

Particles in Raw Sheep Milk Can Modulate the Inflammatory Response in THP-1, a Human Monocyte Cell Line, In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Milk Collection
2.3. Isolation of SMEVs
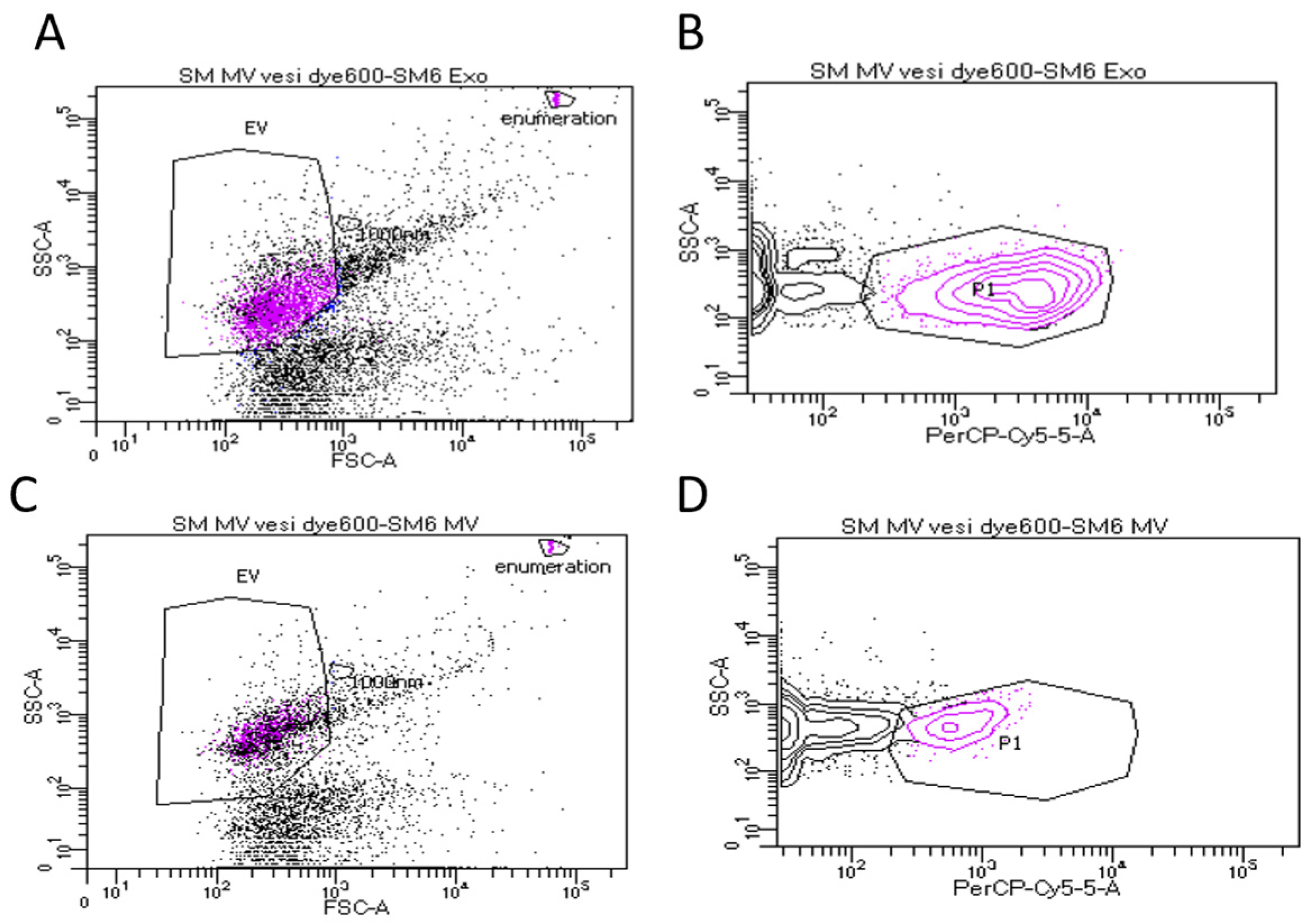
2.4. Flow Cytometry
2.5. Determination of Protein Concentration of EVs
2.6. RNA Isolation and miRNA Analysis
2.7. Cell Culture
2.8. Reactive Oxygen Species (ROS) Assays
2.9. Measurement of CCL2 Secretion
2.10. Statistical Analysis
3. Results
