
Divergent Gene Expression Profiles in Alaskan Sea Otters: An Indicator of Chronic Domoic Acid Exposure?

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reference Sea Otters
2.2. Kachemak Bay Otters
2.3. Blood Collection and RNA Extraction
2.4. cDNA Synthesis
2.5. Real-Time PCR
2.6. Analysis of Urine and Feces for the Presence of Domoic Acid
2.7. Statistical Analysis
3. Results
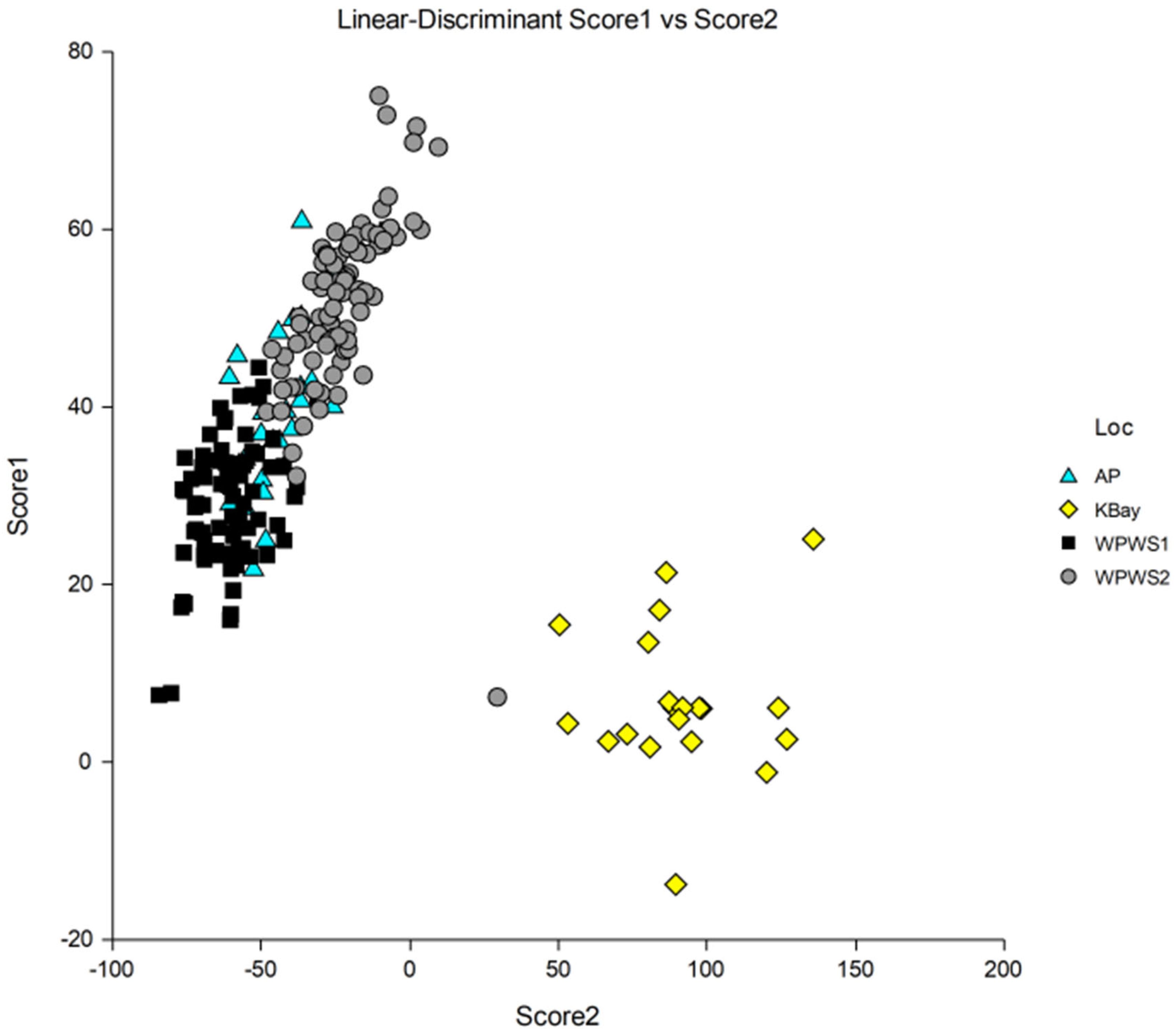
3.1. Gene Expression Analyses
3.2. Analysis of Urine and Feces for the Presence of Domoic Acid
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NCCOS (National Centers for Coastal Ocean Science). State of Kachemak Bay Report Highlights Long-Term Monitoring Data—NCCOS Coastal Science Website (noaa.gov); State of Kachemak Bay; NOAA Kasitsna Bay Laboratory: Homer, AK, USA, 2021. [Google Scholar]
- Wilcox, B.A.; Aguirre, A.A. One ocean, one health. EcoHealth 2004, 1, 211–212. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwacke, L.H.; Gulland, F.M.; White, S. Sentinel species in oceans and human health. In Environmental Toxicology; Springer: New York, NY, USA, 2013; pp. 503–528. [Google Scholar] [CrossRef]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Burge, C.A.; Hershberger, P.K.; Behringer, D.C.; Silliman, B.R.; Lafferty, K.D. Climate change can drive marine diseases. In Marine Disease Ecology; Behringer, D.C., Silliman, B.R., Lafferty, K.D., Eds.; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Landsberg, J.H.; Lefebvre, K.A.; Flewelling, L.J. Effects of Toxic Microalgae on Marine Organisms. In Toxins and Biologically Active Compounds from Microalgae, 1st ed.; Rossini, G.P., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 379–449. [Google Scholar]
- Jessup, D.A.; Miller, M.; Kreuder-Johnson, C. Sea otters in a dirty ocean. J. Am. Vet. Med. Assoc. 2007, 231, 1648–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a Novel Marine Harmful Algal Bloom: Cyanotoxin (Microcystin) Transfer from Land to Sea Otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef]
- Oates, S.C.; Miller, M.A.; Byrne, B.A.; Chouicha, N.; Hardin, D.; Jessup, D.; Dominik, C.; Roug, A.; Schriewer, A.; Jang, S.S.; et al. Epidemiology and potential land-sea transfer of enteric bacteria from terrestrial to marine species in the Monterey Bay Region of California. J. Wildl. Dis. 2012, 48, 654–668. [Google Scholar] [CrossRef]
- Shapiro, K.; Silver, M.W.; Largier, J.L.; Conrad, P.A.; Mazet, J.A.K. Association of Toxoplasma gondii oocysts with fresh, estuarine, and marine macroaggregates. Limnol. Oceanogr. 2012, 57, 449–456. [Google Scholar] [CrossRef]
- Garlich-Miller, J.L.; Esslinger, G.G.; Weitzman, B.P. Aerial Surveys of Sea Otters (Enhydra lutris) in Lower Cook Inlet; Alaska. U.S. Fish and Wildlife Technical Report MMM 2018-01; U.S. Fish and Wildlife Service: Anchorage, AK, USA, 2018. [Google Scholar]
- Gill, V.A. Marine Mammal Unusual Mortality Event Initiation Protocol for Northern Sea Otters; U.S. Fish and Wildlife Service Report; U.S. Fish and Wildlife Service: Anchorage, AK, USA, 2006. [Google Scholar]
- Huntington, K.A.B.; Gill, V.A.; Berrian, A.M.; Goldstein, T.; Tuomi, P.; Byrne, B.A.; Worman, K.; Mazet, J. Causes of Mortality of Northern Sea Otters (Enhydra lutris kenyoni) in Alaska From 2002 to 2012. Front. Mar. Sci. 2021, 8, 105. [Google Scholar] [CrossRef]
- Banuet-Martínez, M.; Espinosa-de Aquino, W.; Elorriaga-Verplancken, F.R.; Flores-Morán, A.; García, O.P.; Camacho, M.; Acevedo-Whitehouse, K. Climatic anomaly affects the immune competence of California sea lions. PLoS ONE 2017, 12, e0179359. [Google Scholar] [CrossRef] [Green Version]
- Neves, R.A.; Nascimento, S.M.; Santos, L.N. Harmful algal blooms and shellfish in the marine environment: An overview of the main molluscan responses, toxin dynamics, and risks for human health. Environ. Sci. Pollut. Res. 2021, 28, 55846–55868. [Google Scholar] [CrossRef]
- Berdalet, E.; Fleming, L.E.; Gowen, R.; Davidson, K.; Hess, P.; Backer, L.C.; Moore, S.K.; Hoagland, P.; Enevoldsen, H. Marine harmful algal blooms, human health and wellbeing: Challenges and opportunities in the 21st century. J. Mar. Biol. Assoc. 2016, 96, 61–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine harmful algal blooms (HABs) in the United States: History, current status and future trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef]
- Suryan, R.M.; Arimitsu, M.L.; Coletti, H.A.; Hopcraft, R.R.; Lindeberg, M.R.; Barbeaux, S.J.; Batten, S.D.; Burt, W.J.; Bishop, M.A.; Bodkin, J.L.; et al. Ecosystem response persists after a prolonged marine heatwave. Sci. Rep. 2021, 11, 6235. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, K.A.; Frame, E.R.; Gulland, F.; Hansen, J.D.; Kendrick, P.S.; Beyer, R.P.; Bammler, T.K.; Farin, F.M.; Hiolski, E.M.; Smith, D.R.; et al. A novel antibody-based biomarker for chronic algal toxin exposure and sub-acute neurotoxicity. PLoS ONE 2012, 7, e36213. [Google Scholar] [CrossRef] [Green Version]
