
Evaluation of Antidiabetic Potential of Mangifera indica Leaf in Streptozotocin-Induced Type 2 Diabetic Rats: Focus on Glycemic Control and Cholesterol Regulation
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Collection and Extract Preparation
2.2. In Vitro Insulin-Releasing Studies Using BRIN-BD11 Cells
2.3. In Vitro Insulin-Releasing Studies Using Isolated Mouse Islets
2.4. In Vitro Starch Digestion Using EEMI
2.5. In Vitro Glucose Diffusion Using EEMI
2.6. In Vitro DPPH Assay Using EEMI
2.7. Animals
2.8. Acute Effects of EEMI on Feeding Test
2.9. Acute Effects of EEMI Leaves on Oral Glucose Tolerance Test
2.10. Effect of EEMI on Body Weight, Fasting Blood Glucose, and Plasma Insulin
2.11. Effects of EEMI Leaves on Liver Glycogen Content
2.12. Effects of EEMI on Lipid Profile
2.13. Phytochemical Screening of EEMI
2.14. Statistical Analysis
3. Results
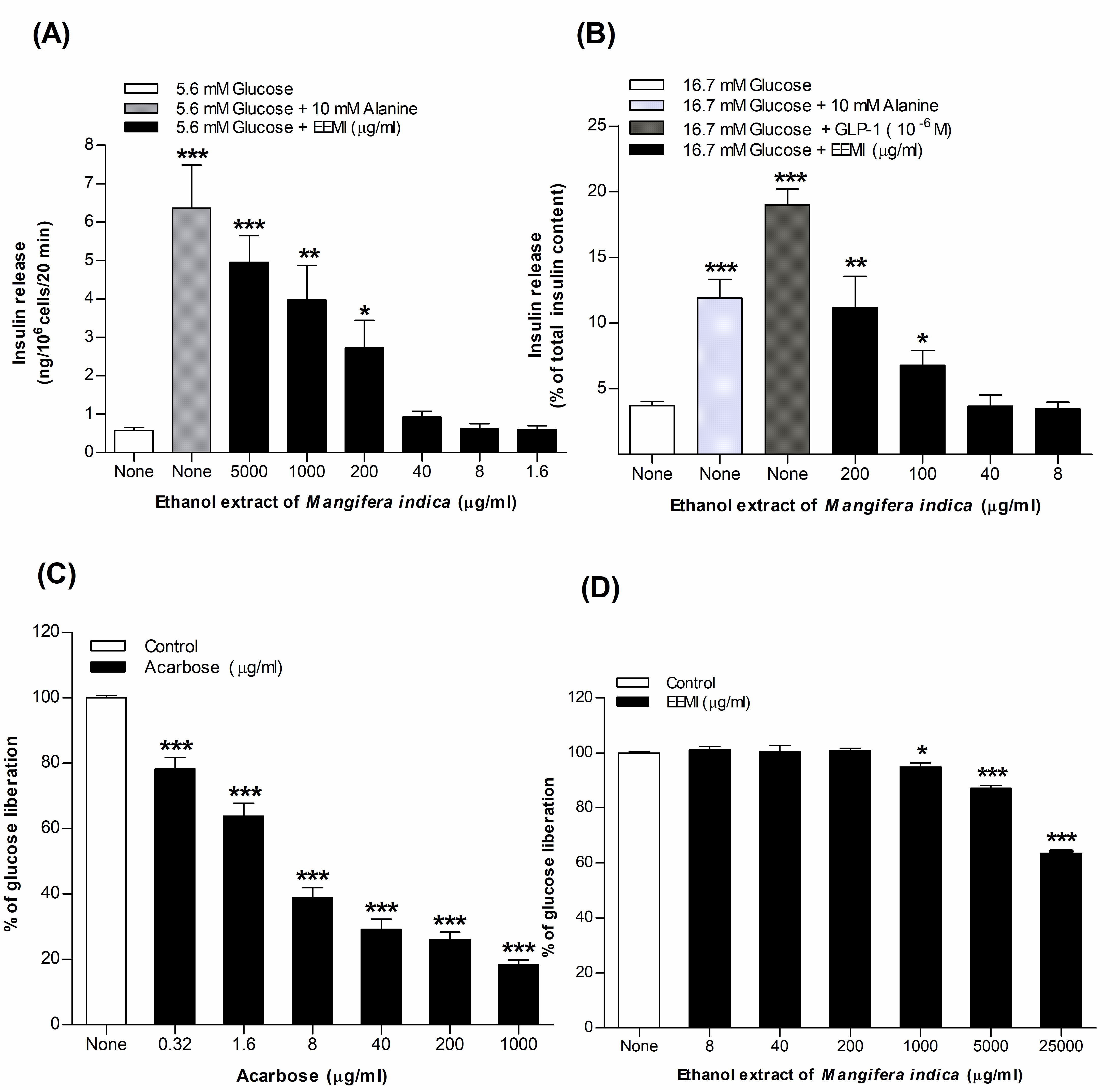
3.1. Insulin Release from BRIN-BD11 Cells Using EEMI
3.2. Insulin Release from Isolated Mouse Islets Using EEMI
3.3. Starch Digestion Using EEMI
3.4. DPPH Assay Using EEMI
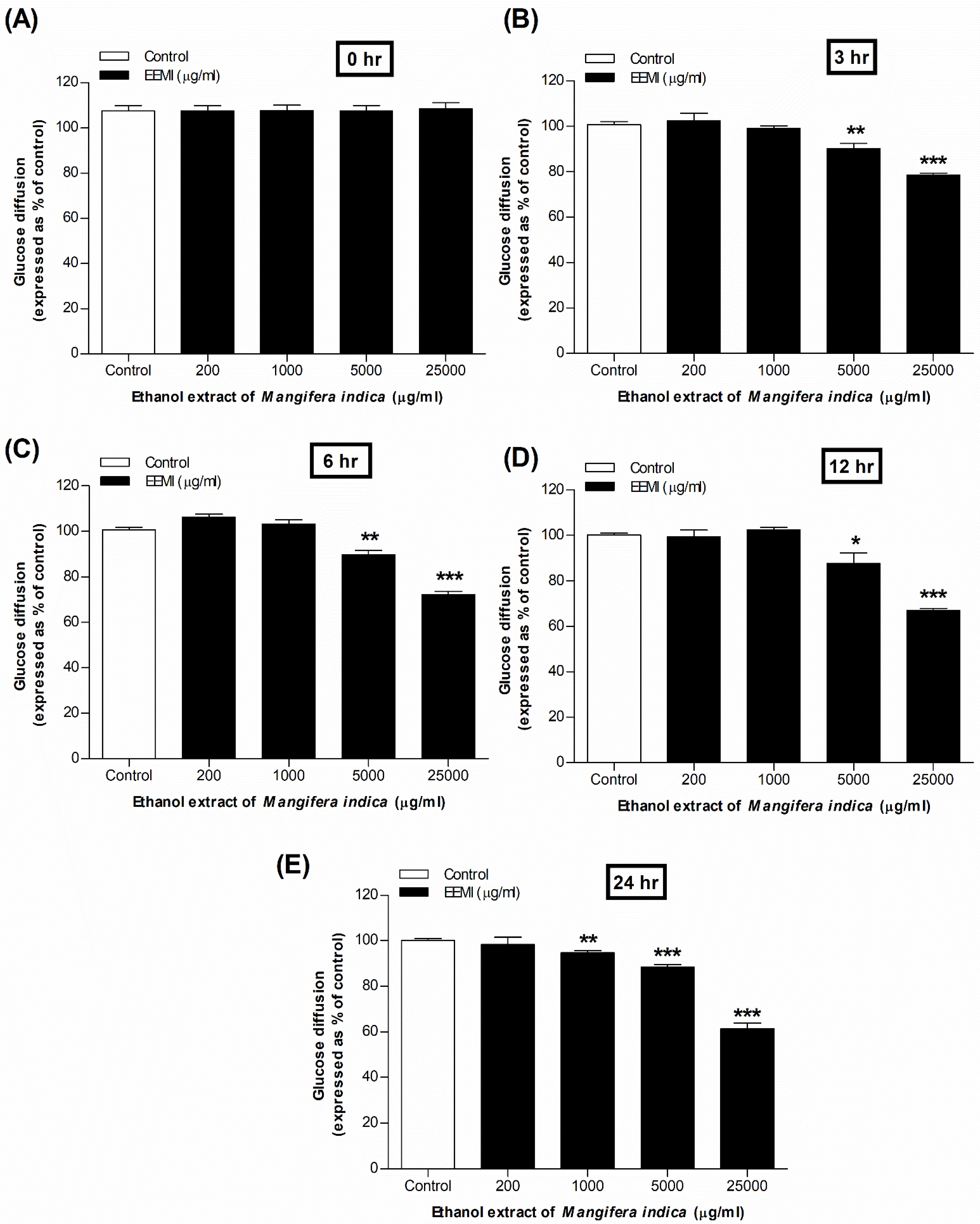
3.5. Glucose Diffusion Using EEMI
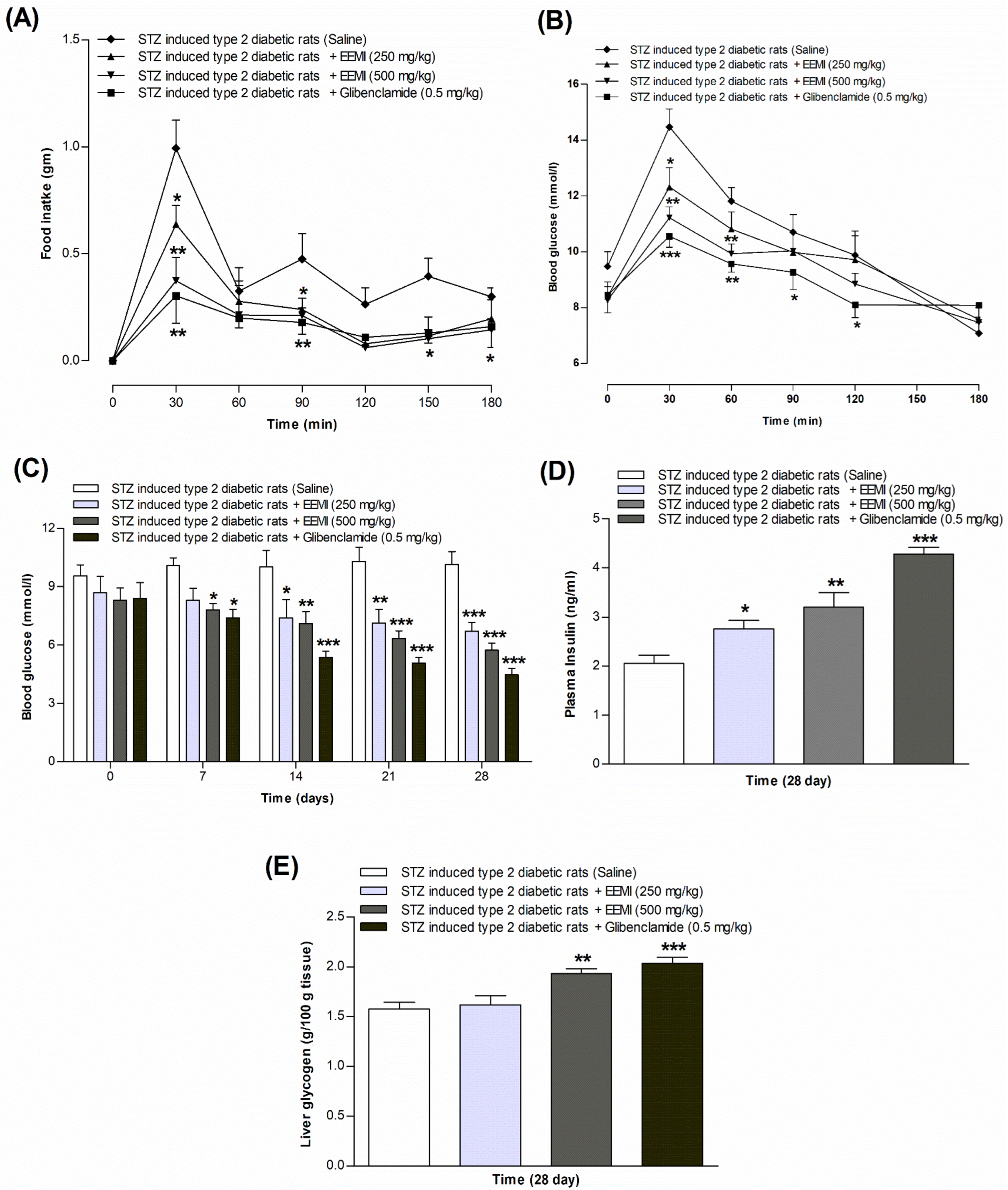
3.6. Feeding Test Using EEMI
3.7. Oral Glucose Tolerance Test Using EEMI
3.8. Fasting Blood Glucose, Plasma Insulin, and Liver Glycogen Using EEMI
3.9. Chronic Effects of Using EEMI
3.10. Phytochemical Screening Using EEMI
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BRIN-BD11 | Clonal pancreatic BRIN-BD11 β-cells |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| GLP-1 | Glucagon-Like Peptide 1 |
| HDL | High-Density Lipoprotein |
| KATP | ATP-sensitive K+ channel |
| LDL | Low-Density Lipoprotein |
| T2DM | Type 2 diabetes mellitus |
| VLDL | Very-Low-Density Lipoprotein |
References
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of Diabetes: An Overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Samia, J.F.; Khan, J.T.; Rafi, M.R.; Rahman, S.; Rahman, A.B.; Abdel-Wahab, Y.H.A.; Seidel, V. Protective Effects of Medicinal Plant-Based Foods against Diabetes: A Review on Pharmacology, Phytochemistry, and Molecular Mechanisms. Nutrients 2023, 15, 3266. [Google Scholar] [CrossRef] [PubMed]
- Fan, W. Epidemiology in Diabetes Mellitus and Cardiovascular Disease. Cardiovasc. Endocrinol. Metab. 2017, 6, 8–16. [Google Scholar] [CrossRef] [PubMed]
- George, M.M.; Copeland, K.C. Current Treatment Options for Type 2 Diabetes Mellitus in Youth: Today’s Realities and Lessons from the TODAY Study. Curr. Diabetes Rep. 2013, 13, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Abu-Odeh, A.M.; Talib, W.H. Middle East Medicinal Plants in the Treatment of Diabetes: A Review. Molecules 2021, 26, 742. [Google Scholar] [CrossRef] [PubMed]
