]
- Grodzinsky, Y.; Santi, A. The battle for Broca’s region. Trends Cogn. Sci. 2008, 12, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Fadiga, L.; Craighero, L.; D’Ausilio, A. Broca’s area in language, action, and music. Ann. N. Y. Acad. Sci. 2009, 1169, 448–458. [Google Scholar] [CrossRef]
- Clerget, E.; Badets, A.; Duqué, J.; Olivier, E. Role of Broca’s area in motor sequence programming: A cTBS study. Neuroreport 2011, 22, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, P. Uniquely Human: The Evolution of Speech, Thought, and Selfless Behavior; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Tallal, P.; Miller, S.; Fitch, R. Neurobiological basis of speech: A case for the preeminence of temporal processing. Ann. N. Y. Acad. Sci. 1993, 682, 27–47. [Google Scholar] [CrossRef]
- Wittmann, M.; Burtscher, A.; Fries, W.; von Steinbüchel, N. Effects of brain-lesion size and location on temporal-order judgment. Neuroreport 2004, 15, 2401–2405. [Google Scholar] [CrossRef]
- Platel, H.; Price, C.; Baron, J.-C.; Wise, R.; Lambert, J.; Frackowiak, R.S.; Lechevalier, B.; Eustache, F. The structural components of music perception. A functional anatomical study. Brain 1997, 120, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, J.R.; Bookheimer, S.Y. Dissociating neural mechanisms of temporal sequencing and processing phonemes. Neuron 2003, 38, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Binder, M. Neural correlates of audiovisual temporal processing—Comparison of temporal order and simultaneity judgments. Neuroscience 2015, 300, 432–447. [Google Scholar] [CrossRef]
- Chapin, H.L.; Zanto, T.; Jantzen, K.J.; Kelso, S.J.; Steinberg, F.; Large, E.W. Neural responses to complex auditory rhythms: The role of attending. Front. Psychol. 2010, 1, 224. [Google Scholar] [CrossRef] [PubMed]
- Wencil, E.B.; Coslett, H.B.; Aguirre, G.K.; Chatterjee, A. Carving the clock at its component joints: Neural bases for interval timing. J. Neurophysiol. 2010, 104, 160–168. [Google Scholar] [CrossRef]
- Coull, J.T.; Davranche, K.; Nazarian, B.; Vidal, F. Functional anatomy of timing differs for production versus prediction of time intervals. Neuropsychologia 2013, 51, 309–319. [Google Scholar] [CrossRef]
- Carver, F.W.; Elvevåg, B.; Altamura, M.; Weinberger, D.R.; Coppola, R. The neuromagnetic dynamics of time perception. PLoS ONE 2012, 7, e42618. [Google Scholar] [CrossRef]
- Carpenter, R.L.; Rutherford, D.R. Acoustic cue discrimination in adult aphasia. J. Speech Lang. Hear. Res. 1973, 16, 534–544. [Google Scholar] [CrossRef]
- Sasanuma, S.; Tatsumi, I.F.; Kiritani, S.; Fujisaki, H. Auditory perception of signal duration in aphasic patients. Res. Inst. Logop. Phoniatr. Annu. Bull. 1973, 7, 65–72. [Google Scholar]
- Tallal, P.; Piercy, M. Developmental aphasia: The perception of brief vowels and extended stop consonants. Neuropsychologia 1975, 13, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Stefanatos, G.A.; Braitman, L.E.; Madigan, S. Fine grain temporal analysis in aphasia: Evidence from auditory gap detection. Neuropsychologia 2007, 45, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Efron, R. Temporal perception, aphasia and déjà vu. Brain 1963, 86, 403–424. [Google Scholar] [CrossRef]
- von Steinbόchel, N.; Wittmann, M.; Strasburger, H.; Szelag, E. Auditory temporal-order judgement is impaired in patients with cortical lesions in posterior regions of the left hemisphere. Neurosci. Lett. 1999, 264, 168–171. [Google Scholar] [CrossRef]
- Fink, M.; Churan, J.; Wittmann, M. Temporal processing and context dependency of phoneme discrimination in patients with aphasia. Brain Lang. 2006, 98, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Mohan, B.M.; Pavithra, K.; Naveen, C.P. Exploring (central) auditory processing deficits in individuals with Broca’s aphasia: Based on case study. Indian J. Otol. 2016, 22, 135–138. [Google Scholar] [CrossRef]
- Stefaniak, J.D.; Lambon Ralph, M.A.; De Dios Perez, B.; Griffiths, T.D.; Grube, M. Auditory beat perception is related to speech output fluency in post-stroke aphasia. Sci. Rep. 2021, 11, 3168. [Google Scholar] [CrossRef]