- Moriarty, M.E.; Tinker, M.T.; Miller, M.A.; Tomoleoni, J.A.; Staedler, M.M.; Fujii, J.A.; Batac, F.I.; Dodd, E.M.; Kudela, R.M.; Zubkousky-White, V.; et al. Exposure to domoic acid is an ecological driver of cardiac disease in southern sea otters. Harmful Algae 2021, 101, 101973. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Whitehouse, K.; Duffus, A.L.J. Effects of Environmental Change on Wildlife Health. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3429–3438. [Google Scholar] [CrossRef] [Green Version]
- Hiolski, E.M.; Kendrick, P.S.; Frame, E.R.; Myers, M.S.; Bammler, T.K.; Beyer, R.P.; Farin, F.M.; Wilkerson, H.W.; Smith, D.R.; Marcinek, D.J.; et al. Chronic low-level domoic acid exposure alters gene transcription and impairs mitochondrial function in the CNS. Aquat. Toxicol. 2014, 155, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Tilton, S.C.; Bammler, T.K.; Beyer, R.P.; Srinouanprachan, S.; Stapleton, P.L.; Farin, F.M.; Gallagher, E.P. Gene expression profiles in zebrafish brain after acute exposure to domoic acid at symptomatic and asymptomatic doses. Toxicol. Sci. 2009, 107, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.C.; Morey, J.S.; Ramsdell, J.S.; van Dolah, F.M. Acute phase gene expression in mice exposed to the marine neurotoxin domoic acid. Neuroscience 2005, 136, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Blanchong, J.A.; Robinson, S.J.; Samuel, M.D.; Foster, J.T. Application of genetics and genomics to wildlife epidemiology. J. Wildl. Manag. 2016, 80, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Carugati, L.; Berzano, M.; Cahill, A.E.; Carvalho, S.; Chenuil, A.; Corinaldesi, C.; Cristina, S.; David, R.; Dell’Anno, A.; et al. Implementing and innovating marine monitoring approaches for assessing marine environmental status. Front. Mar. Sci. 2016, 3, 213. [Google Scholar] [CrossRef]
- McLoughlin, K.; Turteltaub, K.; Bankaitis-Davis, D.; Gerren, R.; Siconolfi, L.; Storm, K.; Cheronis, J.; Trollinger, D.; Macejak, D.; Tryon, V.; et al. Limited Dynamic Range of Immune Response Gene Expression Observed in Healthy Blood Donors Using RT-PCR. Mol. Med. 2006, 12, 185–195. [Google Scholar] [CrossRef]
- Bowen, L.; Miles, A.K.; Ballachey, B.; Waters, S.; Bodkin, J. Gene Transcript Profiling in Sea Otters Post-Exxon Valdez Oil Spill: A Tool for Marine Ecosystem Health Assessment. J. Mar. Sci. Technol. 2016, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Tinker, M.T.; Bodkin, J.L.; Bowen, L.; Ballachey, B.; Bentall, G.; Burdin, A.; Coletti, H.; Esslinger, G.; Hatfield, B.B.; Kenner, M.C.; et al. Sea otter population collapse in southwest Alaska: Assessing ecological covariates, consequences, and causal factors. Ecol. Monogr. 2021, 91, e01472. [Google Scholar] [CrossRef]
- Bowen, L.; Miles, A.K.; Murray, M.; Haulena, M.; Tuttle, J.; Van Bonn, W.; Adams, L.; Bodkin, J.L.; Ballachey, B.E.; Estes, J.A.; et al. Gene transcription in sea otters (Enhydra lutris); emerging diagnostics in marine mammal and ecosystem health. Mol. Ecol. Resour. 2012, 12, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Miles, A.K.; Bowen, L.; Ballachey, B.; Bodkin, J.; Murray, M.; Estes, J.L.; Keister, R.A.; Stott, J.L. Variations of transcript profiles between sea otters (Enhydra lutris) from Prince William Sound, Alaska, and clinically normal reference otters. Mar. Ecol. Prog. Ser. 2012, 451, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Bowen, L.; Miles, A.K.; Waters, S.; Gustine, D.; Joly, K.; Hilderbrand, G. Using gene transcription to assess ecological and anthropogenic stressors in brown bears. EcoHealth 2017, 15, 121–131. [Google Scholar] [CrossRef]
- Murray, M.J. Chapter 7—Veterinary medicine and sea otter conservation. In Sea Otter Conservation; Larson, S.E., Bodkin, J.L., Van Blaricom, G.R., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2015; pp. 159–195. [Google Scholar] [CrossRef]
- Bommer, U.; Thiele, B. The translationally controlled tumor protein (TCTP). Int. J. Biochem. Cell B 2004, 36, 379–385. [Google Scholar] [CrossRef]
- Tuynder, M.; Fiucci, G.; Prieur, S.; Lespagnol, A.; Géant, A.; Beaucourt, S.; Duflaut, D.; Besse, S.; Susini, L.; Cavarelli, J.; et al. Translationally controlled tumor protein is a target of tumor reversion. Proc. Natl. Acad. Sci. USA 2004, 101, 15364–15369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Geng, Y.; Xu, W.; Wu, Y.; He, F.; Shu, W.; Huang, M.; Du, H.; Li, M. The role of translationally controlled tumor protein in tumor growth and metastasis of colon adenocarcinoma cells. J. Proteome Res. 2010, 9, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Bowen, L.; Schwartz, J.; Aldridge, B.; Riva, F.; Miles, A.; Stott, J.L. Differential gene expression induced by exposure of captive mink to fuel oil: A model for the sea otter. EcoHealth 2007, 4, 298–309. [Google Scholar] [CrossRef]
- Raisuddin, S.; Kwok, K.W.H.; Leung, K.M.Y.; Schlenk, D.; Lee, J. The copepod Tigriopus: A promising marine model organism for ecotoxicology and environmental genomics. Aquat. Toxicol. 2007, 83, 161–173. [Google Scholar] [CrossRef]
- Zheng, S.; Song, Y.; Qiu, X.; Sun, T.; Ackland, M.L.; Zhang, W. Annetocin and TCTP expressions in the earthworm Eisenia fetida exposed to PAHs in artificial soil. Ecotox. Environ. Safe 2008, 71, 566–573. [Google Scholar] [CrossRef]
