
A Critical Review of the Deviance Detection Theory of Mismatch Negativity


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
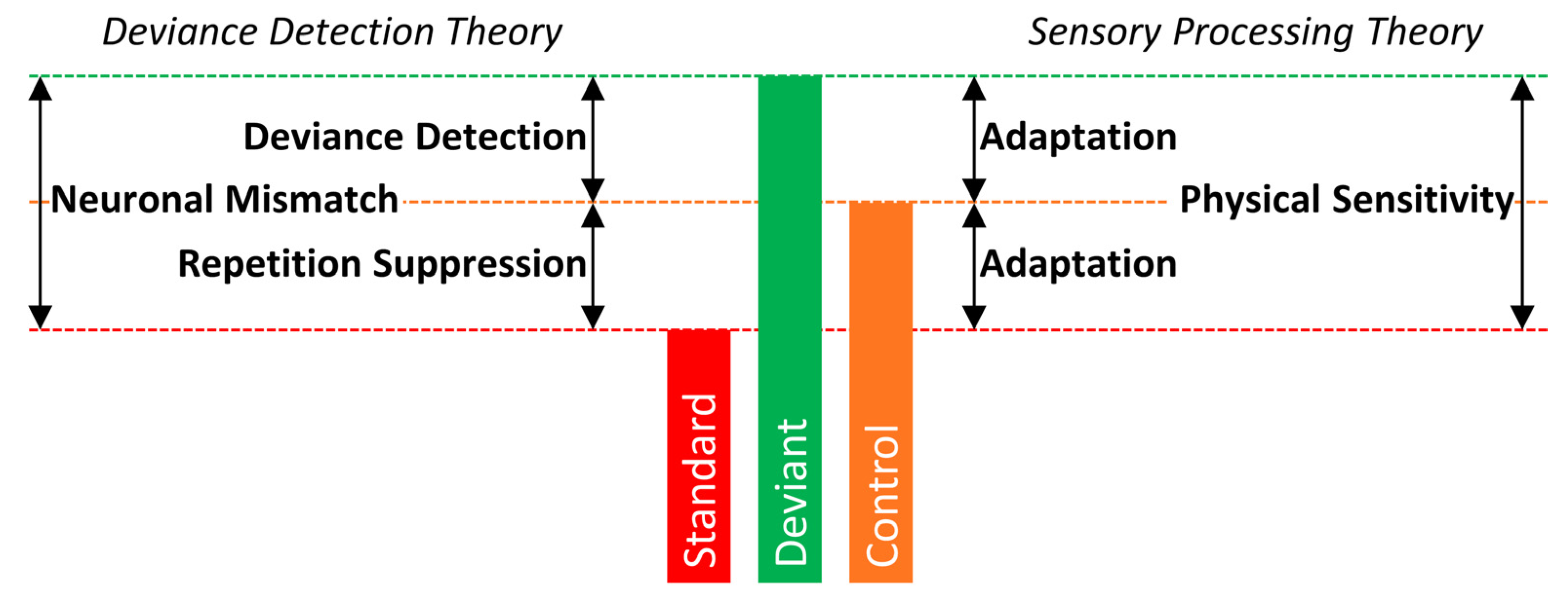
2. Deviance Detection Theory
3. Fresh Afferents Theory
4. Sensory Processing Theory
5. Historical Perspective of MMN Research
6. Clinical Applications of MMN
7. Limitations Affecting Interpretation of MMN as Evidence of the Deviance Detection Theory
7.1. Passive Oddball Paradigm
7.2. Control Paradigms
7.3. Electrophysiology
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fitzgerald, K.; Todd, J. Making Sense of Mismatch Negativity. Front. Psychiatry 2020, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Kujala, T.; Light, G. Mismatch Negativity: A Window to the Brain; Oxford University Press: Oxford, UK, 2019; ISBN 9780198705079. [Google Scholar]
- Ross, J.M.; Hamm, J.P. Cortical Microcircuit Mechanisms of Mismatch Negativity and Its Underlying Subcomponents. Front. Neural Circuits 2020, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Pakarinen, S.; Takegata, R.; Rinne, T.; Huotilainen, M.; Näätänen, R. Measurement of extensive auditory discrimination profiles using the mismatch negativity (MMN) of the auditory event-related potential (ERP). Clin. Neurophysiol. 2007, 118, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Pakarinen, S.; Rinne, T.; Takegata, R. The mismatch negativity (MMN): Towards the optimal paradigm. Clin. Neurophysiol. 2004, 115, 140–144. [Google Scholar] [CrossRef]
- Frodl-Bauch, T.; Kathmann, N.; Möller, H.-J.; Hegerl, U. Dipole Localization and Test-Retest Reliability of Frequency and Duration Mismatch Negativity Generator Processes. Brain Topogr. 1997, 10, 3–8. [Google Scholar] [CrossRef]
- Molholm, S.; Martinez, A.; Ritter, W.; Javitt, D.C.; Foxe, J.J. The Neural Circuitry of Pre-attentive Auditory Change-detection: An fMRI Study of Pitch and Duration Mismatch Negativity generators. Cereb. Cortex 2004, 15, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, T. Left hemispheric dipole locations of the neuromagnetic mismatch negativity to frequency, intensity and duration deviants. Cogn. Brain Res. 2003, 16, 83–90. [Google Scholar] [CrossRef]
- An, H.; Auksztulewicz, R.; Kang, H.; Schnupp, J.W. Cortical mapping of mismatch responses to independent acoustic features. Hear. Res. 2020, 399, 107894. [Google Scholar] [CrossRef]
- Loewy, D.H.; Campbell, K.B.; Bastien, C. The mismatch negativity to frequency deviant stimuli during natural sleep. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 493–501. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, F.; Wang, L.; Wang, Y.; Wang, C.; Wang, Q.; Huang, L. Cortical Areas Associated With Mismatch Negativity: A Connectivity Study Using Propofol Anesthesia. Front. Hum. Neurosci. 2018, 12, 392. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Morlet, D.; Giard, M.-H. Mismatch Negativity and N100 in Comatose Patients. Audiol. Neurotol. 2000, 5, 192–197. [Google Scholar] [CrossRef]
