
Serum IL-1ra Is Associated with but Has No Genetic Link to Type 1 Diabetes
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Fluorescent Immuno-Assay Measurement of IL-1ra
2.3. Data Acquisition
2.4. Statistical Analysis
3. Results
3.1. Clinical and Demographic Characteristics of DAISY Cohort
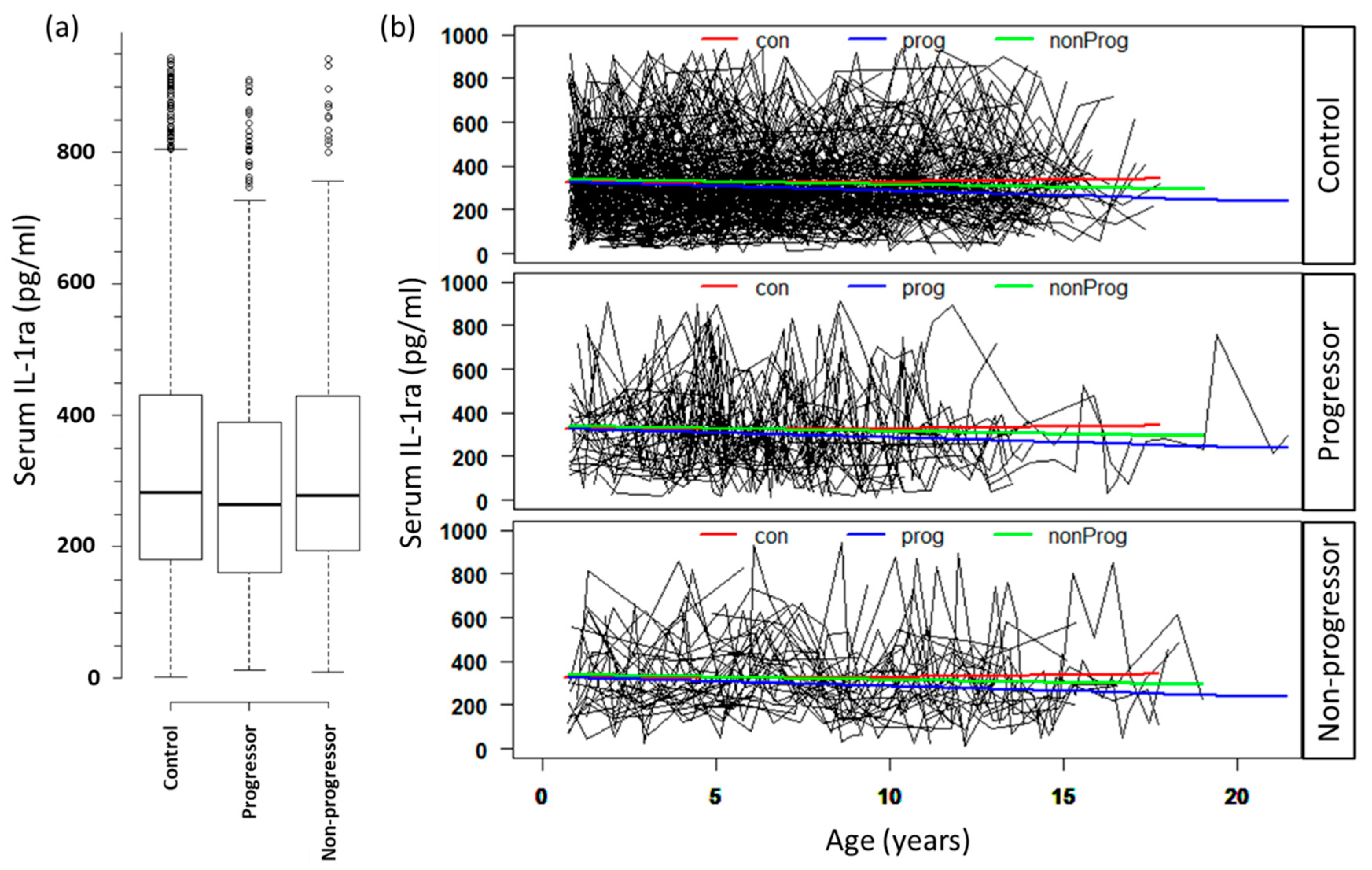
3.2. Serum Levels of IL-1ra Are Associated with Progression to T1D
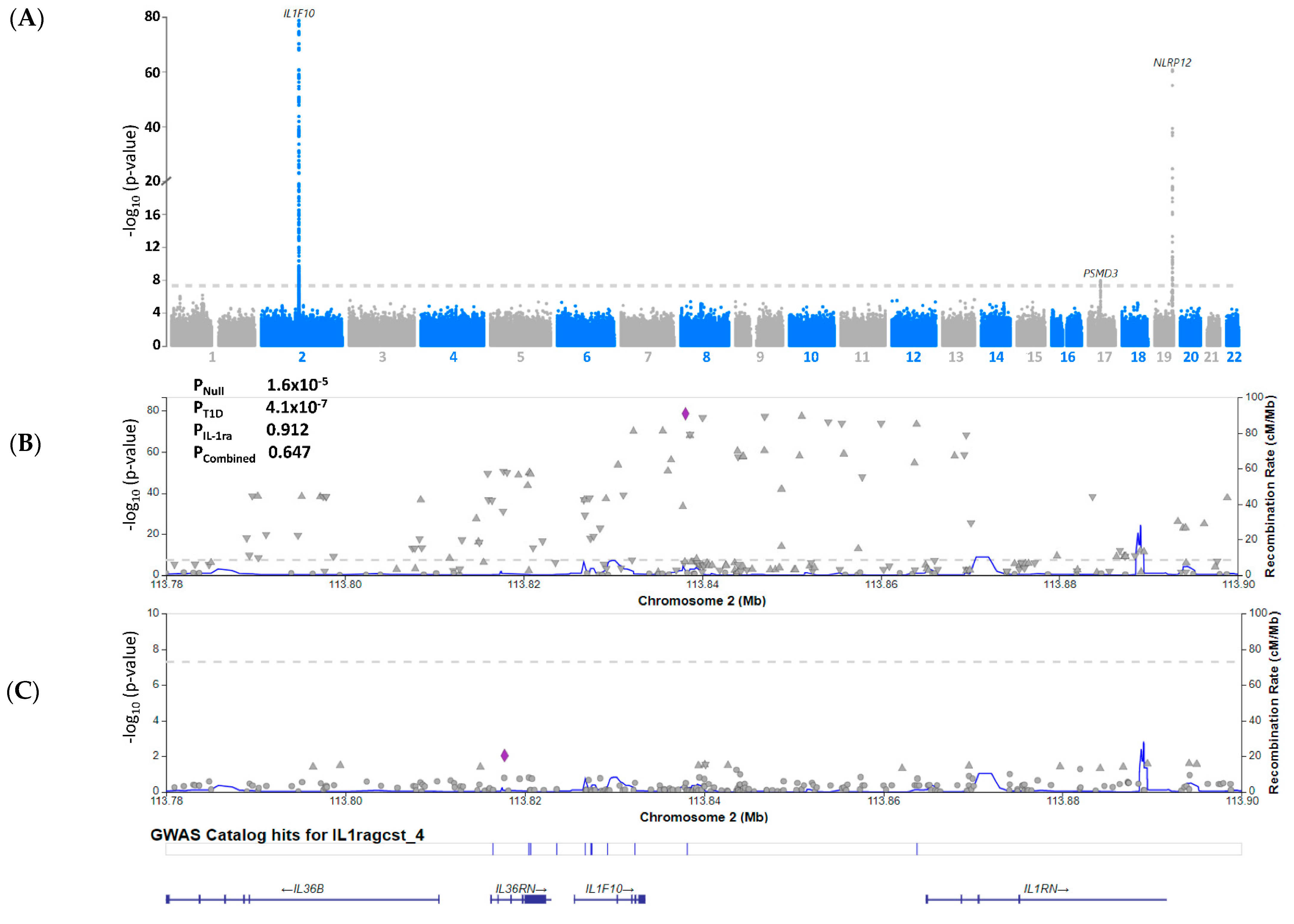
3.3. Serum IL-1ra Levels Are Not Genetically Associated with T1D
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atkinson, M.A.; Eisenbarth, G.S. Type 1 diabetes: New perspectives on disease pathogenesis and treatment. Lancet 2001, 358, 221–229. [Google Scholar] [CrossRef]
- Grieco, F.A.; Vendrame, F.; Spagnuolo, I.; Dotta, F. Innate immunity and the pathogenesis of type 1 diabetes. Semin. Immunopathol. 2011, 33, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Ting, C.; Bansal, V.; Batal, I.; Mounayar, M.; Chabtini, L.; El Akiki, G.; Azzi, J. Impairment of immune systems in diabetes. Adv. Exp. Med. Biol. 2012, 771, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Padgett, L.E.; Broniowska, K.A.; Hansen, P.A.; Corbett, J.A.; Tse, H.M. The role of reactive oxygen species and proinflammatory cytokines in type 1 diabetes pathogenesis. Ann. N. Y. Acad. Sci. 2013, 1281, 16–35. [Google Scholar] [CrossRef]
- Christen, U.; Kimmel, R. Chemokines as Drivers of the Autoimmune Destruction in Type 1 Diabetes: Opportunity for Therapeutic Intervention in Consideration of an Optimal Treatment Schedule. Front. Endocrinol. (Lausanne) 2020, 11, 591083. [Google Scholar] [CrossRef]
- Hanifi-Moghaddam, P.; Kappler, S.; Seissler, J.; Müller-Scholze, S.; Martin, S.; Roep, B.O.; Strassburger, K.; Kolb, H.; Schloot, N.C. Altered chemokine levels in individuals at risk of Type 1 diabetes mellitus. Diabetic Med. 2006, 23, 156–163. [Google Scholar] [CrossRef]
- Cnop, M.; Welsh, N.; Jonas, J.C.; Jörns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54 (Suppl. 2), S97–S107. [Google Scholar] [CrossRef]