3.1. Characterization of Sheep Milk Derived LEVs and sEVs
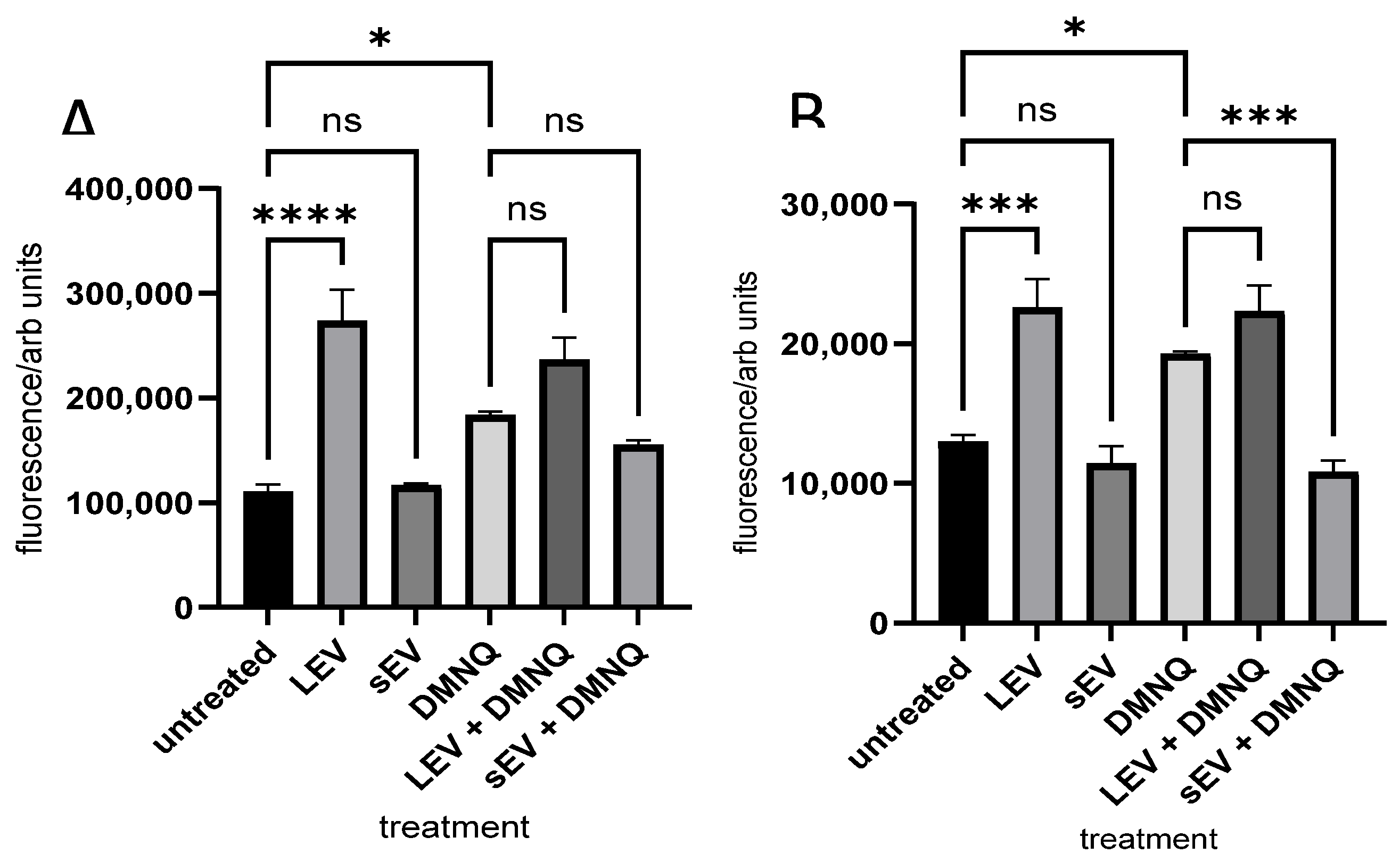
3.2. Monocyte and Macrophage ROS Production Induced by Sheep Milk-Derived LEVs and sEVs
3.3. Secretion of CCL2 Chemokine Induced by Sheep Milk-Derived LEVs and sEVs
3.4. miRNA Contents of Sheep Milk-Derived LEVs and sEVs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- AHDB. Dairy Market Dashboard. Available online: https://ahdb.org.uk/dairy-market-dashboard (accessed on 29 December 2023).