- Sofowora, A.; Ogunbodede, E.; Onayade, A. The Role and Place of Medicinal Plants in the Strategies for Disease Prevention. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 210–229. [Google Scholar] [CrossRef] [PubMed]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P.A. Indian Herbs and Herbal Drugs Used for the Treatment of Diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Prasad, S.; Kumar, R.; Hemalatha, S. An overview on Antidiabetic Medicinal Plants Having Insulin Mimetic Property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Pang, G.-M.; Li, F.-X.; Yan, Y.; Zhang, Y.; Kong, L.-L.; Zhu, P.; Wang, K.-F.; Zhang, F.; Liu, B.; Lu, C. Herbal Medicine in the Treatment of Patients with Type 2 Diabetes Mellitus. Chin. Med. J. 2019, 132, 78–85. [Google Scholar] [CrossRef]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. A Review on Ethnopharmacological Applications, Pharmacological Activities, and Bioactive Compounds of Mangifera indica (Mango). Evid.-Based Complement. Altern. Med. 2017, 2017, 6949835. [Google Scholar] [CrossRef]
- Shah, K.; Patel, M.; Patel, R.; Parmar, P. Mangifera indica (Mango). Pharmacogn. Rev. 2010, 4, 42–48. [Google Scholar] [CrossRef]
- Akram, M.; Hamid, A.; Khalil, A.; Ghaffar, A.; Tayyaba, N.; Saeed, A.; Ali, M.; Naveed, A. Review on Medicinal Uses, Pharmacological, Phytochemistry and Immunomodulatory Activity of Plants. Int. J. Immunopathol. Pharmacol. 2014, 27, 313–319. [Google Scholar] [CrossRef]
- Saleem, M.; Tanvir, M.; Akhtar, M.F.; Iqbal, M.; Saleem, A. Antidiabetic Potential of Mangifera indica L. cv. Anwar Ratol Leaves: Medicinal Application of Food Wastes. Medicina 2019, 55, 353. [Google Scholar] [CrossRef] [PubMed]
- Aderibigbe, A.O.; Emudianughe, T.S.; Lawal, B.A.S. Evaluation of the antidiabetic action of Mangifera indica in mice. Phytotherapy Res. 2001, 15, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Yoopum, S.; Wongmanee, N.; Rojanaverawong, W.; Rattanapunya, S.; Sumsakul, W.; Hanchang, W. Mango (Mangifera indica L.) seed kernel extract suppresses hyperglycemia by modulating pancreatic β cell apoptosis and dysfunction and hepatic glucose metabolism in diabetic rats. Environ. Sci. Pollut. Res. 2023, 30, 123286–123308. [Google Scholar] [CrossRef] [PubMed]
- Meneses, M.A.; Caputo, G.; Scognamiglio, M.; Reverchon, E.; Adami, R. Antioxidant Phenolic Compounds Recovery from Mangifera indica L. by-Products by Supercritical Antisolvent Extraction. J. Food Eng. 2015, 163, 45–53. [Google Scholar] [CrossRef]