- Goldsby, R.; Kindt, T.; Osborne, B.; Kuby, J. Immunology, 5th ed.; WH Freeman and Company: New York, NY, USA, 2003. [Google Scholar]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Jenne, D.E.; Tschopp, J. Molecular structure and functional characterization of a human complement cytolysis inhibitor found in blood and seminal plasma: Identity to sulfated glycoprotein 2, a constituent of rat testis fluid. Proc. Natl. Acad. Sci. USA 1989, 86, 7123–7127. [Google Scholar] [CrossRef] [Green Version]
- Portnoy, D.S.; Fields, A.T.; Greer, J.B.; Schlenk, D. Genetics and Oil: Transcriptomics, Epigenetics, and Population Genomics as Tools to Understand Animal Responses to Exposure Across Different Time Scales. In Deep Oil Spills—Facts, Fate, and Effects; Murawski, A., Ainsworth, C.H., Gilbert, S., Hollander, D.J., Paris, C.B., Schlüter, M., Wetzel, D.L., Eds.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Ehrlenbach, S.; Rosales, A.; Posch, W.; Wilflingseder, D.; Hermann, M.; Brockmeyer, J.; Karch, H.; Satchell, S.C.; Würzner, R.; Orth-Höller, D. Shiga toxin 2 reduces complement inhibitor CD59 expression on human renal tubular epithelial and glomerular endothelial cells. Infect. Immun. 2013, 81, 2678–2685. [Google Scholar] [CrossRef] [Green Version]
- Oksjoki, R.; Kovanen, P.T.; Meri, S.; Pentikainen, M.O. Function and regulation of the complement system in cardiovascular diseases. Front. Biosci. 2007, 12, 4696–4708. [Google Scholar] [CrossRef] [Green Version]
- Oesch-Bartlomowicz, B.; Oesch, F. Phosphorylation of cytochromes P450: First discovery of a posttranslational modification of a drug-metabolizing enzyme. Biochem. Biophys. Res. Commun. 2005, 338, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Quintana, F.J.; Basso, A.S.; Iglesias, A.H.; Korn, T.; Farez, M.F.; Bettelli, E.; Caccamo, M.; Oukka, M.; Weiner, H.L. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature 2008, 453, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.C.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liang, X.F.; Zhang, W.B.; Mai, K.S.; Huang, Y.; Shen, D. Amnesic shellfish poisoning toxin stimulates the transcription of CYP1A possibly through AHR and ARNT in the liver of red sea bream Pagrus major. Mar. Pollut. Bull 2009, 58, 1643–1648. [Google Scholar] [CrossRef]
- Incardona, J.P. Molecular Mechanisms of Crude Oil Developmental Toxicity in Fish. Arch. Environ. Contam. Toxicol. 2017, 73, 19–32. [Google Scholar] [CrossRef]
- Tabuchi, M.; Veldhoen, N.; Dangerfield, N.; Jeffries, S.; Helbing, C.C.; Ross, P.S. PCB related alteration of thyroid hormones and thyroid hormone receptor gene expression in free-ranging harbor seals (Phoca vitulina). Environ. Health Perspect. 2006, 114, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.J.; O’Malley, B.W. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu. Rev. Biochem. 1994, 63, 451–486. [Google Scholar] [CrossRef] [PubMed]
- Iwama, G.K.; Vijayan, M.M.; Forsyth, R.B.; Ackerman, P.A. Heat shock proteins and physiological stress in fish. Am. Zool. 1999, 39, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Tsan, M.; Gao, B. Cytokine function of heat shock proteins. Am. J. Physiol. Cell Physiol. 2004, 286, C739–C744. [Google Scholar] [CrossRef]
- De Maio, A. Heat shock proteins: Facts, thoughts and dreams. Shock 1999, 11, 1–12. [Google Scholar] [CrossRef]
- Tanabe, S.; Iwat, H.; Tatsukawa, R. Global contamination by persistent organochlorines and their ecotoxicological impact on marine mammals. Sci. Total Environ. 1994, 154, 163–177. [Google Scholar] [CrossRef]
- Krumm, B.; Meng, X.; Li, Y.; Xiang, Y.; Deng, J. Structural basis for antagonism of human interleukin 18 by poxvirus interleukin-18 binding protein. Proc. Natl. Acad. Sci. USA 2008, 105, 20711–20715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beineke, A.; Siebert, U.; Muller, G.; Baumgartner, W. Increased blood interleukin-10 mRNA levels in diseased free-ranging harbor porpoises (Phocoena phocoena). Vet. Immunol. Immunopathol. 2007, 115, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Rigopoulou, E.; Abbott, W.; Haigh, P.; Naoumov, N. Blocking of interleukin-10 receptor—A novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Bowen, L.; Aldridge, B.; Beckmen, K.; Gelatt, T.; Rea, L.; Burek, K.; Pitcher, K.; Stott, J. Differential expression of immune response genes in Steller sea lions (Eumetopias jubatus): An indicator of ecosystem health? EcoHealth 2006, 3, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Wegner, K.M.; Kalbe, M.; Rauch, G.; Kurtz, J.; Schaschl, H.; Reusch, T.B.H. Genetic variation in MHC class II expression and interactions with MHC sequence polymorphism in three-spined sticklebacks. Mol. Ecol. 2006, 15, 1153–1164. [Google Scholar] [CrossRef]
- Dong, L.; Ma, Q.; Whitlock, J.P., Jr. Down-regulation of major histocompatibility complex Q1b gene expression by 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 1997, 272, 29614–29619. [Google Scholar] [CrossRef] [Green Version]