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited review: Milk lactose-Current status and future challenges in dairy cattle. J. Dairy. Sci. 2019, 102, 5883–5898. [Google Scholar] [CrossRef]
- Mee, J.F.; Boyle, L.A. Assessing whether dairy cow welfare is “better” in pasture-based than in confinement-based management systems. N. Z. Vet. J. 2020, 68, 168–177. [Google Scholar] [CrossRef]
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, Structure, and Digestive Dynamics of Milk From Different Species-A Review. Front. Nutr. 2020, 7, 577759. [Google Scholar] [CrossRef]
- Abou El Qassim, L.; Martínez, B.; Rodríguez, A.; Dávalos, A.; López de Las Hazas, M.C.; Menéndez Miranda, M.; Royo, L.J. Effects of Cow’s Milk Processing on MicroRNA Levels. Foods 2023, 12, 2950. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A compendium for extracellular vesicles with continuous community annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Chen, M.; Li, N.; Han, R.; Yang, Y.; Zheng, N.; Zhao, S.; Zhang, Y. Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review. Foods 2023, 12, 3755. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, K.; Lago-Novais, D.; Bevilacqua, C.; Mobuchon, L.; Crapart, N.; Faulconnier, Y.; Boby, C.; Carvalho, G.; Martin, P.; Leroux, C. Different miRNA contents between mammary epithelial cells and milk fat globules: A random or a targeted process? Mol. Biol. Rep. 2020, 47, 8259–8264. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, J.; Aguilar-Lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J. Nutr. 2017, 147, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; You, L.; Zhang, Z.; Cui, X.; Zhong, H.; Sun, X.; Ji, C.; Chi, X. Biological Properties of Milk-Derived Extracellular Vesicles and Their Physiological Functions in Infant. Front. Cell Dev. Biol. 2021, 9, 693534. [Google Scholar] [CrossRef]
- Benmoussa, A.; Lee, C.H.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef] [PubMed]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Gallier, S.; Van den Abbeele, P.; Prosser, C. Comparison of the Bifidogenic Effects of Goat and Cow Milk-Based Infant Formulas to Human Breast Milk in an. Front. Nutr. 2020, 7, 608495. [Google Scholar] [CrossRef] [PubMed]
- Arntz, O.J.; Pieters, B.C.; Oliveira, M.C.; Broeren, M.G.; Bennink, M.B.; de Vries, M.; van Lent, P.L.; Koenders, M.I.; van den Berg, W.B.; van der Kraan, P.M.; et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol. Nutr. Food Res. 2015, 59, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, A.; Santoro, J.; O’Driscoll, L. Extracellular vesicle separation from milk and infant milk formula using acid precipitation and ultracentrifugation. STAR Protoc. 2021, 2, 100821. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 3, 3.22.1–3.22.29. [Google Scholar] [CrossRef]
- Heinrich, L.F.; Andersen, D.K.; Cleasby, M.E.; Lawson, C. Long-term high fat feeding of rats results in increased numbers of circulating microvesicles with pro-inflammatory effects on endothelial cells. Br. J. Nutr. 2015, 113, 1704–1711. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Enderle, D.; Spiel, A.; Coticchia, C.M.; Berghoff, E.; Mueller, R.; Schlumpberger, M.; Sprenger-Haussels, M.; Shaffer, J.M.; Lader, E.; Skog, J.; et al. Characterization of RNA from Exosomes and Other Extracellular Vesicles Isolated by a Novel Spin Column-Based Method. PLoS ONE 2015, 10, e0136133. [Google Scholar] [CrossRef]