- McClenaghan, N.H.; Barnett, C.R.; Ah-Sing, E.; Abdel-Wahab, Y.H.A.; O’Harte, F.P.M.; Yoon, T.-W.; Swanston-Flatt, S.K.; Flatt, P.R. Characterization of a Novel Glucose-Responsive Insulin-Secreting Cell Line, BRIN-BD11, Produced by Electrofusion. Diabetes 1996, 45, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, Y.H.; Power, G.J.; Flatt, P.R.; Woodhams, D.C.; Rollins-Smith, L.A.; Conlon, J.M. A Peptide of the Phylloseptin Family from the Skin of the Frog Hylomantis Lemur (Phyllomedusinae) with Potent In Vitro and In Vivo Insulin-Releasing Activity. Peptides 2008, 29, 2136–2143. [Google Scholar] [CrossRef] [PubMed]
- Moran, B.M.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; McKillop, A.M. Evaluation of the Insulin-Releasing and Glucose-Lowering Effects of GPR120 Activation in Pancreatic β-Cells. Diabetes Obes. Metab. 2014, 16, 1128–1139. [Google Scholar] [CrossRef]
- Ansari, P.; Choudhury, S.T.; Abdel-Wahab, Y.H.A. Insulin Secretory Actions of Ethanol Extract of Eucalyptus citriodora Leaf, including Plasma DPP-IV and GLP-1 Levels in High-Fat-Fed Rats, as Well as Characterization of Biologically Effective Phytoconstituents. Metabolites 2022, 12, 757. [Google Scholar] [CrossRef]
- Ansari, P.; Hannan, J.; Seidel, V.; Abdel-Wahab, Y.H. Polyphenol-Rich Leaf of Annona squamosa Stimulates Insulin Release from BRIN-BD11 Cells and Isolated Mouse Islets, Reduces (CH2O)n Digestion and Absorption, and Improves Glucose Tolerance and GLP-1 (7-36) Levels in High-Fat-Fed Rats. Metabolites 2022, 12, 995. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Hannan, J.M.A.; Choudhury, S.T.; Islam, S.S.; Talukder, A.; Seidel, V.; Abdel-Wahab, Y.H.A. Antidiabetic Actions of Ethanol Extract of Camellia sinensis Leaf Ameliorates Insulin Secretion, Inhibits the DPP-IV Enzyme, Improves Glucose Tolerance, and Increases Active GLP-1 (7–36) Levels in High-Fat-Diet-Fed Rats. Medicines 2022, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Islam, S.S.; Akther, S.; Khan, J.T.; Shihab, J.A.; Abdel-Wahab, Y.H.A. Insulin Secretory Actions of Ethanolic Extract of Acacia Arabica Bark in High Fat-Fed Diet-Induced Obese Type 2 Diabetic Rats. Biosci. Rep. 2023, 43, BSR20230329. [Google Scholar] [CrossRef]
- Ansari, P.; Hannan, J.A.; Abdel-Wahab, Y.H.; Flatt, P.R. Antidiabetic and Insulinotropic Properties of Bark of Heritiera Fomes: Inhibits Starch Digestion, Protein Glycation, DPP-IV Activity, and Glucose Absorption in Gut. Planta Medica 2021, 87, 1252. [Google Scholar] [CrossRef]