- Tumpey, T.M.; Szretter, K.J.; Van Hoeven, N.; Katz, J.M.; Kochs, G.; Haller, O.; García-Sastre, A.; Staeheli, P. The Mx1 gene protects mice against the pandemic 1918 and highly lethal human H5N1 influenza viruses. J. Virol. 2007, 81, 10818–10821. [Google Scholar] [CrossRef] [Green Version]
- Kibenge, M.J.T.; Munir, K.; Kibenge, F.S.B. Constitutive expression of Atlantic salmon Mx1 protein in CHSE-214 cells confers resistance to Infectious Salmon Anaemia virus. Virol. J. 2005, 2, 75. [Google Scholar] [CrossRef] [Green Version]
- Gurish, M.F.; Humbles, A.; Tao, H.; Finkelstein, S.; Boyce, J.A.; Gerard, C.; Friend, D.S.; Austen, K.F. CCR3 is required for tissue eosinophilia and larval cytotoxicity after infection with Trichinella spiralis. J. Immunol. 2002, 168, 5730–5736. [Google Scholar] [CrossRef] [Green Version]
- Kringel, H.; Iburg, T.; Dawson, H.; Aasted, B.; Roepstorff, A. A time course study of immunological responses in Trichuris suis infected pigs demonstrates induction of a local type 2 response associated with worm burden. Int. J. Parasitol. 2006, 36, 915–924. [Google Scholar] [CrossRef]
- Jennings, K.A.; Loder, M.K.; Sheward, W.J.; Pei, Q.; Deacon, R.M.J.; Benson, M.A.; Olverman, H.J.; Hastie, N.D.; Harmar, A.J.; Shen, S.; et al. Increased expression of the 5-HT transporter confers a low-anxiety phenotype linked to decreased 5-HT transmission. J. Neurosci. 2006, 30, 8955–8964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squire, L.; Berg, D.; Bloom, F.E.; du Lac, S.; Ghosh, A.; Spitzer, N.C. Fundamental Neuroscience; Elsevier Academic Press: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Pazos, A.J.; Ventoso, P.; Martínez-Escauriaza, R.; Pérez-Parallé, M.L.; Blanco, J.; Triviño, J.C.; Sánchez, J.L. Transcriptional response after exposure to domoic acid-producing Pseudo-nitzschia in the digestive gland of the mussel Mytilus galloprovincialis. Toxicon 2017, 140, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Pulido, O.M. Domoic acid toxicologic pathology: A review. Mar. Drugs 2008, 6, 180–219. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, R.; Liu, Z.P.; Li, M.; Aihara, K. Detecting early-warning signasls for suddent deterioration of complex diseases by dynamical network biomarkers. Sci. Rep. 2012, 2, 342. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xie, L.; Zhang, C.; Zhang, Y.; Gu, M. Cloning and expression of a pivotal calcium metabolism regulator: Calmodulin involved in shell formation from pearl oyster (Pinctada fucata). Comp. Biochem. Physiol. B, Biochem. 2004, 138, 235–243. [Google Scholar] [CrossRef]
- Plested, A.J.; Mayer, M.L. Structure and mechanism of kainate receptor modulation by anions. Neuron 2007, 53, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Berman, F.W.; LePage, K.T.; Murray, T.F. Domoic acid neurotoxicity in cultured cerebellar granule neurons is controlled preferentially by the NMDA receptor Ca2+ influx pathway. Brain Res. 2002, 924, 20–29. [Google Scholar] [CrossRef]
- Choi, D.W. Excitotoxic cell death. J. Neurobiol. 1992, 23, 1261–1276. [Google Scholar] [CrossRef]
- Choi, D.W. Calcium and excitotoxic neuronal injury. Ann. N. Y. Acad. Sci. 1994, 747, 162–171. [Google Scholar] [CrossRef]
- Bloodgood, B.L.; Sabatini, B.L. Nonlinear Regulation of Unitary Synaptic Signals by CaV2.3 Voltage-Sensitive Calcium Channels Located in Dendritic Spines. Neuron 2007, 53, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalbuyeva, N.; Brustovetsky, T.; Bolshakov, A.; Brustovetsky, N. Calcium-dependent spontaneously reversible remodeling of brain mitochondria. J. Biol. Chem. 2006, 281, 37547–37558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 2016, 55, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coletti, H.A.; Bodkin, J.L.; Monson, D.H.; Ballachey, B.E.; Dean, T.A. Detecting and inferring cause of change in an Alaska nearshore marine ecosystem. Ecosphere 2016, 7, e01489. [Google Scholar] [CrossRef]
- Saucillo, D.C.; Gerriets, V.A.; Sheng, J.; Rathmell, J.C.; MacIver, N.J. Leptin metabolically licenses T cells for activation to link nutrition and immunity. J. Immunol. 2014, 192, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Monson, D.H.; Doak, D.F.; Ballachey, B.E.; Bodkin, J.L. Could residual oil from the Exxon Valdez spill create a long-term population “sink” for sea otters in Alaska? Ecol. Appl. 2011, 21, 2917–2932. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Powell, C.L.; Busman, M.; Doucette, G.J.; Moeller, P.D.R.; Silver, J.B.; Miller, P.E.; Hughes, M.P.; Singaram, S.; Silver, M.W.; et al. Detection of domoic acid in northern anchovies and california sea lions associated with an unusual mortality event. Nat. Toxins 1999, 7, 85–92. [Google Scholar] [CrossRef]
- Bargu, S.; Silver, M.; Goldstein, T.; Roberts, K.; Gulland, F. Complexity of domoic acid-related sea lion strandings in Monterey Bay, California: Foraging patterns, climate events, and toxic blooms. Mar. Ecol. Prog. Ser. 2010, 418, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Beltrán, A.P.; Palafox-Uribe, M.; Grajales-Montiel, J.; Cruz-Villacorta, A.; Ochoa, J.L. Sea bird mortality at Cabo San Lucas, Mexico: Evidence that toxic diatom blooms are spreading. Toxicon 1997, 35, 447–453. [Google Scholar] [CrossRef]