- Namgaladze, D.; Preiss, S.; Dröse, S.; Brandt, U.; Brüne, B. Phospholipase A2-modified low density lipoprotein induces mitochondrial uncoupling and lowers reactive oxygen species in phagocytes. Atherosclerosis 2010, 208, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Dommel, S.; Blüher, M. Does C-C Motif Chemokine Ligand 2 (CCL2) Link Obesity to a Pro-Inflammatory State? Int. J. Mol. Sci. 2021, 22, 1500. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front. Endocrinol. 2023, 14, 1217165. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.P.; Singh, N.P.; Murphy, E.A.; Price, R.L.; Fayad, R.; Nagarkatti, M.; Nagarkatti, P.S. Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine 2016, 77, 44–49. [Google Scholar] [CrossRef]
- Moadab, F.; Khorramdelazad, H.; Abbasifard, M. Role of CCL2/CCR2 axis in the immunopathogenesis of rheumatoid arthritis: Latest evidence and therapeutic approaches. Life Sci. 2021, 269, 119034. [Google Scholar] [CrossRef]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Liu, A. Chemokines in Prediabetes and Type 2 Diabetes: A Meta-Analysis. Front. Immunol. 2021, 12, 622438. [Google Scholar] [CrossRef]
- Puthumana, J.; Thiessen-Philbrook, H.; Xu, L.; Coca, S.G.; Garg, A.X.; Himmelfarb, J.; Bhatraju, P.K.; Ikizler, T.A.; Siew, E.D.; Ware, L.B.; et al. Biomarkers of inflammation and repair in kidney disease progression. J. Clin. Investig. 2021, 131, e139927. [Google Scholar] [CrossRef]
- Georgakis, M.K.; Bernhagen, J.; Heitman, L.H.; Weber, C.; Dichgans, M. Targeting the CCL2-CCR2 axis for atheroprotection. Eur. Heart J. 2022, 43, 1799–1808. [Google Scholar] [CrossRef]
- Martynowicz, H.; Janus, A.; Nowacki, D.; Mazur, G. The role of chemokines in hypertension. Adv. Clin. Exp. Med. 2014, 23, 319–325. [Google Scholar] [CrossRef]
- Chen, X.; Gao, C.; Li, H.; Huang, L.; Sun, Q.; Dong, Y.; Tian, C.; Gao, S.; Dong, H.; Guan, D.; et al. Identification and characterization of microRNAs in raw milk during different periods of lactation, commercial fluid, and powdered milk products. Cell Res. 2010, 20, 1128–1137. [Google Scholar] [CrossRef]
- Li, W.; Li, W.; Wang, X.; Zhang, H.; Wang, L.; Gao, T. Comparison of miRNA profiles in milk-derived extracellular vesicles and bovine mammary glands. Int. Dairy J. 2022, 134, 105444. [Google Scholar] [CrossRef]
- Quan, S.; Nan, X.; Wang, K.; Jiang, L.; Yao, J.; Xiong, B. Characterization of Sheep Milk Extracellular Vesicle-miRNA by Sequencing and Comparison with Cow Milk. Animals 2020, 10, 331. [Google Scholar] [CrossRef]
- Kirchner, B.; Pfaffl, M.W.; Dumpler, J.; von Mutius, E.; Ege, M.J. microRNA in native and processed cow’s milk and its implication for the farm milk effect on asthma. J. Allergy Clin. Immunol. 2016, 137, 1893–1895.e1813. [Google Scholar] [CrossRef]
- Lai, Y.C.; Fujikawa, T.; Maemura, T.; Ando, T.; Kitahara, G.; Endo, Y.; Yamato, O.; Koiwa, M.; Kubota, C.; Miura, N. Inflammation-related microRNA expression level in the bovine milk is affected by mastitis. PLoS ONE 2017, 12, e0177182. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ou-Yang, W.X.; Zhang, H.; Jiang, T.; Tang, L.; Tan, Y.F.; Luo, H.Y.; Xiao, Z.H.; Li, S.J. MiR-125b enhances autophagic flux to improve septic cardiomyopathy via targeting STAT3/HMGB1. Exp. Cell Res. 2021, 409, 112842. [Google Scholar] [CrossRef] [PubMed]
- Luoreng, Z.M.; Wei, D.W.; Wang, X.P. MiR-125b regulates inflammation in bovine mammary epithelial cells by targeting the NKIRAS2 gene. Vet. Res. 2021, 52, 122. [Google Scholar] [CrossRef] [PubMed]
- AHDB. Available online: https://ruminanthw.org.uk/wp-content/uploads/2021/05/Cattle-and-sheep-HW-priorities-survey-May-2021-FINAL.pdf (accessed on 29 December 2023).