- Yue, X.; Fan, Y.; Xue, F.; Brauth, S.E.; Tang, Y.; Fang, G. The First Call Note Plays a Crucial Role in Frog Vocal Communication. Sci. Rep. 2017, 7, 10128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Featherstone, R.E.; Melnychenko, O.; Siegel, S.J. Mismatch negativity in preclinical models of schizophrenia. Schizophr. Res. 2018, 191, 35–42. [Google Scholar] [CrossRef]
- Schall, U.; Müller, B.W.; Kärgel, C.; Güntürkün, O. Electrophysiological mismatch response recorded in awake pigeons from the avian functional equivalent of the primary auditory cortex. NeuroReport 2015, 26, 239–244. [Google Scholar] [CrossRef]
- Romanchek, B.A.; Scheifele, P.M. Investigating Atlantic bottlenose dolphin mismatch negativity response to pure tone stimuli. J. Acoust. Soc. Am. 2019, 145, 1776. [Google Scholar] [CrossRef]
- Fishman, Y.I. The Mechanisms and Meaning of the Mismatch Negativity. Brain Topogr. 2014, 27, 500–526. [Google Scholar] [CrossRef]
- Light, G.A.; Näätänen, R. Mismatch negativity is a breakthrough biomarker for understanding and treating psychotic disorders. Proc. Natl. Acad. Sci. USA 2013, 110, 15175–15176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etodd, J.; Eharms, L.; Eschall, U.; Michie, P.T. Mismatch Negativity: Translating the Potential. Front. Psychiatry 2013, 4, 171. [Google Scholar] [CrossRef] [Green Version]
- Tada, M.; Kirihara, K.; Mizutani, S.; Uka, T.; Kunii, N.; Koshiyama, D.; Fujioka, M.; Usui, K.; Nagai, T.; Araki, T.; et al. Mismatch negativity (MMN) as a tool for translational investigations into early psychosis: A review. Int. J. Psychophysiol. 2019, 145, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Gaillard, A.; Mäntysalo, S. Early selective-attention effect on evoked potential reinterpreted. Acta Psychol. 1978, 42, 313–329. [Google Scholar] [CrossRef]
- Näätänen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef] [PubMed]
- Rauschecker, J.P.; Romanski, L.M. Auditory Cortical Organization: Evidence for Functional Streams. In The Auditory Cortex; Springer: Boston, MA, USA, 2010; pp. 99–116. [Google Scholar]
- Wacongne, C.; Changeux, J.-P.; Dehaene, S. A Neuronal Model of Predictive Coding Accounting for the Mismatch Negativity. J. Neurosci. 2012, 32, 3665–3678. [Google Scholar] [CrossRef]
- Garrido, M.I.; Kilner, J.M.; Stephan, K.E.; Friston, K.J. The mismatch negativity: A review of underlying mechanisms. Clin. Neurophysiol. 2009, 120, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaaskelainen, I.P.; Ahveninen, J.; Bonmassar, G.; Dale, A.M.; Ilmoniemi, R.J.; Levanen, S.; Lin, F.-H.; May, P.; Melcher, J.; Stufflebeam, S.; et al. Human posterior auditory cortex gates novel sounds to consciousness. Proc. Natl. Acad. Sci. USA 2004, 101, 6809–6814. [Google Scholar] [CrossRef] [Green Version]
- May, P.J.C.; Tiitinen, H. Mismatch negativity (MMN), the deviance-elicited auditory deflection, explained. Psychophysiology 2010, 47, 66–122. [Google Scholar] [CrossRef] [PubMed]