- Work, T.M.; Barr, B.; Beale, A.M.; Fritz, L.; Quilliam, M.A.; Wright, J.L.C. Epidemiology of domoic acid poisoning in brown pelicans (Pelicanus occidentalis) and Brandt’s cormorants (Phalacrocorax penicillatus) in California. J. Zoo Wildl. Med. 1993, 24, 54–62. [Google Scholar]
- Gibble, C.M.; Kudela, R.M.; Knowles, S.; Bodenstein, B.; Lefebvre, K.A. Domoic acid and saxitoxin in seabirds in the United States between 2007 and 2018. Harmful Algae 2021, 103, 101981. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, J.S.; Zatorre, R.J.; Carpenter, S.; Gendron, D.; Evans, A.C.; Gjedde, A.; Cashman, N.R. Neurologic sequelae of domoic acid intoxication due to the ingestion of contaminated mussels. N. Engl. J. Med. 1990, 322, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, K.A.; Kendrick, P.S.; Ladiges, W.C.; Hiolski, E.M.; Ferriss, B.E.; Smith, D.R.; Marcinek, D.J. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 2017, 64, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, T.; Mazet, J.A.K.; Zabka, T.S.; Langlois, G.; Colegrove, K.M.; Silver, M.; Bargu, S.; Van Dolah, F.; Leighfield, T.; Conrad, P.A.; et al. Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): An increasing risk to marine mammal health. Philos. Trans. R. Soc. B Biol. Sci 2008, 275, 267–276. [Google Scholar] [CrossRef] [Green Version]
- McKibben, S.M.; Peterson, W.; Wood, A.M.; Trainer, V.L.; Hunter, M.; White, A.E. Climatic regulation of the neurotoxin domoic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabka, T.S.; Goldstein, T.; Cross, C.; Mueller, R.W.; Kreuder-Johnson, C.; Gill, S.; Gulland, F.M.D. Characterization of a Degenerative Cardiomyopathy Associated with Domoic Acid Toxicity in California Sea Lions (Zalophus californianus). Vet. Pathol. 2009, 46, 105–119. [Google Scholar] [CrossRef]
- Washburn, B.S.; Rein, K.S.; Baden, D.G.; Walsh, P.J.; Hinton, D.E.; Tullis, K.; Denison, M.S. Brevetoxin-6 (PbTx-6), a Nonaromatic Marine Neurotoxin, Is a Ligand of the Aryl Hydrocarbon Receptor. Arch. Biochem. Biophys. 1997, 343, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.; Pulido, O. Distribution of Glutamate Receptors in Peripheral Tissues. In Glutamate Receptors in Peripheral Tissues; Gill, S., Pulido, O., Eds.; Kluwer Academic Plenum Press: New York, NY, USA, 2005; pp. 3–26, ISBN 0306479737/9780306479731. [Google Scholar]
- Gill, S.S.; Mueller, R.W.; McGuire, P.F.; Pulido, O.M. Potential target sites in peripheral tissues for excitatory neurotransmission and excitotoxicity. Toxicol. Pathol. 2000, 28, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Pulido, O.M. Glutamate receptors in peripheral tissues: Current knowledge, future research, and implications for toxicology. Toxicol. Pathol. 2001, 29, 208–223. [Google Scholar] [CrossRef]
- Gill, S.; Veinot, J.; Kavanagh, M.; Pulido, O. Human heart glutamate receptors—Implications for toxicology, food safety, and drug discovery. Toxicol. Pathol. 2007, 35, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Gulland, F.M.; Haulena, M.; Fauquier, D.; Langlois, G.; Lander, M.E.; Zabka, T.; Duerr, R. Domoic acid toxicity in Californian sea lions (Zalophus californianus): Clinical signs, treatment and survival. Vet. Rec. 2002, 150, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Kreuder, C.; Miller, M.A.; Lowenstine, L.J.; Conrad, P.A.; Carpenter, T.E.; Jessup, D.A.; Mazet, J.A. Evaluation of cardiac lesions and risk factors associated with myocarditis and dilated cardiomyopathy in southern sea otters (Enhydra lutris nereis). Am. J. Vet. Res. 2005, 66, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Incardona, J.P.; Linbo, T.L.; Scholz, N.L. Cardiac toxicity of 5-ring polycyclic aromatic hydrocarbons is differentially dependent on the aryl hydrocarbon receptor 2 isoform during zebrafish development. Toxicol. Appl. Pharm. 2011, 257, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, M.; Duran, R.; Arufe, M.C. Effect of excitatory amino acids on serum TSH and thyroid hormone levels in freely moving rats. Horm. Res. Paediatr. 2000, 54, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Arufe, M.C.; Arias, B.; Duran, R.; Alfonso, M. Effects of domoic acid on serum levels of TSH and thyroid hormones. Endocr. Res. 1995, 21, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Danielson, S.L.; Hennon, T.D.; Monson, D.H.; Suryan, R.; Holderied, K.; Baird, S.; Holderied, K.; Weingartner, T.J. Thermal variability and the importance of in situ temperature observations across the northern Gulf of Alaska. In The Pacific Marine Heatwave: Monitoring During a Major Perturbation in the Gulf of Alaska; Suryan, R.M., Lindeberg, M.R., Aderhold, D.R., Eds.; Exxon Valdez Oil Spill Trustee Council: Anchorage, AK, USA, 2020. [Google Scholar]
- KBNERR (Kachemak Bay National Estuarine Research Reserve). Harmful Algal Bloom 2019 Progress Report; Kachemak Bay National Estuarine Research Reserve Alaska Center for Conservation Science: Anchorage, AK, USA, 2019; Available online: https://accs.uaa.alaska.edu/wp-content/uploads/2019-KBNERR-HAB-Report.pdf (accessed on 2 March 2021).