- Whatford, L.; van Winden, S.; Häsler, B. A systematic literature review on the economic impact of endemic disease in UK sheep and cattle using a One Health conceptualisation. Prev. Vet. Med. 2022, 209, 105756. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.M.; Willis, Z.N.; Blakeley, M.; Lovatt, F.; Purdy, K.J.; Green, L.E. Bacterial species and their associations with acute and chronic mastitis in suckler ewes. J. Dairy. Sci. 2015, 98, 7025–7033. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.; Smith, E.M.; Green, L.E. A longitudinal study of factors associated with acute and chronic mastitis and their impact on lamb growth rate in 10 suckler sheep flocks in Great Britain. Prev. Vet. Med. 2016, 127, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J. Dairy. Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, M.A. microRNAs and the immune response. Trends Immunol. 2008, 29, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, Y.; Fan, Q.; Li, Z.; Qi, X.; Chen, F.; Xu, L.; Zhou, S.; Chen, X.; Li, Y.; et al. MiR-155 promotes acute organ injury in LPS-induced endotoxemic mice by enhancing CCL-2 expression in macrophages. Shock 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Barkema, H.W.; Gao, J.; Yang, J.; Wang, Y.; Kastelic, J.P.; Khan, S.; Liu, G.; Han, B. MicroRNA miR-223 modulates NLRP3 and Keap1, mitigating lipopolysaccharide-induced inflammation and oxidative stress in bovine mammary epithelial cells and murine mammary glands. Vet. Res. 2023, 54, 78. [Google Scholar] [CrossRef]
- Li, Y.X.; Jiao, P.; Wang, X.P.; Wang, J.P.; Feng, F.; Bao, B.W.; Dong, Y.W.; Luoreng, Z.M.; Wei, D.W. RNA-seq reveals the role of miR-223 in alleviating inflammation of bovine mammary epithelial cells. Res. Vet. Sci. 2023, 159, 257–266. [Google Scholar] [CrossRef]
- Boulton, A.C.; Rushton, J.; Wathes, C.M. Past trends and future challenges for a sustainable UK dairy industry. R. Agric. Soc. Engl. J. 2011, 172, 1–7. [Google Scholar]
- Shortall, O. Cows eat grass, don’t they? Contrasting sociotechnical imaginaries of the role of grazing in the UK and Irish diary sectors. J. Rural. Stud. 2019, 72, 45–57. [Google Scholar] [CrossRef]
- Trewern, J.; Chenoweth, J.; Christie, I.; Keller, E.; Halevy, S. Are UK retailers well placed to deliver ‘less and better’meat and dairy to consumers? Sustain. Prod. Consum. 2021, 28, 154–163. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of Animal and Marine Origin: Structure, Function, and Anti-Inflammatory Properties. Molecules 2017, 22, 1964. [Google Scholar] [CrossRef]
- Scherer, L.; Behrens, P.; Tukker, A. Opportunity for a dietary win-win-win in nutrition, environment, and animal welfare. One Earth 2019, 1, 349–360. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Small EV | Large EV | |||||
|---|---|---|---|---|---|---|
| Total RNA (ng/µL) | Total Protein (µg/µL) | VesiDye-Positive EV/µL | Total RNA (ng/µL) | Total Protein (µg/µL) | VesiDye-Positive EV/µL | |
| SMEV1 | 22.1 | 145.58 | 18,411.52 | 59.053 | 1675.15 | 90,541.44 |
| SMEV2 | 52.0 | 140.23 | 26,384.11 | 159.075 | 1915.58 | 39,303.23 |
| SMEV3 | 23.0 | 153.25 | 36,564.71 | 79.977 | 1915.85 | 17,200.00 |
| SMEV4 | 100.3 | 127.4 | 32,495.47 | 92.803 | 1414.13 | 13,4833.3 |
| SMEV5 | 49.4 | 197.95 | 23,083.8 | 54.648 | 1810.05 | 56,615.38 |
| SMEV6 | 39.0 | 136.35 | 30,837.61 | 54.485 | 1618.4 | 18,077.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simbi, B.; Pink, R.C.; Whatford, L.; Lawson, C. Particles in Raw Sheep Milk Can Modulate the Inflammatory Response in THP-1, a Human Monocyte Cell Line, In Vitro. Dairy 2024, 5, 161-172. https://doi.org/10.3390/dairy5010013
Simbi B, Pink RC, Whatford L, Lawson C. Particles in Raw Sheep Milk Can Modulate the Inflammatory Response in THP-1, a Human Monocyte Cell Line, In Vitro. Dairy. 2024; 5(1):161-172. https://doi.org/10.3390/dairy5010013
Chicago/Turabian StyleSimbi, Bigboy, Ryan C. Pink, Louise Whatford, and Charlotte Lawson. 2024. "Particles in Raw Sheep Milk Can Modulate the Inflammatory Response in THP-1, a Human Monocyte Cell Line, In Vitro" Dairy 5, no. 1: 161-172. https://doi.org/10.3390/dairy5010013