Ontogeny of Orientation during the Early Life History of the Pelagic Teleost Mahi-Mahi, Coryphaena hippurus Linnaeus, 1758

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Rearing and Handling of the Early Life History Stages of Mahi-Mahi
2.3. DISC Experiment
2.4. Data Analysis
2.5. Data Availability
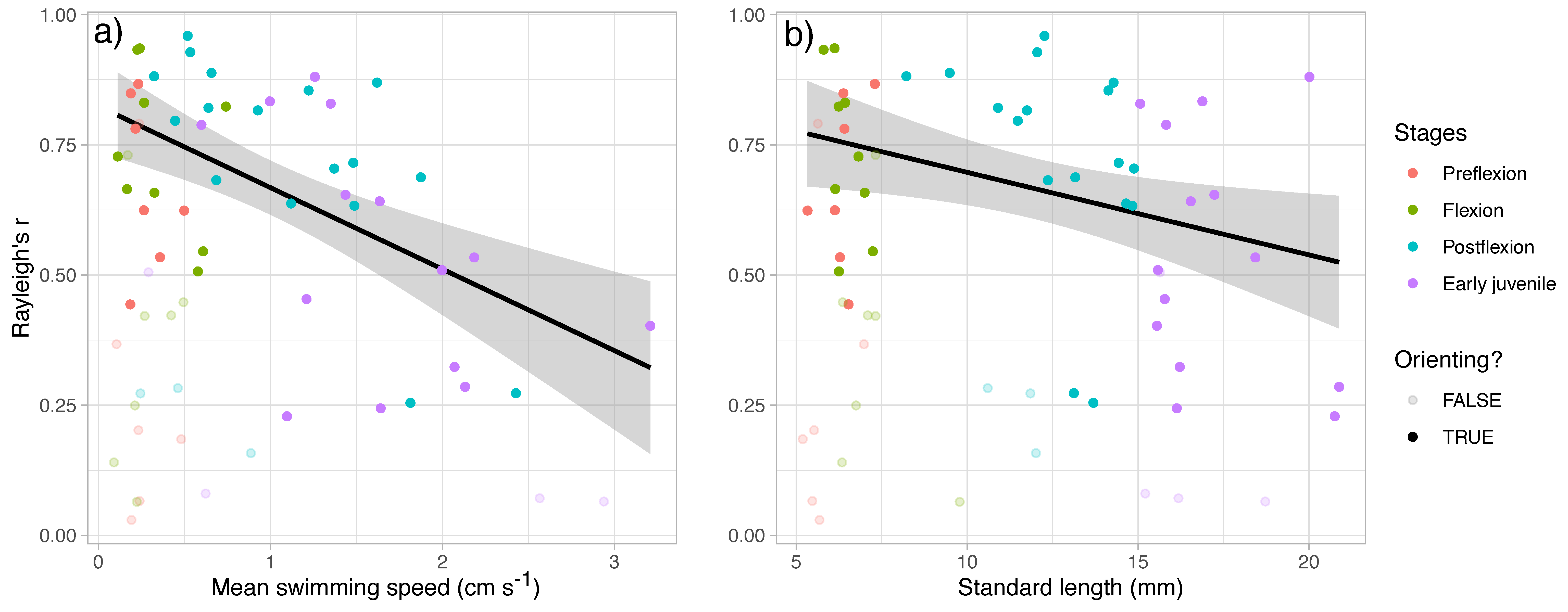
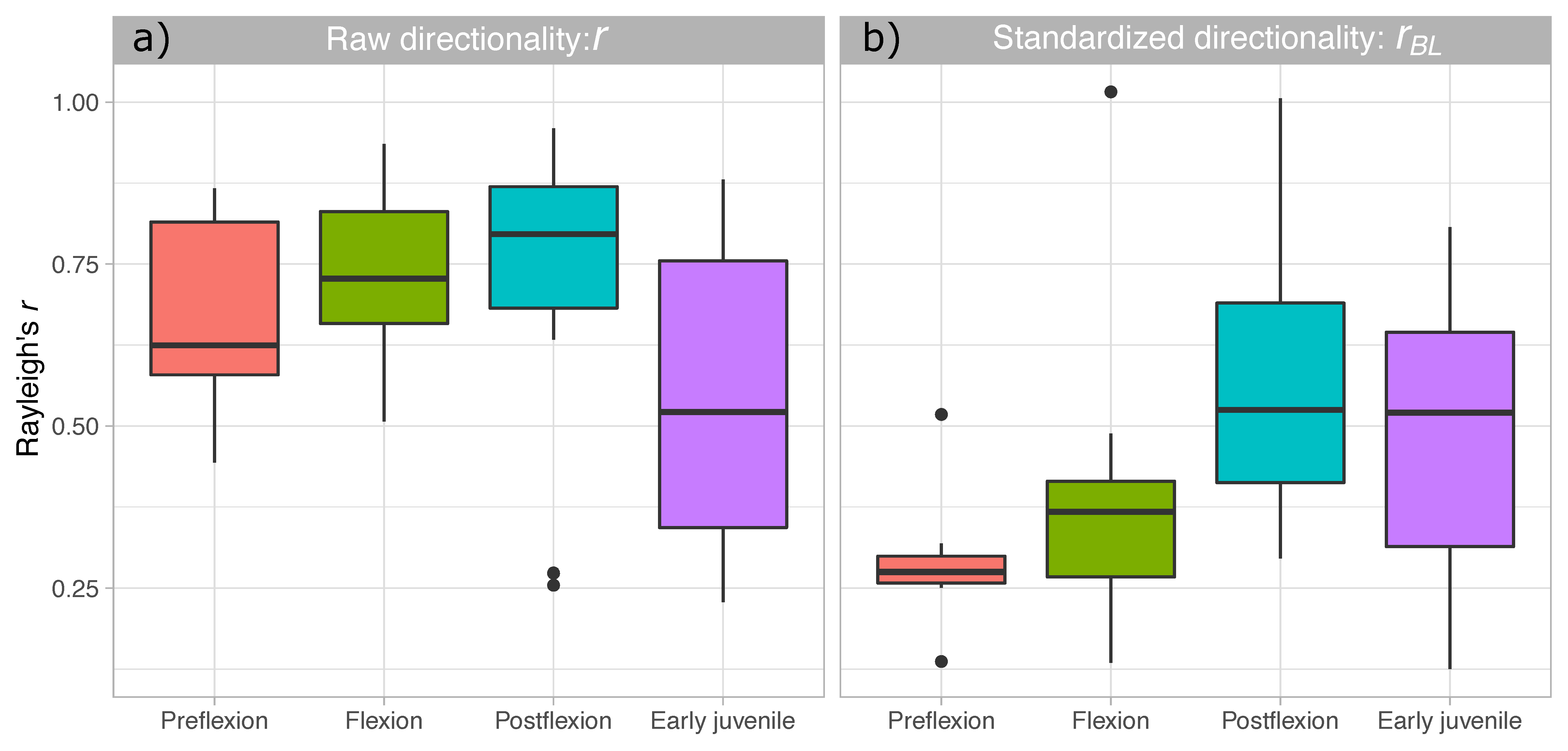
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ragatz, L.; Jiang, Z.Y.; Bauer, C.E.; Gest, H. Macroscopic phototactic behavior of the purple photosynthetic bacterium Rhodospirillum centenum. Arch Microbiol. 1995, 163, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Masuda, R.; Tsukamoto, K. Morphological development in relation to phototaxis and rheotaxis in the striped jack, Pseudocaranx dentex. Mar. Freshw. Behav. Physiol. 1996, 28, 75–90. [Google Scholar] [CrossRef]
- Ina, Y.; Sakakura, Y.; Yamada, T.; Kumon, K.; Eba, T.; Konishi, J.; Takashi, T.; Gen, K.; Tanaka, Y.; Hashimoto, H. Development of phototaxis in the early life stages of Pacific bluefin tuna Thunnus orientalis. Fish. Sci. 2017, 83, 537–542. [Google Scholar] [CrossRef]