- Schloot, N.C.; Hanifi-Moghaddam, P.; Aabenhus-Andersen, N.; Alizadeh, B.Z.; Saha, M.T.; Knip, M.; Devendra, D.; Wilkin, T.; Bonifacio, E.; Roep, B.O.; et al. Association of immune mediators at diagnosis of Type 1 diabetes with later clinical remission. Diabetic Med. 2007, 24, 512–520. [Google Scholar] [CrossRef]
- Pfleger, C.; Mortensen, H.B.; Hansen, L.; Herder, C.; Roep, B.O.; Hoey, H.; Aanstoot, H.J.; Kocova, M.; Schloot, N.C. Association of IL-1ra and adiponectin with C-peptide and remission in patients with type 1 diabetes. Diabetes 2008, 57, 929–937. [Google Scholar] [CrossRef]
- Kaas, A.; Pfleger, C.; Hansen, L.; Buschard, K.; Schloot, N.C.; Roep, B.O.; Mortensen, H.B. Association of adiponectin, interleukin (IL)-1ra, inducible protein 10, IL-6 and number of islet autoantibodies with progression patterns of type 1 diabetes the first year after diagnosis. Clin. Exp. Immunol. 2010, 161, 444–452. [Google Scholar] [CrossRef]
- Purohit, S.; Sharma, A.; Hopkins, D.; Steed, L.; Bode, B.; Anderson, S.W.; Reed, J.C.; Steed, R.D.; Yang, T.; She, J.X. Large-Scale Discovery and Validation Studies Demonstrate Significant Reductions in Circulating Levels of IL8, IL-1Ra, MCP-1, and MIP-1β in Patients With Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2015, 100, E1179–E1187. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.M.; Faulenbach, M.; Vaag, A.; Vølund, A.; Ehses, J.A.; Seifert, B.; Mandrup-Poulsen, T.; Donath, M.Y. Interleukin-1-receptor antagonist in type 2 diabetes mellitus. N. Engl. J. Med. 2007, 356, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.E.; Irawaty, W.; Darwiche, R.; Brodnicki, T.C.; Santamaria, P.; Allison, J.; Kay, T.W. IL-1 receptor deficiency slows progression to diabetes in the NOD mouse. Diabetes 2004, 53, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, J.O.; Eizirik, D.L.; Sandler, S. IL-1 receptor antagonist inhibits recurrence of disease after syngeneic pancreatic islet transplantation to spontaneously diabetic non-obese diabetic (NOD) mice. Clin. Exp. Immunol. 1997, 108, 314–317. [Google Scholar] [CrossRef]
- Ablamunits, V.; Henegariu, O.; Hansen, J.B.; Opare-Addo, L.; Preston-Hurlburt, P.; Santamaria, P.; Mandrup-Poulsen, T.; Herold, K.C. Synergistic reversal of type 1 diabetes in NOD mice with anti-CD3 and interleukin-1 blockade: Evidence of improved immune regulation. Diabetes 2012, 61, 145–154. [Google Scholar] [CrossRef]