- Kurkela, J.L.O.; Lipponen, A.; Kyläheiko, I.; Astikainen, P. Electrophysiological evidence of memory-based detection of auditory regularity violations in anesthetized mice. Sci. Rep. 2018, 8, 3027. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Kujala, T.; Escera, C.; Baldeweg, T.; Kreegipuu, K.; Carlson, S.; Ponton, C. The mismatch negativity (MMN)—A unique window to disturbed central auditory processing in ageing and different clinical conditions. Clin. Neurophysiol. 2012, 123, 424–458. [Google Scholar] [CrossRef]
- Naatanen, R.; Jacobsen, T.; Winkler, I. Memory-based or afferent processes in mismatch negativity (MMN): A review of the evidence. Psychophysiology 2005, 42, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Harms, L.; Michie, P.T.; Näätänen, R. Criteria for determining whether mismatch responses exist in animal models: Focus on rodents. Biol. Psychol. 2016, 116, 28–35. [Google Scholar] [CrossRef]
- Duque, D.; Wang, X.; Nieto-Diego, J.; Krumbholz, K.; Malmierca, M.S. Neurons in the inferior colliculus of the rat show stimulus-specific adaptation for frequency, but not for intensity. Sci. Rep. 2016, 6, 24114. [Google Scholar] [CrossRef] [Green Version]
- Harpaz, M.; Jankowski, M.M.; Khouri, L.; Nelken, I. Emergence of abstract sound representations in the ascending auditory system. Prog. Neurobiol. 2021, 102049. [Google Scholar] [CrossRef]
- Jacobsen, T.; Schröger, E. Measuring duration mismatch negativity. Clin. Neurophysiol. 2003, 114, 1133–1143. [Google Scholar] [CrossRef]
- Picton, T.W.; Woods, D.L.; Baribeau Braun, J.; Healey, T.M.G. Evoked potential audiometry. J. Otolaryngol. 1977, 6, 90–119. [Google Scholar]
- Park, Y.-M.; Lee, S.-H.; Kim, S.; Bae, S.-M. The loudness dependence of the auditory evoked potential (LDAEP) in schizophrenia, bipolar disorder, major depressive disorder, anxiety disorder, and healthy controls. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Juckel, G. Serotonin: From sensory processing to schizophrenia using an electrophysiological method. Behav. Brain Res. 2015, 277, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Kopp-Scheinpflug, C.; Sinclair, J.L.; Linden, J.F. When Sound Stops: Offset Responses in the Auditory System. Trends Neurosci. 2018, 41, 712–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picton, T.W.; Hillyard, S.A.; Galambos, R. Habituation and Attention in the Auditory System. In Auditory System; Springer: Berlin/Heidelberg, Germany, 1976; pp. 343–389. [Google Scholar] [CrossRef]
- Näätänen, R.; Hukkanen, S.; Järvilehto, T.; Näätínen, R. Magnitude of Stimulus Deviance and Brain Potentials. Prog. Brain Res. 1980, 54, 337–342. [Google Scholar] [CrossRef]
- Ulanovsky, N.; Las, L.; Farkas, D.; Nelken, I. Multiple Time Scales of Adaptation in Auditory Cortex Neurons. J. Neurosci. 2004, 24, 10440–10453. [Google Scholar] [CrossRef] [PubMed]