- Anderson, D.M.; Richlen, M.L.; Lefebvre, K.A. Harmful Algal Blooms in the Arctic [in Arctic Report Card 2018]. Available online: https://arctic.noaa.gov/Report-Card/Report-Card-2018/ArtMID/7878/ArticleID/789/Harmful-Algal-Blooms-in-the-Arctic (accessed on 15 March 2021).
- Anderson, D.M.; Fachon, E.; Robert, S.P.; Lin, P.; Alexis, D.F.; Mindy, L.R.; Uva, V.; Michael, L.B.; McRaven, L.; Bahr, F.; et al. Evidence for massive and recurrent toxic blooms of Alexandrium catenella in the Alaskan Arctic. Proc. Natl. Acad. Sci. USA 2021, 118, e2107387118. [Google Scholar] [CrossRef] [PubMed]
- de Nadal, E.; Ammerer, G.; Posas, F. Controlling gene expression in response to stress. Nat. Rev. Genet. 2011, 12, 833–845. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Gene Function |
|---|---|
| S9 | Ribosomal Reference Gene [32] |
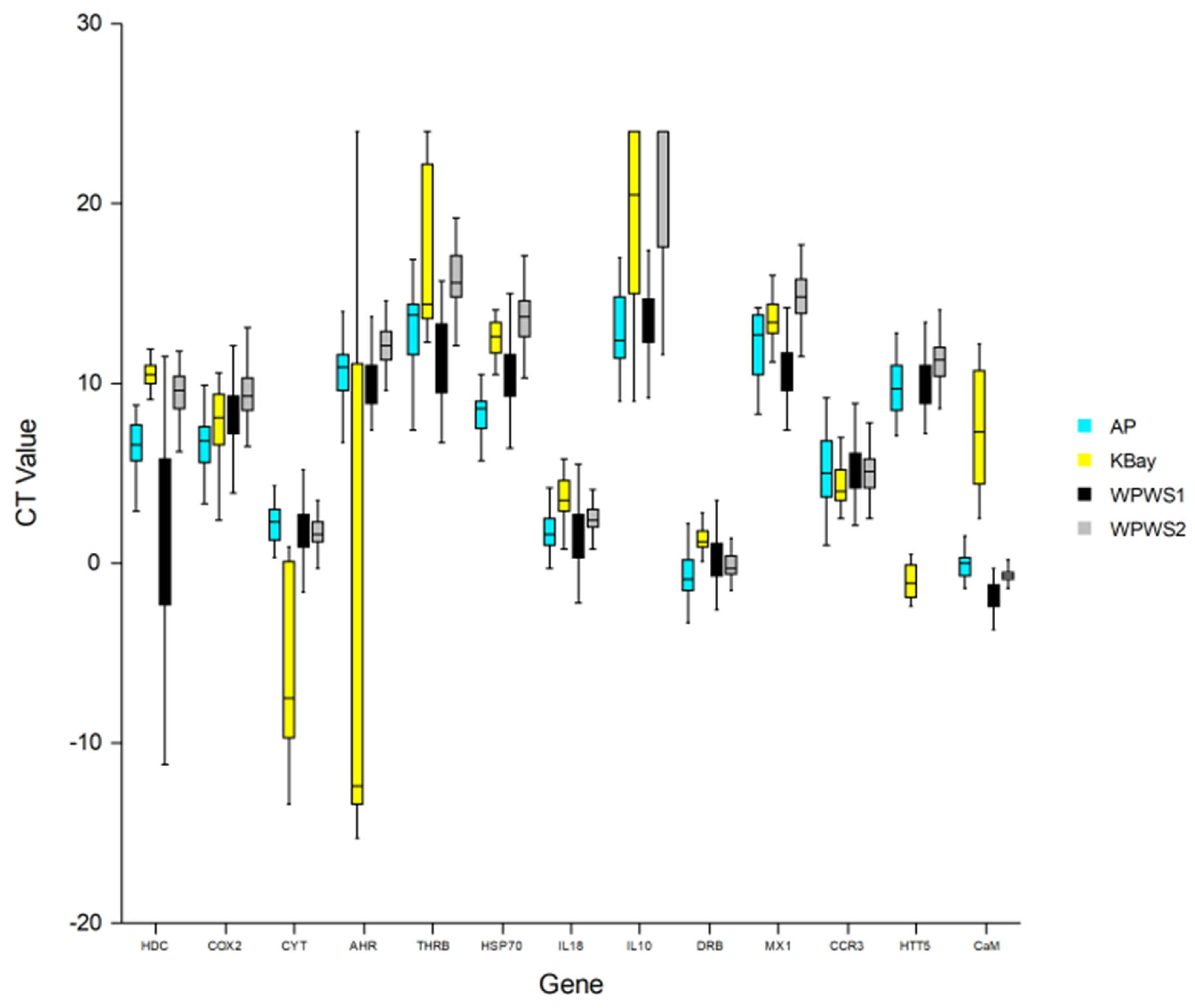
| HDC | The HDCMB21P gene codes for a translationally controlled tumor protein (TCTP) implicated in cell growth, cell cycle progression, malignant transformation, tumor progression, and in the protection of cells against various stress conditions and apoptosis [38,39,40]. The upregulation of HDC is indicative of the development or existence of cancer. Environmental triggers may be responsible for the population-based upregulation of HDC. HDC expression is known to increase with exposure to carcinogenic compounds, such as polycyclic aromatic hydrocarbons [41,42,43]. |
| COX2 | Cyclooxygenase-2 catalyzes the production of prostaglandins that are responsible for promoting inflammation [44]. Cox2 is responsible for the conversion of arachidonic acid to prostaglandin H2, a lipoprotein critical to the promotion of inflammation [45]. The upregulation of Cox2 is indicative of cellular or tissue damage and an associated inflammatory response. |
| CYT | The complement cytolysis inhibitor protects against cell death [46]. The upregulation of CYT is indicative of cell or tissue death. It is now believed that domoic acid-induced altered Ca2+ homeostasis is key in excitotoxic apoptosis, which is consistent with our finding of significantly higher levels of CYT in KBay otters [47]; increased levels of CYT have also been associated with cardiomyopathy [48,49]. |
| AHR | The arylhydrocarbon receptor responds to classes of environmental toxicants including polycyclic aromatic hydrocarbons, polyhalogenated hydrocarbons, dibenzofurans, and dioxin [50]. Depending upon the ligand, AHR signaling can modulate T-regulatory (TREG) (immune-suppressive) or T-helper type 17 (TH17) (pro-inflammatory) immunologic activity [51,52]. Wang et al. [53] were the first to identify substantial activation of AHR by domoic acid exposure in fish, a transcriptional response of phase I XME through ligand-activated AHR and ARNT to domoic acid exposure. AHR binds to toxins, initiating a detoxification cascade and an altered immune response. Activation of the AHR pathway also contributes to cardiac malformation [54]. |
| THRB | The thyroid hormone receptor beta can be used as a mechanistically based means of characterizing the thyroid-toxic potential of complex contaminant mixtures [55]. Thus, increases in THR expression may indicate exposure to organic compounds, including PCBs, and associated potential health effects, such as developmental abnormalities and neurotoxicity [55]. Hormone-activated transcription factors bind DNA in the absence of hormones, usually leading to transcriptional repression [56]. |