- Ansari, P.; Azam, S.; Seidel, V.; Abdel-Wahab, Y.H.A. In Vitro and In Vivo Antihyperglycemic Activity of the Ethanol Extract of Heritiera Fomes Bark and Characterization of Pharmacologically Active Phytomolecules. J. Pharm. Pharmacol. 2022, 74, rgac010. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, V.; Milos, M.; Kulisic, T.; Jukic, M. Screening of 70 Medicinal Plant Extracts for Antioxidant Capacity and Total Phenols. Food Chem. 2006, 94, 550–557. [Google Scholar] [CrossRef]
- Chaves, N.; Santiago, A.; Alías, J.C. Quantification of the Antioxidant Activity of Plant Extracts: Analysis of Sensitivity and Hierarchization Based on the Method Used. Antioxidants 2020, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Lenzen, S. The Mechanisms of Alloxan- and Streptozotocin-Induced Diabetes. Diabetologia 2008, 51, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Shill, M.C.; Rakib, A.A.; Khan, S.I.; Hossain, M.; Alam, S.; Hossain, H.; Karmakar, U.K.; Bhuia, S.; Shahriar, M.; Reza, H.M.; et al. Polyphenol-Standardized Aphanamixis Polystachya Leaf Extract Ameliorates Diabetes, Oxidative Stress, Inflammation, and Fibrosis in Streptozotocin-Induced Diabetic Rats. J. Food Biochem. 2024, 2024, e9441968. [Google Scholar] [CrossRef]
- López-Soldado, I.; Guinovart, J.J.; Duran, J. Increasing Hepatic Glycogen Moderates the Diabetic Phenotype in Insulin-Deficient Akita Mice. J. Biol. Chem. 2021, 296, 100498. [Google Scholar] [CrossRef]
- Hannan, J.M.A.; Ansari, P.; Azam, S.; Flatt, P.R.; Wahab, Y.H.A.A. Effects of Spirulina Platensis on Insulin Secretion, Dipeptidyl Peptidase IV Activity and both Carbohydrate Digestion and Absorption Indicate Potential as an Adjunctive Therapy for Diabetes. Br. J. Nutr. 2020, 124, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Gul, R.; Jan, S.U.; Faridullah, S.; Sherani, S.; Jahan, N. Preliminary Phytochemical Screening, Quantitative Analysis of Alkaloids, and Antioxidant Activity of Crude Plant Extracts from Ephedra intermedia Indigenous to Balochistan. Sci. World J. 2017, 2017, 5873648. [Google Scholar] [CrossRef]
- Dal Canto, E.; Ceriello, A.; Rydén, L.; Ferrini, M.; Hansen, T.B.; Schnell, O.; Standl, E.; Beulens, J.W. Diabetes as a Cardiovascular Risk Factor: An Overview of Global Trends of Macro and Micro Vascular Complications. Eur. J. Prev. Cardiol. 2019, 26 (Suppl. S2), 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ansari, P.; Akther, S.; Khan, J.T.; Islam, S.S.; Masud, S.R.; Rahman, A.; Seidel, V.; Abdel-Wahab, Y.H.A. Hyperglycaemia-Linked Diabetic Foot Complications and Their Management Using Conventional and Alternative Therapies. Appl. Sci. 2022, 12, 11777. [Google Scholar] [CrossRef]