- Partridge, G.; Benetti, D.; Stieglitz, J.; Hutapea, J.; McIntyre, A.; Chen, B.; Hutchinson, W.; Scholey, V. The effect of a 24-hour photoperiod on the survival, growth and swim bladder inflation of pre-flexion yellowfin tuna (Thunnus albacares) larvae. Aquaculture 2011, 318, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Nicolaisen, O.; Bolla, S. Behavioural responses to visual environment in early stage Atlantic cod Gadus morhua L. larvae. Aquac Res. 2016, 47, 189–198. [Google Scholar] [CrossRef]
- Chen, X.; Engert, F. Navigational strategies underlying phototaxis in larval zebrafish. Front. Syst. Neurosci. 2014, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Leis, J.M. Are larvae of demersal fishes plankton or nekton? Adv. Mar. Biol. 2006, 51, 57–141. [Google Scholar] [CrossRef]
- Irisson, J.; Paris, C.B.; Leis, J.M.; Yerman, M.N. With a little help from my friends: Group orientation by larvae of a coral reef fish. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Faillettaz, R.; Blandin, A.; Paris, C.B.; Koubbi, P.; Irisson, J.-O. Sun-compass orientation in mediterranean fish larvae. PLoS ONE 2015, 10, e0135213. [Google Scholar] [CrossRef] [Green Version]