| HSP70 | The heat shock protein 70 is produced in response to thermal or other stress [57,58]. In addition to being expressed in response to a wide array of stressors (including hyperthermia, oxygen radicals, heavy metals, and ethanol), heat shock proteins act as molecular chaperones [59]. For example, heat shock proteins aid in the transport of the AHR/toxin complex in the initiation of detoxification [60]. |
| IL-18 | Interleukin-18 is a pro-inflammatory cytokine [44]. It plays an important role in inflammation and host defense against microbes [61]. |
| IL-10 | Interleukin-10 is an anti-inflammatory cytokine [44]. Levels of IL-10 have been correlated with the relative health of free-ranging harbor porpoises, e.g., increased amounts of IL-10 correlated with chronic disease, whereas the cytokine was relatively reduced in apparently fit animals experiencing acute disease [62]. The association of IL-10 expression with chronic disease has also been documented in humans [63]. |
| DRB | A component of the major histocompatibility complex, the DRB class II gene, is responsible for the binding and presentation of processed antigen to TH lymphocytes, thereby facilitating the initiation of an immune response [44,64]. The upregulation of MHC genes has been positively correlated with parasite load [65], whereas the downregulation of MHC has been associated with contaminant exposure [66]. |
| Mx1 | The Mx1 gene responds to viral infection [67]. Vertebrates have an early strong innate immune response against viral infection, characterized by the induction and secretion of cytokines that mediate an antiviral state, leading to the upregulation of the MX-1 gene [68]. |
| CCR3 | The chemokine receptor 3 binds at least seven different chemokines, and it is expressed on eosinophils, mast cells (MC), and a subset of Th cells (Th2) that generate cytokines implicated in mucosal immune responses [69,70]. The upregulation of CCR3 occurs in the presence of parasites [69,70]. |
| HTT5 | The serotonin transport gene codes for an integral membrane protein that transports the neurotransmitter serotonin from the synaptic spaces into presynaptic neurons. This transport of serotonin by the SERT protein terminates the action of serotonin and recycles it in a sodium-dependent manner [71,72]. Algal toxins have been associated with increased expression of HTT5 [73]; at the cellular level, domoic acid is an excitatory amino acid analogue of glutamate, a major excitatory neurotransmitter in the brain known to activate glutamate receptors [74]. |
| CaM | Calmodulin (CaM) is a small acidic Ca2+-binding protein, with a structure and function that is highly conserved in all eukaryotes. CaM activates various Ca2+-dependent enzyme reactions, thereby modulating a wide range of cellular events, including metabolism control, muscle contraction, exocytosis of hormones and neurotransmitters, and cell division and differentiation [75]. CaM has also been reported to be a pivotal calcium metabolism regulator in shell formation [76]. Algal toxicity is associated with increased intracellular Ca2+ [74,77,78,79,80,81,82]. This intracellular excess is toxic to the cells and triggers the activation of several detrimental cascading effects [47]. |
| Gene | Primer Name | FP1 | Primer Name | RP1rc | Expected Amplicon (bp) |
|---|---|---|---|---|---|
| HDC | Enlu HDC F1 | ATGTTCTCCGACATCTACAAGATCC | Enlu HDC R1rc | GTTTCCTGCAGGTGATGGTTCATG | 82.6 |
| COX2 | Mv COX2 F1 | CATTCCTGATCCCCAGGGCAC | Mv COX2 R1rc | GTCCACCCCATGGCCCAGTC | 79.4 |
| CYT | Mv CYT F1 | GCTGGACGAGCAGTTTAGCTGG | Mv CYT R1rc | GACGCCAGAGGGAGCACTGG | 81.8 |
| AHR | Enlu AHR F1 | GCGCTGAGTACCATATACGGATGA | Enlu AHR R1rc | CACTAAGCGTGCATTAGACTGAAC | 76.8 |