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Ferrannini, E.; Holman, R.R.; Sherwin, R.; Zinman, B. Medical Management of Hyperglycemia in Type 2 Diabetes: A Consensus Algorithm for the Initiation and Adjustment of Therapy. Diabetes Care 2009, 32, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Saurabh, V.; Tomar, M.; Hasan, M.; Changan, S.; Sasi, M.; Maheshwari, C.; Prajapati, U.; Singh, S.; Prajapat, R.K.; et al. Mango (Mangifera indica L.) Leaves: Nutritional Composition, Phytochemical Profile, and Health-Promoting Bioactivities. Antioxidants 2021, 10, 299. [Google Scholar] [CrossRef]
- Kulkarni, V.M.; Rathod, V.K. Extraction of Mangiferin from Mangifera indica Leaves Using Three Phase Partitioning Coupled with Ultrasound. Ind. Crop Prod. 2014, 52, 292–297. [Google Scholar] [CrossRef]
- Henquin, J.C. Triggering and Amplifying Pathways of Regulation of Insulin Secretion by Glucose. Diabetes 2000, 49, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Porte, D. Clinical Importance of Insulin Secretion and Its Interaction with Insulin Resistance in the Treatment of Type 2 Diabetes Mellitus and Its Complications. Diabetes Metab. Res. Rev. 2001, 17, 181–188. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Kim, Y.S.; Ryu, S.Y.; Choi, Y.H.; Cha, M.-R.; Yang, H.J.; Park, S. Platyconic Acid, a Saponin from Platycodi Radix, Improves Glucose Homeostasis by Enhancing Insulin Sensitivity In Vitro and In Vivo. Eur. J. Nutr. 2012, 51, 529–540. [Google Scholar] [CrossRef]
- Kumari, D.M.; Jain, S. Tannin: An Antinutrient with Positive Effect to Manage Diabetes. Res. J. Recent Sci. 2012, 2277, 2502. [Google Scholar]
- Muhammad, I.; Rahman, N.; Nayab, G.E.; Nishan, U.; Shah, M. Antidiabetic Activities of Alkaloids Isolated from Medicinal Plants. Braz. J. Pharm. Sci. 2021, 57, e19130. [Google Scholar] [CrossRef]
- Hajiaghaalipour, F.; Khalilpourfarshbafi, M.; Arya, A. Modulation of Glucose Transporter Protein by Dietary Flavonoids in Type 2 Diabetes Mellitus. Int. J. Biol. Sci. 2015, 11, 508–524. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.; Savarino, V.; Paredes-Diaz, A.; Fotopoulos, G. A Review of the Role of Soluble Fiber in Health with Specific Reference to Wheat Dextrin. J. Int. Med. Res. 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Ngo, D.-N.; Vo, T.T.N.; Vo, T.S. Mechanism of Action of Mangifera Indica Leaves for Anti-Diabetic Activity. Sci. Pharm. 2019, 87, 13. [Google Scholar] [CrossRef]