- Leis, J.; Siebeck, U.; Hay, A.; Paris, C.; Chateau, O.; Wantiez, L. In situ orientation of fish larvae can vary among regions. Mar. Ecol. Prog. Ser. 2015, 537, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Paris, C.B.; Atema, J.; Irisson, J.-O.; Kingsford, M.; Gerlach, G.; Guigand, C.M. Reef odor: A wake up call for navigation in reef fish larvae. PLoS ONE 2013, 8, e72808. [Google Scholar] [CrossRef] [PubMed]
- Leis, J.M. Ontogeny of behaviour in larvae of marine demersal fishes. Ichthyol. Res. 2010, 57, 325–342. [Google Scholar] [CrossRef]
- Leis, J.M.; Hay, A.C.; Clark, D.L.; Chen, I.S.; Shao, K.T. Behavioral ontogeny in larvae and early juveniles of the giant trevally (Caranx ignobilis) (Pisces: Carangidae). Fish. Bull. 2004, 104, 401–414. [Google Scholar]
- Leis, J.M.; Hay, A.C.; Trnski, T. In situ ontogeny of behaviour in pelagic larvae of three temperate, marine, demersal fishes. Mar. Biol. 2006, 148, 655–669. [Google Scholar] [CrossRef]
- Irisson, J.-O. Approche Comportementale de la Dispersion Larvaire en Milieu Marin. Sciences du Vivant [q-bio]. Ecole pratique des hautes études—EPHE PARIS, 2008. HAL Id: tel-00344625. Available online: https://tel.archives-ouvertes.fr/file/index/docid/344625/filename/THESE-Irisson.pdf (accessed on 1 October 2020).