| THRB | Mv THRB F1 | GGACAAACCGAAGCACTGTCCAG | Mv THRB R1rc | GGAATATYGAGCTAAGTCCAAGTGG | 81.8 |
| HSP70 | Mv HSP70 F | CCAGGTGGCGCTGAACCCGC | Mv HSP70 Rrc | CCTTGTAGCTCACCTGCACCTTG | 85.6 |
| IL-18 | Mv IL-18 F | GTACAGAAAACGCATCCCATACC | Mv IL-18 Rrc | CTGGAGGTCTCATTTCCTTAAAGG | 76.2 |
| IL-10 | Mv IL-10 F2 | GACTTTAAGRGTTACCTGGGTTGC | Mv IL-10 R2rc | TCCACSGCCTTGCTCTTRTTYTC | 83.7 |
| DRB | Mv DRB F | CGGCGAGTGGAGCCTATAGTG | Mv DRB Rrc | CTCCTCTTCCTGGCCATTCCG | 81.0 |
| Mx1 | Enlu Mx1 F | CAAGCAGCTCATCAGGAAGTACA | Enlu Mx1 Rrc | GGTGGCGATGTCCACGTT | 79.5 |
| S9 | Mv S9 F | CCAGCGCCACATCAGGGTCCG | Mv S9 Rrc | CCCTGGCCTTTCTTGGCGTTC | 83.4 |
| CCR3 | Enlu CCR3 F | CTGCTGGGCAATGTGGTGGTG | Enlu CCR3 Rrc | GAAAGAGCAAGTCAGAAATGGCC | 106 |
| CaM | Enlu CaM F1 | GCGAGGCATTCCGAGTC | Enlu CaM R1rc | TCTGATCATTTCATCTACTTCT | 122 |
| HTT5 | Enlu HTT5 F2 | TCCTCCTGCCCTACACGA | Enlu HTT5 R2rc | CGGTGGTACTGGCCCAG | 82 |
| Genes | Kachemak Bay (2019) | Alaska Peninsula (2009) | WPWS1 (2006–2008) | WPWS2 (2010–2012) | ANOVA p Value |
|---|---|---|---|---|---|
| N = 20 | N = 27 | N = 81 | N = 87 | ||
| Mean (SD) | Mean (SD) | Mean (SD) | Mean (SD) | ||
| HDC | 10.68 a (1.21) | 6.39 b (1.73) | 1.05 b (4.79) | 9.18 a (1.85) | 0.00 |
| COX2 | 7.73 a,b (2.00) | 6.77 b (1.78) | 8.19 a (1.76) | 9.44 c (1.53) | 0.00 |
| CYT | −2.64 a (12.10) | 2.29 b (1.28) | 1.61 b (2.30) | 1.76 b (1.02) | 0.00 |
| AHR | −2.41 a (14.21) | 10.64 a (1.89) | 10.41 a (2.38) | 12.16 b (1.39) | 0.00 |
| THRB | 16.46 a,c 4.57 | 13.40 a 3.20 | 11.87 b 3.32 | 16.27 c 2.81 | 0.00 |
| HSP70 | 12.42 a,c (1.07) | 8.65 b (1.57) | 10.11 b (2.30) | 13.91 c (2.56) | 0.00 |
| IL-18 | 3.55 a (1.22) | 2.65 b,c (4.42) | 1.91 c (3.02) | 2.45 b (1.02) | 0.063 |
| IL-10 | 19.26 a,c (5.10) | 13.32 b (2.96) | 13.75 b (2.96) | 20.69 c (3.78) | 0.00 |
| DRB | 2.42 a (5.12) | −0.87 c (1.54) | 0.37 b,d (1.46) | −0.073 c,d (0.83) | 0.00 |
| MX1 | 13.47 a,c (1.18) | 12.90 a (3.62) | 10.58 b (1.65) | 15.17 c (2.35) | 0.00 |
| CCR3 | 4.31 (1.17) | 5.31 (2.13) | 5.16 (1.40) | 5.11 (1.15) | 0.074 |
| HTT5 | −1.03 a (0.92) | 9.76 b (1.49) | 9.99 b (1.27) | 11.07 c (2.20) | 0.00 |
| CaM | 7.48 a (3.34) | −0.093 b (0.70) | −1.75 c (0.92) | −0.67 b (0.40) | 0.00 |
| Gene | Pearson Correlation | p Value |
|---|---|---|
| HDC | 0.39 | 0.39 |
| COX2 | 0.097 | 0.84 |
| CYT | −0.31 | 0.50 |
| AHR | 0.73 | 0.062 |
| THRB | 0.18 | 0.69 |
| HSP70 | 0.52 | 0.23 |
| IL-18 | 0.31 | 0.49 |
| IL-10 | 0.035 | 0.94 |
| DRB | 0.53 | 0.22 |
| Mx1 | 0.16 | 0.73 |
| CCR3 | 0.74 | 0.057 |
| CaM | 0.23 | 0.62 |
| HTT5 | 0.12 | 0.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowen, L.; Knowles, S.; Lefebvre, K.; St. Martin, M.; Murray, M.; Kloecker, K.; Monson, D.; Weitzman, B.; Ballachey, B.; Coletti, H.; et al. Divergent Gene Expression Profiles in Alaskan Sea Otters: An Indicator of Chronic Domoic Acid Exposure? Oceans 2022, 3, 401-418. https://doi.org/10.3390/oceans3030027
Bowen L, Knowles S, Lefebvre K, St. Martin M, Murray M, Kloecker K, Monson D, Weitzman B, Ballachey B, Coletti H, et al. Divergent Gene Expression Profiles in Alaskan Sea Otters: An Indicator of Chronic Domoic Acid Exposure? Oceans. 2022; 3(3):401-418. https://doi.org/10.3390/oceans3030027
Chicago/Turabian StyleBowen, Lizabeth, Susan Knowles, Kathi Lefebvre, Michelle St. Martin, Michael Murray, Kim Kloecker, Daniel Monson, Benjamin Weitzman, Brenda Ballachey, Heather Coletti, and et al. 2022. "Divergent Gene Expression Profiles in Alaskan Sea Otters: An Indicator of Chronic Domoic Acid Exposure?" Oceans 3, no. 3: 401-418. https://doi.org/10.3390/oceans3030027