- Moran, A.; Bundy, B.; Becker, D.J.; DiMeglio, L.A.; Gitelman, S.E.; Goland, R.; Greenbaum, C.J.; Herold, K.C.; Marks, J.B.; Raskin, P.; et al. Interleukin-1 antagonism in type 1 diabetes of recent onset: Two multicentre, randomised, double-blind, placebo-controlled trials. Lancet 2013, 381, 1905–1915. [Google Scholar] [CrossRef]
- Sanda, S.; Bollyky, J.; Standifer, N.; Nepom, G.; Hamerman, J.A.; Greenbaum, C. Short-term IL-1beta blockade reduces monocyte CD11b integrin expression in an IL-8 dependent fashion in patients with type 1 diabetes. Clin. Immunol. 2010, 136, 170–173. [Google Scholar] [CrossRef]
- Rewers, M.; Norris, J.M.; Eisenbarth, G.S.; Erlich, H.A.; Beaty, B.; Klingensmith, G.; Hoffman, M.; Yu, L.; Bugawan, T.L.; Blair, A.; et al. Beta-cell autoantibodies in infants and toddlers without IDDM relatives: Diabetes autoimmunity study in the young (DAISY). J. Autoimmun. 1996, 9, 405–410. [Google Scholar] [CrossRef]
- Rewers, M.; Bugawan, T.L.; Norris, J.M.; Blair, A.; Beaty, B.; Hoffman, M.; McDuffie, R.S., Jr.; Hamman, R.F.; Klingensmith, G.; Eisenbarth, G.S.; et al. Newborn screening for HLA markers associated with IDDM: Diabetes autoimmunity study in the young (DAISY). Diabetologia 1996, 39, 807–812. [Google Scholar] [CrossRef]
- Norris, J.M.; Yin, X.; Lamb, M.M.; Barriga, K.; Seifert, J.; Hoffman, M.; Orton, H.D.; Barón, A.E.; Clare-Salzler, M.; Chase, H.P.; et al. Omega-3 polyunsaturated fatty acid intake and islet autoimmunity in children at increased risk for type 1 diabetes. Jama 2007, 298, 1420–1428. [Google Scholar] [CrossRef]
- Su, L.F.; Kidd, B.A.; Han, A.; Kotzin, J.J.; Davis, M.M. Virus-specific CD4(+) memory-phenotype T cells are abundant in unexposed adults. Immunity 2013, 38, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Folkersen, L.; Gustafsson, S.; Wang, Q.; Hansen, D.H.; Hedman, Å.K.; Schork, A.; Page, K.; Zhernakova, D.V.; Wu, Y.; Peters, J. Genomic and drug target evaluation of 90 cardiovascular proteins in 30,931 individuals. Nat. Metab. 