- Latimer, K.W.; Barbera, D.; Sokoletsky, M.; Awwad, B.; Katz, Y.; Nelken, I.; Lampl, I.; Fairhall, A.L.; Priebe, N.J. Multiple Timescales Account for Adaptive Responses across Sensory Cortices. J. Neurosci. 2019, 39, 10019–10033. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.A.; Conway, B.A. Classical and controlled auditory mismatch responses to multiple physical deviances in anaesthetised and conscious mice. Eur. J. Neurosci. 2020, 15072. [Google Scholar] [CrossRef]
- Schröger, E.; Wolff, C. Mismatch response of the human brain to changes in sound location. NeuroReport 1996, 7, 3005–3008. [Google Scholar] [CrossRef] [PubMed]
- Koshiyama, D.; Kirihara, K.; Tada, M.; Nagai, T.; Fujioka, M.; Usui, K.; Araki, T.; Kasai, K. Reduced Auditory Mismatch Negativity Reflects Impaired Deviance Detection in Schizophrenia. Schizophr. Bull. 2020, 46, 937–946. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, T.; Kreitschmann-Andermahr, I. The effects of ketamine on the mismatch negativity (MMN) in humans—A meta-analysis. Clin. Neurophysiol. 2016, 127, 1387–1394. [Google Scholar] [CrossRef]
- Parras, G.G.; Nieto-Diego, J.; Carbajal, G.V.; Valdés-Baizabal, C.; Escera, C.; Malmierca, M.S. Neurons along the auditory pathway exhibit a hierarchical organization of prediction error. Nat. Commun. 2017, 8, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchsbaum, M.S. Average evoked response and stimulus intensity in identical and fraternal twins. Physiol. Psychol. 1974, 2, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, S. Science controversies past and present. Phys. Today 2011, 64, 39. [Google Scholar] [CrossRef] [Green Version]
- Ioannidis, J.P.A. Why Most Published Research Findings Are False. PLoS Med. 2005, 2, e124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, B.K.; Vuletich, H.A.; Lundberg, K.B. The Bias of Crowds: How Implicit Bias Bridges Personal and Systemic Prejudice. Psychol. Inq. 2017, 28, 233–248. [Google Scholar] [CrossRef]
- Danchev, V.; Rzhetsky, A.; Evans, J.A. Centralized scientific communities are less likely to generate replicable results. eLife 2019, 8, e43094. [Google Scholar] [CrossRef]
- Fryer, G. Scientific Method in Practice; Cambridge University Press: Cambridge, UK, 2003; ISBN 9780511815034. [Google Scholar] [CrossRef] [Green Version]
- Stahl, B.C. Critical Social Information Systems Research; Oxford University Press (OUP): Oxford, UK, 2011. [Google Scholar]
- Schnupp, J.; Nelken, I.; King, A. Auditory Neuroscience: Making Sense of Sound; MIT Press: Cambridge, MA, USA, 2011; ISBN 9780262518024. [Google Scholar]
- Joshi, Y.B.; Breitenstein, B.; Tarasenko, M.; Thomas, M.L.; Chang, W.-L.; Sprock, J.; Sharp, R.F.; Light, G.A. Mismatch negativity impairment is associated with deficits in identifying real-world environmental sounds in schizophrenia. Schizophr. Res. 2018, 191, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Sehatpour, P.; Hoptman, M.J.; Lakatos, P.; Dias, E.C.; Kantrowitz, J.T.; Martinez, A.M.; Javitt, D.C. Neural mechanisms of mismatch negativity dysfunction in schizophrenia. Mol. Psychiatry 2017, 22, 1585–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.; Shinn-Cunningham, B.; Tager-Flusberg, H. Meta-analysis and systematic review of the literature characterizing auditory mismatch negativity in individuals with autism. Neurosci. Biobehav. Rev. 2018, 87, 106–117. [Google Scholar] [CrossRef]