- Staaterman, E.; Paris, C.B. Modelling larval fish navigation: The way forward. ICES J. Mar. Sci. 2014, 71, 918–924. [Google Scholar] [CrossRef] [Green Version]
- Faillettaz, R.; Paris, C.B.; Irisson, J.-O. Larval fish swimming behavior alters dispersal patterns from marine protected areas in the North-Western Mediterranean Sea. Front. Mar. Sci. 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Staaterman, E.; Paris, C.B.; Helgers, J. Orientation behavior in fish larvae: A missing piece to Hjort’s critical period hypothesis. J. Theor. Biol. 2012, 304, 188–196. [Google Scholar] [CrossRef]
- Faillettaz, R.; Durand, E.; Paris, C.B.; Koubbi, P.; Irisson, J.O. Swimming speeds of Mediterranean settlement-stage fish larvae nuance Hjort’s aberrant drift hypothesis. Limnol. Oceanogr. 2018, 63, 509–523. [Google Scholar] [CrossRef]
- Bode, M.; Leis, J.M.; Mason, L.B.; Williamson, D.H.; Harrison, H.B.; Choukroun, S.; Jones, G.P. Successful validation of a larval dispersal model using genetic parentage data. PLoS Biol. 2019, 17, e3000380. [Google Scholar] [CrossRef] [Green Version]
- Leis, J.J.M. Behaviour as input for modelling dispersal of fish larvae: Behaviour, biogeography, hydrodynamics, ontogeny, physiology and phylogeny meet hydrography. Mar. Ecol. Prog. Ser. 2007, 347, 185–193. [Google Scholar] [CrossRef]
- Swearer, S.E.; Treml, E.A.; Shima, J.S. A review of biophysical models of marine larval dispersal. In Oceanography and Marine Biology; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar] [CrossRef]
- Selden Burke, J.; Tanaka, M.; Seikai, T. Influence of light and salinity on behaviour of larval Japanese flounder (Paralichthys olivaceus) and implications for inshore migration. Neth. J. Sea Res. 1995, 34, 59–69. [Google Scholar] [CrossRef]
- Champalbert, G.; Le Direach-Boursier, L. Influence of light and feeding conditions on swimming activity rhythms of larval and juvenile turbot Scophthalmus maximus L.: An experimental study. J. Sea Res. 1998, 40, 333–345. [Google Scholar] [CrossRef]
- Blaxter, B.Y.J.H.S. Visual thresholds and spectral sensitivity of herring larvae. J. Exp. Biol. 1968, 48, 39–53. [Google Scholar]
- Hartmann, S.; Vogt, R.; Kunze, J.; Rauschert, A.; Kuhnert, K.-D.; Wanzenböck, J.; Lamatsch, D.K.; Witte, K. Zebrafish larvae show negative phototaxis to near-infrared light. PLoS ONE 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Montgomery, J.C.; Tolimieri, N.; Haine, O.S. Active habitat selection by pre-settlement reef fishes. Fish Fish. 2001, 2, 261–277. [Google Scholar] [CrossRef]
- Simpson, S.D.; Meekan, M.; Montgomery, J.; McCauley, R.; Jeffs, A. Homeward sound. Science (80-) 2005, 308, 221. [Google Scholar] [CrossRef]
- Teodósio, M.A.; Paris, C.B.; Wolanski, E.; Morais, P. Biophysical processes leading to the ingress of temperate fish larvae into estuarine nursery areas: A review. Estuar. Coast. Shelf Sci. 2016, 183, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Atema, J.; Gerlach, G.; Paris, C.B. Sensory biology and navigation behavior of reef fish larvae. In Ecology of Fishes on Coral Reefs; Mora, C., Ed.; Cambridge University Press: Cambridge, UK, 2015; pp. 3–15. [Google Scholar]
- O’Connor, J.; Muheim, R. Pre-settlement coral-reef fish larvae respond to magnetic field changes during the day. J. Exp. Biol. 2017, 220, 2874–2877. [Google Scholar] [CrossRef] [Green Version]
- Cresci, A.; Paris, C.B.; Foretich, M.A.; Durif, C.M.; Shema, S.D.; O’Brien, C.; Vikebø, F.B.; Skiftesvik, A.-B.; Browman, H.I. Atlantic haddock (Melanogrammus aeglefinus) larvae have a magnetic compass that guides their orientation. iScience 2019, 19, 1173–1178. [Google Scholar] [CrossRef] [Green Version]