- Alcock, K.J.; Wade, D.; Anslow, P.; Passingham, R.E. Pitch and timing abilities in adult left-hemisphere-dysphasic and right-hemisphere-damaged subjects. Brain Lang. 2000, 75, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Zipse, L.; Worek, A.; Guarino, J.A.; Hufnagel, S.S. Tapped out: Do people with aphasia have rhythm processing deficits? J. Speech Lang. Hear. Res. 2014, 57, 2234–2245. [Google Scholar] [CrossRef] [PubMed]
- Coslett, H.B.; Shenton, J.; Dyer, T.; Wiener, M. Cognitive timing: Neuropsychology and anatomic basis. Brain Res. 2009, 1254, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Binkofski, F.; Buccino, G. Motor functions of the Broca’s region. Brain Lang. 2004, 89, 362–369. [Google Scholar] [CrossRef]
- Christianson, M.; Leathem, J. Development and standardisation of the computerised finger tapping test: Com-parison with other finger tapping instruments. N. Z. J. Psychol. 2004, 33, 44–49. [Google Scholar]
- Witt, S.T.; Laird, A.R.; Meyerand, M.E. Functional neuroimaging correlates of finger-tapping task variations: An ALE meta-analysis. NeuroImage 2008, 42, 343–356. [Google Scholar]
- Rao, S.M.; Harrington, D.L.; Haaland, K.Y.; Bobholz, J.A.; Cox, R.W.; Binder, J.R. Distributed neural systems underlying the timing of movements. J. Neurosci. 1997, 17, 5528–5535. [Google Scholar] [CrossRef]
- De Pretto, M.; Deiber, M.P.; James, C.E. Steady-state evoked potentials distinguish brain mechanisms of self-paced versus synchronization finger tapping. Hum. Mov. Sci. 2018, 61, 151–166. [Google Scholar] [CrossRef]
- Tsapkini, K.; Vlahou, C.H.; Potagas, C. Adaptation and validation of standardized aphasia tests in different languages: Lessons from the Boston Diagnostic Aphasia Examination—Short Form in Greek. Behav. Neurol. 2010, 22, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.C.; O’Donnell, F.B.; Shekhar, A.; Hetrick, P.W. Timing dysfunctions in schizophrenia as measured by repetitive finger tapping task. Brain Cogn. 2009, 71, 345–353. [Google Scholar] [CrossRef]
- Myers, L.S.; Gamst, G.; Guarino, A.J. Applied Multivariate Research: Design and Interpretation; Sage Publications: Thousand Oaks, CA, USA, 2006. [Google Scholar]
- Kolers, P.A.; Brewster, J.M. Rhythms and responses. J. Exp. Psychol. Hum. Percept. Perform. 1985, 11, 150–167. [Google Scholar] [CrossRef]
- Truman, G.; Hammond, G.R. Temporal regularity of tapping by the left and right hands in timed and untimed finger tapping. J. Mot. Behav. 1990, 22, 521–535. [Google Scholar] [CrossRef]
- Flach, R. The transition from synchronization to continuation tapping. Hum. Mov. Sci. 2005, 24, 465–483. [Google Scholar] [CrossRef]
- Elazary, A.S.; Attia, R.; Bergman, H.; Ben-Pazi, H. Age-related accelerated tapping response in healthy population. Percept. Mot. Ski. 2003, 96, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Szelag, E.; von Steinbüchel, N.; Pöppel, E. Temporal processing disorders in patients with Broca’s aphasia. Neurosci. Lett. 1997, 235, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Wing, A.M. The long and short of timing in response sequences. In Tutorials in Motor Behavior; Stelmach, G.E., Requin, J., Eds.; North-Holland: Amsterdam, The Netherlands, 1980; pp. 469–486. [Google Scholar]
- Mayer, J.F.; Murray, L.L. Measuring working memory deficits in aphasia. J. Commun. Disord. 2012, 45, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Choinski, M.; Szelag, E.; Wolak, T.; Szymaszek, A. Working Memory in Aphasia: The Role of Temporal Information Processing. Front. Hum. Neurosci. 2020, 14, 589802. [Google Scholar] [CrossRef] [PubMed]
- Potagas, C.; Kasselimis, D.; Evdokimidis, I. Short-term and working memory impairments in aphasia. Neuropsychologia 2011, 49, 2874–2878. [Google Scholar] [CrossRef]
- Schubotz, R.I.; Friederici, A.D.; von Cramon, D.Y. Time perception and motor timing: A common cortical and subcortical basis revealed by fMRI. NeuroImage 2000, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schubotz, R.I.; von Cramon, D.Y. Interval and ordinal properties of sequences are associated with distinct premotor areas. Cereb. Cortex 2001, 11, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Ullén, F.; Bengtsson, S.L.; Ehrsson, H.H.; Forssberg, H. Neural control of rhythmic sequences. Ann. N. Y. Acad. Sci. 2005, 1060, 368–376. [Google Scholar] [CrossRef]