- Chen, T.-C.; Hsieh, M.H.; Lin, Y.-T.; Chan, P.-Y.S.; Cheng, C.-H. Mismatch negativity to different deviant changes in autism spectrum disorders: A meta-analysis. Clin. Neurophysiol. 2020, 131, 766–777. [Google Scholar] [CrossRef]
- Brima, T.; Molholm, S.; Molloy, C.J.; Sysoeva, O.V.; Nicholas, E.; Djukic, A.; Freedman, E.G.; Foxe, J.J. Auditory sensory memory span for duration is severely curtailed in females with Rett syndrome. Transl. Psychiatry 2019, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Bi, H.-Y. Auditory processing deficit in individuals with dyslexia: A meta-analysis of mismatch negativity. Neurosci. Biobehav. Rev. 2020, 116, 396–405. [Google Scholar] [CrossRef]
- Schwade, L.F.; Rosa, R.F.M.; Zen, P.R.G.; Sleifer, P. Mismatch negativity in children with complaints of learning difficulties. Rev. Logop. Foniatría Audiol. 2020, 40, 23–29. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Hsu, W.-Y.; Lin, Y.-Y. Effects of physiological aging on mismatch negativity: A meta-analysis. Int. J. Psychophysiol. 2013, 90, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Foxe, J.J.; Burke, K.M.; Andrade, G.N.; Djukic, A.; Frey, H.-P.; Molholm, S. Automatic cortical representation of auditory pitch changes in Rett syndrome. J. Neurodev. Disord. 2016, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Moser, J.; Schleger, F.; Weiss, M.; Sippel, K.; Dehaene-Lambertz, G.; Preissl, H. Magnetoencephalographic signatures of hierarchical rule learning in newborns. Dev. Cogn. Neurosci. 2020, 46, 100871. [Google Scholar] [CrossRef]
- Todd, J.; Michie, P.T.; Schall, U.; Karayanidis, F.; Yabe, H.; Näätänen, R. Deviant Matters: Duration, Frequency, and Intensity Deviants Reveal Different Patterns of Mismatch Negativity Reduction in Early and Late Schizophrenia. Biol. Psychiatry 2008, 63, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liao, X.; Li, R.; Liang, S.; Ding, R.; Li, J.; Zhang, J.; He, W.; Liu, K.; Pan, J.; et al. Single-neuron representation of learned complex sounds in the auditory cortex. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Sams, M.; Alho, K.; Näätänen, R. Short-Term Habituation and Dishabituation of the Mismatch Negativity of the ERP. Psychophysiology 1984, 21, 434–441. [Google Scholar] [CrossRef]
- Harms, L. Mismatch responses and deviance detection in N-methyl-D-aspartate (NMDA) receptor hypofunction and developmental models of schizophrenia. Biol. Psychol. 2016, 116, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Saloranta, A.; Alku, P.; Peltola, M.S. Listen-and-repeat training improves perception of second language vowel duration: Evidence from mismatch negativity (MMN) and N1 responses and behavioral discrimination. Int. J. Psychophysiol. 2020, 147, 72–82. [Google Scholar] [CrossRef]