- Irondi, E.A.; Oboh, G.; Akindahunsi, A.A. Antidiabetic Effects of Mangifera Indica Kernel Flour-Supplemented Diet in Streptozotocin-Induced Type 2 Diabetes in Rats. Food Sci. Nutr. 2016, 4, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Gondi, M.; Basha, S.A.; Bhaskar, J.J.; Salimath, P.V.; Rao, U.J.S.P. Anti-Diabetic Effect of Dietary Mango (Mangifera indica L.) Peel in Streptozotocin-Induced Diabetic Rats. J. Sci. Food Agric. 2015, 95, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B., 3rd. Diabetes, Oxidative Stress, and Antioxidants: A Review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Van Den Oever, I.A.M.; Raterman, H.G.; Nurmohamed, M.T.; Simsek, S. Endothelial Dysfunction, Inflammation, and Apoptosis in Diabetes Mellitus. Mediat. Inflamm. 2010, 2010, 792393. [Google Scholar] [CrossRef]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C.-M. Determination of Antioxidants by DPPH Radical Scavenging Activity and Quantitative Phytochemical Analysis of Ficus religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef]
- Minniti, G.; Laurindo, L.F.; Machado, N.M.; Duarte, L.G.; Guiguer, E.L.; Araujo, A.C.; Dias, J.A.; Lamas, C.B.; Nunes, Y.C.; Bechara, M.D.; et al. Mangifera indica L., By-Products, and Mangiferin on Cardio-Metabolic and Other Health Conditions: A Systematic Review. Life 2023, 13, 2270. [Google Scholar] [CrossRef] [PubMed]
- Boas, G.R.V.; Lemos, J.M.R.; de Oliveira, M.W.; dos Santos, R.C.; da Silveira, A.P.S.; Bacha, F.B.; Ito, C.N.A.; Cornelius, E.B.; Lima, F.B.; Rodrigues, A.M.S.; et al. Aqueous Extract from Mangifera indica Linn. (Anacardiaceae) Leaves Exerts Long-Term Hypoglycemic Effect, Increases Insulin Sensitivity and Plasma Insulin Levels on Diabetic Wistar Rats. PLoS ONE 2020, 15, e0227105. [Google Scholar] [CrossRef]
- Anila, L.; Vijayalakshmi, N. Flavonoids from Emblica Officinalis and Mangifera Indica—Effectiveness for Dyslipidemia. J. Ethnopharmacol. 2002, 79, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Aromdee, C.; Pranakhon, R.; Pannangpetch, P. Effects of Iriflophenone 3-C-β-Glucoside on Fasting Blood Glucose Level and Glucose Uptake. Pharmacogn. Mag. 2015, 11, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Moore, W.; Wang, Y.; Luo, J.; McMillan, R.P.; Zhen, W.; Zhou, K.; Liu, D. The Flavonoid Kaempferol Ameliorates Streptozotocin-Induced Diabetes by Suppressing Hepatic Glucose Production. Molecules 2018, 23, 2338. [Google Scholar] [CrossRef]
- Somkuwar, D.; Kamble, V.A. Phytochemical Screening of Ethanolic Extracts of Stem, Leaves, Flower and Seed Kernel of Mangifera indica L. Int. J. Pharma Bio Sci. 2013, 4, P383–P389. [Google Scholar]
- Olasehinde, G.I.; Sholotan, K.J.; Openibo, J.O.; Taiwo, O.S.; Bello, O.A.; Ajayi, J.B.; Ayepola, O.O.; Ajayi, A.A. Phytochemical and Antimicrobial Properties of Mangifera Indica Leaf Extracts. Covenant J. Phys. Life Sci. 2018, 6, 1. [Google Scholar]