2020, 2, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Chiou, J.; Geusz, R.J.; Okino, M.L.; Han, J.Y.; Miller, M.; Melton, R.; Beebe, E.; Benaglio, P.; Huang, S.; Korgaonkar, K.; et al. Interpreting type 1 diabetes risk with genetics and single-cell epigenomics. Nature 2021, 594, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Herder, C.; Brunner, E.J.; Rathmann, W.; Strassburger, K.; Tabák, A.G.; Schloot, N.C.; Witte, D.R. Elevated levels of the anti-inflammatory interleukin-1 receptor antagonist precede the onset of type 2 diabetes: The Whitehall II study. Diabetes Care 2009, 32, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Hejblum, B.P.; Simon, N.; Jojic, V.; Dekker, C.L.; Thiébaut, R.; Tibshirani, R.J.; Davis, M.M. Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 869–874. [Google Scholar] [CrossRef]
- Vetrano, M.; Wegman, A.; Koes, B.; Mehta, S.; King, C.A. Serum IL-1RA levels increase from follicular to luteal phase of the ovarian cycle: A pilot study on human female immune responses. PLoS ONE 2020, 15, e0238520. [Google Scholar] [CrossRef]
- Willis, C.; Morris, J.M.; Danis, V.; Gallery, E.D. Cytokine production by peripheral blood monocytes during the normal human ovulatory menstrual cycle. Hum. Reprod. 2003, 18, 1173–1178. [Google Scholar] [CrossRef]
- Cucca, F.; Goy, J.V.; Kawaguchi, Y.; Esposito, L.; Merriman, M.E.; Wilson, A.J.; Cordell, H.J.; Bain, S.C.; Todd, J.A. A male-female bias in type 1 diabetes and linkage to chromosome Xp in MHC HLA-DR3-positive patients. Nat. Genet. 1998, 19, 301–302. [Google Scholar] [CrossRef]
- Sakiani, S.; Olsen, N.J.; Kovacs, W.J. Gonadal steroids and humoral immunity. Nat. Rev. Endocrinol. 2013, 9, 56–62. [Google Scholar] [CrossRef]
- Cabrera, S.M.; Wang, X.; Chen, Y.G.; Jia, S.; Kaldunski, M.L.; Greenbaum, C.J.; Mandrup-Poulsen, T.; Hessner, M.J. Interleukin-1 antagonism moderates the inflammatory state associated with Type 1 diabetes during clinical trials conducted at disease onset. Eur. J. Immunol. 2016, 46, 1030–1046. [Google Scholar] [CrossRef] [Green Version]
- Morran, M.P.; Vonberg, A.; Khadra, A.; Pietropaolo, M. Immunogenetics of type 1 diabetes mellitus. Mol. Asp. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Sumpter, K.M.; Adhikari, S.; Grishman, E.K.; White, P.C. Preliminary studies related to anti-interleukin-1β therapy in children with newly diagnosed type 1 diabetes. Pediatr. Diabetes 2011, 12, 656–667. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Subject Characteristics | Control (n = 222) | Non-Progressor (n = 41) | Progressor (n = 49) | p Value | |