- Polterovich, A.; Jankowski, M.M.; Nelken, I. Deviance sensitivity in the auditory cortex of freely moving rats. PLoS ONE 2018, 13, e0197678. [Google Scholar] [CrossRef]
- Baldeweg, T.; Klugman, A.; Gruzelier, J.; Hirsch, S.R. Mismatch negativity potentials and cognitive impairment in schizophrenia. Schizophr. Res. 2004, 69, 203–217. [Google Scholar] [CrossRef]
- Asadzadeh, S.; Rezaii, T.Y.; Beheshti, S.; Delpak, A.; Meshgini, S. A systematic review of EEG source localization techniques and their applications on diagnosis of brain abnormalities. J. Neurosci. Methods 2020, 339, 108740. [Google Scholar] [CrossRef]
- Deouell, L.Y. The Frontal Generator of the Mismatch Negativity Revisited. Psychophysiology 2007, 21, 188–203. [Google Scholar] [CrossRef]
- Hajizadeh, A.; Matysiak, A.; Brechmann, A.; König, R.; May, P.J.C. Why do humans have unique auditory event-related fields? Evidence from computational modeling and MEG experiments. Psychophysiology 2021, 58, e13769. [Google Scholar] [CrossRef]
- Astikainen, P.; Stefanics, G.; Nokia, M.; Lipponen, A.; Cong, F.; Penttonen, M.; Ruusuvirta, T. Memory-Based Mismatch Response to Frequency Changes in Rats. PLoS ONE 2011, 6, e24208. [Google Scholar] [CrossRef] [Green Version]
- Umbricht, D.; Vyssotki, D.; Latanov, A.; Nitsch, R.; Lipp, H.-P. Deviance-related electrophysiological activity in mice: Is there mismatch negativity in mice? Clin. Neurophysiol. 2005, 116, 353–363. [Google Scholar] [CrossRef]
- Nelken, I.; Ulanovsky, N. Mismatch Negativity and Stimulus-Specific Adaptation in Animal Models. Psychophysiology 2007, 21, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Michie, P.T.; Fulham, W.R.; Todd, J.; Budd, T.W.; Schall, U.; Hunter, M.; Hodgson, D.M. Epidural Auditory Event-Related Potentials in the Rat to Frequency and duration Deviants: Evidence of Mismatch Negativity? Front. Psychol. 2011, 2, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casado-Román, L.; Carbajal, G.V.; Pérez-González, D.; Malmierca, M.S. Prediction error signaling explains neuronal mismatch responses in the medial prefrontal cortex. PLoS Biol. 2020, 18, e3001019. [Google Scholar] [CrossRef]
- Lee, C.C.; Winer, J.A. A Synthesis of Auditory Cortical Connections: Thalamocortical, Commissural and Corticocortical Systems. In The Auditory Cortex; Springer: Boston, MA, USA, 2010; pp. 147–170. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Reilly, J.A.; O’Reilly, A. A Critical Review of the Deviance Detection Theory of Mismatch Negativity. NeuroSci 2021, 2, 151-165. https://doi.org/10.3390/neurosci2020011
O’Reilly JA, O’Reilly A. A Critical Review of the Deviance Detection Theory of Mismatch Negativity. NeuroSci. 2021; 2(2):151-165. https://doi.org/10.3390/neurosci2020011
Chicago/Turabian StyleO’Reilly, Jamie A., and Amonrat O’Reilly. 2021. "A Critical Review of the Deviance Detection Theory of Mismatch Negativity" NeuroSci 2, no. 2: 151-165. https://doi.org/10.3390/neurosci2020011