- Berenshtein, I.; Kiflawi, M.; Shashar, N.; Wieler, U.; Agiv, H.; Paris, C.B. Polarized light sensitivity and orientation in coral reef fish post-larvae. PLoS ONE 2014, 9, e88468. [Google Scholar] [CrossRef] [Green Version]
- Kheradmand, B.; Nieh, J.C. The role of landscapes and landmarks in bee navigation: A review. Insects 2019, 10, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveros, A.J.; Srygley, R.B. Magnetic compasses in insects. In Encycopedia of Animal Behavior, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; pp. 588–597. [Google Scholar]
- Mouritsen, H.; Atema, J.; Kingsford, M.J.; Gerlach, G. Sun compass orientation helps coral reef fish larvae return to their natal reef. PLoS ONE 2013, 8, e66039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palko, B.J.; Beardsley, G.L.; Richards, W.J. Synopsis of the Biological Data on Dolphin-Fishes, Coryphaena Hippurus Linnaeus and Coryphaena Equiselis Linnaeus; FAO Fisheries Synopsis Report; U.S. Department of Commerce: Washington, DC, USA, 1982; Volume 130, p. 34. [Google Scholar]
- Stieglitz, J.D.; Hoenig, R.H.; Kloeblen, S.; Tudela, C.E.; Grosell, M.; Benetti, D.D. Capture, transport, prophylaxis, acclimation, and continuous spawning of Mahi-mahi (Coryphaena hippurus) in captivity. Aquaculture 2017, 479, 1–6. [Google Scholar] [CrossRef]
- Kraul, S. Larviculture of the Mahimahi Coryphaena hippurus in Hawaii, USA. J. World Aquac. Soc. 1993, 24, 410–421. [Google Scholar] [CrossRef]
- Benetti, D.D.; Sardenberg, B.; Welch, A.; Hoenig, R.; Orhun, M.R.; Zink, I. Intensive larval husbandry and fingerling production of cobia Rachycentron canadum. Aquaculture 2008, 281, 22–27. [Google Scholar] [CrossRef]
- Irisson, J.-O.; Guigand, C.; Paris, C.B. Detection and quantification of marine larvae orientation in the pelagic environment. Limnol. Oceanogr. Methods 2009, 7, 664–672. [Google Scholar] [CrossRef]
- Paris, C.B.; Guigand, C.M.; Irisson, J.; Fisher, R.; Alessandro, D. Orientation with no frame of reference (OWNFOR): A novel system to observe and quantify orientation in reef fish larvae. In Caribbean Connectivity: Implications for Marine Protected Area Management, Proceedings of the Special Symposium: 59th Annual Meeting of the Gulf and Caribbean Fisheries Institute. Belize city, Belize, 9–11 November 2006; Grober-Dunsmore, R., Keller, B.D., Eds.; Marine Sanctuaries Conservation Series NMSP- 08-xx. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Sanctuary Program: Silver Spring, MD, USA, 2008; pp. 54–64. [Google Scholar]
- Magnuson, J.T.; Stieglitz, J.D.; Garza, S.A.; Benetti, D.D.; Grosell, M.; Roberts, A.P. Development of visual function in early life stage mahi-mahi (coryphaena hippurus). Mar. Freshw. Behav. Physiol. 2020, in press. [Google Scholar] [CrossRef]
- Batschelet, E. Circular Statistics in Biology; Academic Press: London, UK, 1981. [Google Scholar]
- Leis, J.M. Measurement of swimming ability in larval marine fishes: Comparison of critical speed with in situ speed. Mar. Ecol. Prog. Ser. 2020, in press. [Google Scholar] [CrossRef]
- Stephen, M.A. Use of the kolmogorov-smirnov, cramer-von mises and related statistics without extensive tables. J. R. Stat. Soc. Ser. B. 1970, 32, 115–122. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; p. 201. Available online: http://www.r-project.org/ (accessed on 1 October 2020).
- Agostinelli, C.; Lund, U. R Package Circular: Circular Statistics (version 0.4–93, 2017). Available online: https://r-forge.r-project.org/projects/circular/ (accessed on 1 October 2020).