- Thaut, M.H.; Trimarchi, P.D.; Parsons, L.M. Human brain basis of musical rhythm perception: Common and distinct neural substrates for meter, tempo, and pattern. Brain Sci. 2014, 4, 428–452. [Google Scholar] [CrossRef] [PubMed]
- Pouthas, V.; George, N.; Poline, J.B.; Pfeuty, M.; Vandemoorteele, P.F.; Hugueville, L.; Ferrandez, A.M.; Lehéricy, S.; Lebihan, D.; Renault, B. Neural network involved in time perception: An fMRI study comparing long and short interval estimation. Hum. Brain Mapp. 2005, 25, 433–441. [Google Scholar] [CrossRef]
- Livesey, A.C.; Wall, M.B.; Smith, A.T. Time perception: Manipulation of task difficulty dissociates clock functions from other cognitive demands. Neuropsychologia 2007, 45, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Clos, M.; Amunts, K.; Laird, A.R.; Fox, P.T.; Eickhoff, S.B. Tackling the multifunctional nature of Broca’s region meta-analytically: Co-activation-based parcellation of area 44. NeuroImage 2013, 83, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Stahl, B.; Kotz, S.A.; Henseler, I.; Turner, R.; Geyer, S. Rhythm in disguise: Why singing may not hold the key to recovery from aphasia. Brain 2011, 134, 3083–3093. [Google Scholar] [CrossRef]
- Zumbansen, A.; Peretz, I.; Hébert, S. The Combination of Rhythm and Pitch Can Account for the Beneficial Effect of Melodic Intonation Therapy on Connected Speech Improvements in Broca’s Aphasia. Front. Hum. Neurosci. 2014, 8, 592. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Effect | Measure | df | F | p |
|---|---|---|---|---|
| Group | Accuracy | 1.38 | 6.886 | 0.012 |
| Variability | 1.38 | 9.684 | 0.004 |
| Effect | Wlik’s Λ | Measure | df | F | p |
|---|---|---|---|---|---|
| Hand | 0.801 | Variability | 1.38 | 7.320 | 0.010 |
| Phase | 0.438 | Accuracy | 1.38 | 8.623 | 0.006 |
| Variability | 1.38 | 13.338 | 0.001 | ||
| Tempo | 0.431 | Accuracy | 2.37 | 6.864 | 0.002 |
| Variability | 2.37 | 14.523 | <0.001 | ||
| Phase × Group | 0.749 | Variability | 1.38 | 12.260 | 0.001 |
| Tempo × Group | 0.670 | Accuracy | 2.76 | 4.517 | 0.014 |
| Variability | 2.76 | 5.238 | 0.007 | ||
| Hand × Phase | 0.812 | Variability | 1.38 | 6.885 | 0.012 |
| Hand × Phase × Group | 0.763 | Variability | 1.38 | 11.398 | 0.002 |
| Hand × Tempo | 0.581 | Variability | 2.76 | 8.154 | 0.001 |
| Hand × Tempo × Group | 0.625 | Variability | 2.76 | 10.234 | <0.001 |
| Phase × Tempo | 0.368 | Variability | 2.76 | 7.913 | 0.001 |
| Phase × Tempo × Group | 0.631 | Variability | 2.76 | 8.478 | <0.001 |
| Hand × Phase × Tempo | 0.624 | Variability | 2.76 | 9.282 | <0.001 |
| Hand × Phase × Tempo × Group | 0.513 | Variability | 2.76 | 8.944 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andronoglou, C.; Konstantakopoulos, G.; Simoudi, C.; Kasselimis, D.; Evdokimidis, I.; Tsoukas, E.; Tsolakopoulos, D.; Angelopoulou, G.; Potagas, C. Is There a Role of Inferior Frontal Cortex in Motor Timing? A Study of Paced Finger Tapping in Patients with Non-Fluent Aphasia. NeuroSci 2023, 4, 235-246. https://doi.org/10.3390/neurosci4030020
Andronoglou C, Konstantakopoulos G, Simoudi C, Kasselimis D, Evdokimidis I, Tsoukas E, Tsolakopoulos D, Angelopoulou G, Potagas C. Is There a Role of Inferior Frontal Cortex in Motor Timing? A Study of Paced Finger Tapping in Patients with Non-Fluent Aphasia. NeuroSci. 2023; 4(3):235-246. https://doi.org/10.3390/neurosci4030020
Chicago/Turabian StyleAndronoglou, Chrysanthi, George Konstantakopoulos, Christina Simoudi, Dimitrios Kasselimis, Ioannis Evdokimidis, Evangelos Tsoukas, Dimitrios Tsolakopoulos, Georgia Angelopoulou, and Constantin Potagas. 2023. "Is There a Role of Inferior Frontal Cortex in Motor Timing? A Study of Paced Finger Tapping in Patients with Non-Fluent Aphasia" NeuroSci 4, no. 3: 235-246. https://doi.org/10.3390/neurosci4030020