- Bbosa, G.S.; Kyegombe, D.B.; Ogwal-Okeng, J.; Bukenya-Ziraba, R.; Odyek, O.; Waako, P. Antibacterial activity of Mangifera indica (L.). Afr. J. Ecol. 2007, 45, 13–16. [Google Scholar] [CrossRef]
- Muthusamy, V.; Anand, S.; Sangeetha, K.; Sujatha, S.; Arun, B.; Lakshmi, B. Tannins Present in Cichorium Intybus Enhance Glucose Uptake and Inhibit Adipogenesis in 3T3-L1 Adipocytes through PTP1B Inhibition. Chem. Interactions 2008, 174, 69–78. [Google Scholar] [CrossRef]
- Li, Y.; Park, J.; Wu, Y.; Cui, J.; Jia, N.; Xi, M.; Wen, A. Identification of AMPK Activator from Twelve Pure Compounds Isolated from Aralia Taibaiensis: Implication in Antihyperglycemic and Hypolipidemic Activities. Korean J. Physiol. Pharmacol. 2017, 21, 279–286. [Google Scholar] [CrossRef]
- Perez, G.R.M.; Zavala, S.M.A.; Perez, G.S.; Perez, G.C. Antidiabetic Effect of Compounds Isolated from Plants. Phytomedicine 1998, 5, 55–75. [Google Scholar] [CrossRef] [PubMed]
- Shigemasa, H. Partial purification of crude gymnemic acids by affinity chromatography and effects of purified fractions on the oral glucose tolerance test in rats. Yonago Igaku Zasshi 1992, 43, 350–364. [Google Scholar]
- Esmaeili, M.A.; Zohari, F.; Sadeghi, H. Antioxidant and Protective Effects of Major Flavonoids from Teucrium polium on β-Cell Destruction in a Model of Streptozotocin-Induced Diabetes. Planta Medica 2009, 75, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- AL-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Concentration (µg/mL) | Ascorbic Acid (% Inhibition) | EEMI (% Inhibition) |
|---|---|---|
| 1.6 | 10.82 ± 1.32 ** | 9.95 ± 1.15 ** |
| 8 | 32.91 ± 1.15 *** | 30.42 ± 1.17 *** |
| 40 | 70.47 ± 1.85 *** | 47.25 ± 2.07 *** |
| 200 | 87.11 ± 1.61 *** | 62.14 ± 2.15 *** |
| 1000 | 95.04 ± 1.55 *** | 73.32 ± 2.25 *** |
| 5000 | 97.24 ± 1.10 *** | 80.99 ± 1.35 *** |
| Days | Treatment Group | Body wt. (gm) | HDL (mg/dL) | LDL (mg/dL) | TG (mg/dL) | Total Cholesterol (mg/dL) |
|---|---|---|---|---|---|---|
| 0 days | Diabetic control | 170.0 ± 0.8 | 35.92 ± 2.8 | 57.26 ± 3.1 | 93.22 ± 2.1 | 92.00 ± 1.2 |
| EEMI (250 mg/kg) | 169.3 ± 2.1 | 31.68 ± 2.7 | 46.83 ± 6.1 | 87.17 ± 6.1 | 89.10 ± 2.7 | |
| EEMI (500 mg/kg) | 164.9 ± 2.3 | 33.17 ± 1.7 | 52.78 ± 4.5 | 79.57 ± 5.7 | 85.97 ± 3.0 | |
| Glibenclamide (0.5 mg/kg) | 158.8 ± 5.2 | 34.05 ± 2.1 | 55.54 ± 3.0 | 85.00 ± 2.9 | 94.77 ± 2.4 | |
| 7 days | Diabetic control | 176.1 ± 3.1 | 29.92 ± 2.8 | 77.55 ± 6.9 | 118.0 ± 6.9 | 83.67 ± 3.9 |
| EEMI (250 mg/kg) | 166.2 ± 2.7 | 34.39 ± 2.9 | 66.69 ± 6.1 | 107.0 ± 6.1 | 79.31 ± 1.5 | |
| EEMI (500 mg/kg) | 167.2 ± 1.0 | 42.10 ± 1.7 | 76.32 ± 2.3 | 105.1 ± 4.5 | 72.29 ± 2.8 | |
| Glibenclamide (0.5 mg/kg) | 166.9 ± 2.0 | 62.81 ± 2.6 *** | 36.99 ± 3.1 ** | 47.55 ± 3.1 ** | 76.51 ± 2.9 | |