- Wickham, H. Tidyverse: Easily Install and Load the ‘Tidyverse’, R package version 1.2. 1; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Forsgren, E.; Dupont, S.; Jutfelt, F.; Amundsen, T. Elevated CO2 affects embryonic development and larval phototaxis in a temperate marine fish. Ecol. Evol. 2013, 3, 3637–3646. [Google Scholar] [CrossRef]
- Blaxter, J. Pattern and variety in development. Fish Physiol. 1988, 11, 1–58. [Google Scholar] [CrossRef]
- Downie, A.T.; Illing, B.; Faria, A.M.; Rummer, J.L. Swimming performance of marine fish larvae: Review of a universal trait under ecological and environmental pressure. Rev. Fish Biol. Fish. 2020, 30, 93–108. [Google Scholar] [CrossRef]
- Fisher, R.; Hogan, J.D. Morphological predictors of swimming speed: A case study of pre-settlement juvenile coral reef fishes. J. Exp. Biol. 2007, 210, 2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berenshtein, I.; Faillettaz, R.; Irisson, J.-O.; Kiflawi, M.; Siebeck, U.E.; Leis, J.M.; Paris, C.B. Fish larvae consistently use external cues for orientation. In Proceedings of the 44th Larval Fish Conference, Mystic, CT, USA, 21–26 June 2020. [Google Scholar]
- Foretich, M.A.; Paris, C.B.; Grosell, M.; Stieglitz, J.D.; Benetti, D.D. Dimethyl sulfide is a chemical attractant for reef fish larvae. Sci. Rep. 2017, 7, 2498. [Google Scholar] [CrossRef]
- Horváth, G.; Varjú, D. Underwater refraction-polarization patterns of skylight perceived by aquatic animals through Snell’s window of the flat water surface. Vis. Res. 1995, 35, 1651–1666. [Google Scholar] [CrossRef] [Green Version]
- Rossi, A.; Irisson, J.-O.; Levaray, M.; Pasqualini, V.; Agostini, S. Orientation of Mediterranean fish larvae varies with location. Mar. Biol. 2019, 166, 100. [Google Scholar] [CrossRef]
- Fuiman, L.A.; Higgs, D.M. Ontogeny, growth and the recruitment process. In Early Life History and Recruitment in Fish Populations; Springer: Dodlerk, The Netherlands, 1997; pp. 225–249. [Google Scholar] [CrossRef]
- Codling, E.A.; Hill, N.A. Sampling rate effects on measurements of correlated and biased random walks. J. Theor. Biol. 2005, 233, 573–588. [Google Scholar] [CrossRef]
- Viswanathan, G.M.; Buldyrev, S.V.; Havlin, S.; Da Luz, M.G.E.; Raposo, E.P.; Stanley, H.E. Optimizing the success of random searches. Nature 1999, 401, 911–914. [Google Scholar] [CrossRef]
- Benetti, D.D.; Iversen, E.S.; Ostrowski, A.C. Growth rates of captive dolphin, Coryphaena hippurus, in Hawaii. Fish. Bull. 1995, 93, 152–157. [Google Scholar]
- Graeb, B.D.S.; Dettmers, J.M.; Wahl, D.H.; Cáceres, C.E. Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Trans. Am. Fish. Soc. 2004, 133, 504–514. [Google Scholar] [CrossRef]
- Spudich, J.L.; Yang, C.-S.; Jung, K.-H.; Spudich, E.N. Retinylidene proteins: structures and functions from archaea to humans. Annu. Rev. Cell Dev. Biol. 2000, 16, 365–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faillettaz, R.; Johnson, E.; Dahlmann, P.; Syunkova, A.; Stieglitz, J.; Benetti, D.; Grosell, M.; Paris, C.B. Ontogeny of Orientation during the Early Life History of the Pelagic Teleost Mahi-Mahi, Coryphaena hippurus Linnaeus, 1758. Oceans 2020, 1, 237-250. https://doi.org/10.3390/oceans1040017
Faillettaz R, Johnson E, Dahlmann P, Syunkova A, Stieglitz J, Benetti D, Grosell M, Paris CB. Ontogeny of Orientation during the Early Life History of the Pelagic Teleost Mahi-Mahi, Coryphaena hippurus Linnaeus, 1758. Oceans. 2020; 1(4):237-250. https://doi.org/10.3390/oceans1040017
Chicago/Turabian StyleFaillettaz, Robin, Eve Johnson, Patrick Dahlmann, Alexandra Syunkova, John Stieglitz, Daniel Benetti, Martin Grosell, and Claire B. Paris. 2020. "Ontogeny of Orientation during the Early Life History of the Pelagic Teleost Mahi-Mahi, Coryphaena hippurus Linnaeus, 1758" Oceans 1, no. 4: 237-250. https://doi.org/10.3390/oceans1040017