|---|---|---|---|---|---|
| Age of first sample, median (IQR) | 1.26 (0.78–2.58) | 1.29 (0.85–3.09) | 1.28 (0.80–2.73) | 0.7 | |
| HLA genotype, n (%) | DR3/3 or DR3/X or DRX/X | 90 (40.54) | 13 (31.71) | 12 (24.49) | 0.08 |
| DR3/4 or DR4/4 or DR4/X | 132 (59.46) | 28 (68.29) | 37 (75.51) | ||
| FDR status, n (%) | GP | 97 (43.69) | 16 (39.02) | 17 (34.69) | 0.48 |
| FDR | 125 (56.31) | 25 (60.98) | 32 (65.31) | ||
| Gender, n (%) | Female | 97 (43.69) | 22 (53.66) | 24 (48.98) | 0.45 |
| Male | 125 (56.31) | 19 (46.34) | 25 (51.02) | ||
| Effect | Estimate | Standard Error | p-Value | |
|---|---|---|---|---|
| Intercept | 16.18 | 0.35 | <0.0001 | |
| Age | 0.01 | 0.03 | 0.64 | |
| Gender | Female vs. Male | 0.72 | 0.26 | 0.005 |
| HLA | DR3/4 or DR4/4 or DR4/X vs. DR3/3 or DR3/X or DRX/X | 0.09 | 0.28 | 0.75 |
| First-degree relative | YES vs. NO | 0.88 | 0.27 | 0.001 |
| Progression to T1D | Non-progressor vs. Control | −0.2 | 0.36 | 0.58 |
| Progressor vs. Control | −0.74 | 0.33 | 0.023 | |
| Progressor vs. Non-progressor | −0.54 | 0.43 | 0.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, P.M.H.; Dong, F.; Bin Satter, K.; Richardson, K.P.; Patel, R.; Tran, L.K.H.; Hopkins, D.; Kolhe, R.; Waugh, K.; Rewers, M.; et al. Serum IL-1ra Is Associated with but Has No Genetic Link to Type 1 Diabetes. Endocrines 2022, 3, 570-577. https://doi.org/10.3390/endocrines3030048
Tran PMH, Dong F, Bin Satter K, Richardson KP, Patel R, Tran LKH, Hopkins D, Kolhe R, Waugh K, Rewers M, et al. Serum IL-1ra Is Associated with but Has No Genetic Link to Type 1 Diabetes. Endocrines. 2022; 3(3):570-577. https://doi.org/10.3390/endocrines3030048
Chicago/Turabian StyleTran, Paul M. H., Fran Dong, Khaled Bin Satter, Katherine P. Richardson, Roshni Patel, Lynn K. H. Tran, Diane Hopkins, Ravindra Kolhe, Kathleen Waugh, Marian Rewers, and et al. 2022. "Serum IL-1ra Is Associated with but Has No Genetic Link to Type 1 Diabetes" Endocrines 3, no. 3: 570-577. https://doi.org/10.3390/endocrines3030048