| 14 days | Diabetic control | 171.6 ± 0.9 | 33.91 ± 2.7 | 57.57 ± 4.1 | 98.00 ± 4.1 | 88.14 ± 2.2 |
| EEMI (250 mg/kg) | 167.2 ± 1.9 | 33.61 ± 1.7 | 75.94 ± 6.1 | 116.3 ± 6.1 | 72.18 ± 1.0 ** | |
| EEMI (500 mg/kg) | 164.5 ± 3.5 | 36.05 ± 2.9 | 58.61 ± 4.5 | 84.05 ± 4.5 | 63.43 ± 4.2 ** | |
| Glibenclamide (0.5 mg/kg) | 167.8 ± 1.1 | 70.79 ± 2.9 *** | 39.09 ± 3.8 * | 49.65 ± 3.8 *** | 53.72 ± 1.7 *** | |
| 21 days | Diabetic control | 191.4 ± 1.5 | 37.29 ± 3.2 | 82.05 ± 3.4 | 122.5 ± 3.4 | 100.6 ± 2.5 |
| EEMI (250 mg/kg) | 160.8 ± 4.4 ** | 26.78 ± 1.7 | 72.91 ± 6.1 | 113.2 ± 6.1 | 57.45 ± 5.2 ** | |
| EEMI (500 mg/kg) | 154.1 ± 2.3 *** | 41.05 ± 2.0 * | 61.45 ± 4.5 * | 86.89 ± 4.5 ** | 63.12 ± 2.0 *** | |
| Glibenclamide (0.5 mg/kg) | 155.4 ± 2.7 *** | 85.28 ± 2.5 ** | 39.64 ± 0.1 *** | 46.87 ± 3.4 *** | 48.10 ± 1.8 *** | |
| 28 days | Diabetic control | 196.4 ± 2.1 | 23.19 ± 2.5 | 95.88 ± 3.3 | 136.3 ± 3.3 | 105.6 ± 2.7 |
| EEMI (250 mg/kg) | 154.9 ± 2.3 *** | 31.62 ± 1.7 | 59.69 ± 6.1 ** | 100.0 ± 6.1 ** | 60.96 ± 3.8 *** | |
| EEMI (500 mg/kg) | 150.0 ± 3.1 *** | 42.67 ± 1.9 ** | 57.27 ± 4.5 ** | 82.71 ± 4.5 *** | 57.44 ± 3.6 *** | |
| Glibenclamide (0.5 mg/kg) | 153.1 ± 3.0 *** | 92.34 ± 1.9 *** | 44.37 ± 1.0 *** | 41.59 ± 2.8 *** | 41.66 ± 2.3 *** |
| Group | Results |
|---|---|
| Alkaloids | + |
| Tannins | + |
| Saponins | + |
| Steroids | + |
| Glycosides | + |
| Flavonoids | + |
| Reducing sugar | + |
| Anthraquinone | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, J.T.; Richi, A.E.; Riju, S.A.; Jalal, T.; Orchi, R.J.; Singh, S.; Bhagat, P.; Abdel-Wahab, Y.H.A.; Ansari, P. Evaluation of Antidiabetic Potential of Mangifera indica Leaf in Streptozotocin-Induced Type 2 Diabetic Rats: Focus on Glycemic Control and Cholesterol Regulation. Endocrines 2024, 5, 137-152. https://doi.org/10.3390/endocrines5020010
Khan JT, Richi AE, Riju SA, Jalal T, Orchi RJ, Singh S, Bhagat P, Abdel-Wahab YHA, Ansari P. Evaluation of Antidiabetic Potential of Mangifera indica Leaf in Streptozotocin-Induced Type 2 Diabetic Rats: Focus on Glycemic Control and Cholesterol Regulation. Endocrines. 2024; 5(2):137-152. https://doi.org/10.3390/endocrines5020010
Chicago/Turabian StyleKhan, Joyeeta T., Anika E. Richi, Sanjida A. Riju, Tanjila Jalal, Rejwana J. Orchi, Smita Singh, Phulgen Bhagat, Yasser H. A. Abdel-Wahab, and Prawej Ansari. 2024. "Evaluation of Antidiabetic Potential of Mangifera indica Leaf in Streptozotocin-Induced Type 2 Diabetic Rats: Focus on Glycemic Control and Cholesterol Regulation" Endocrines 5, no. 2: 137-152. https://doi.org/10.3390/endocrines5020010