Effects of Dietary L-Carnitine Supplementation on Platelets and Erythrogram of Dairy Cows with Special Emphasis on Parturition

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment and Design
2.2. Sample Collection
2.3. Laboratory Methods
2.3.1. Hematology
2.3.2. Antioxidant Enzyme Activities
2.3.3. Ferric Reducing Ability of Plasma (FRAP)
2.3.4. Derivatives of Reactive Oxygen Metabolites (dROM)
2.4. Statistical Analysis
3. Results
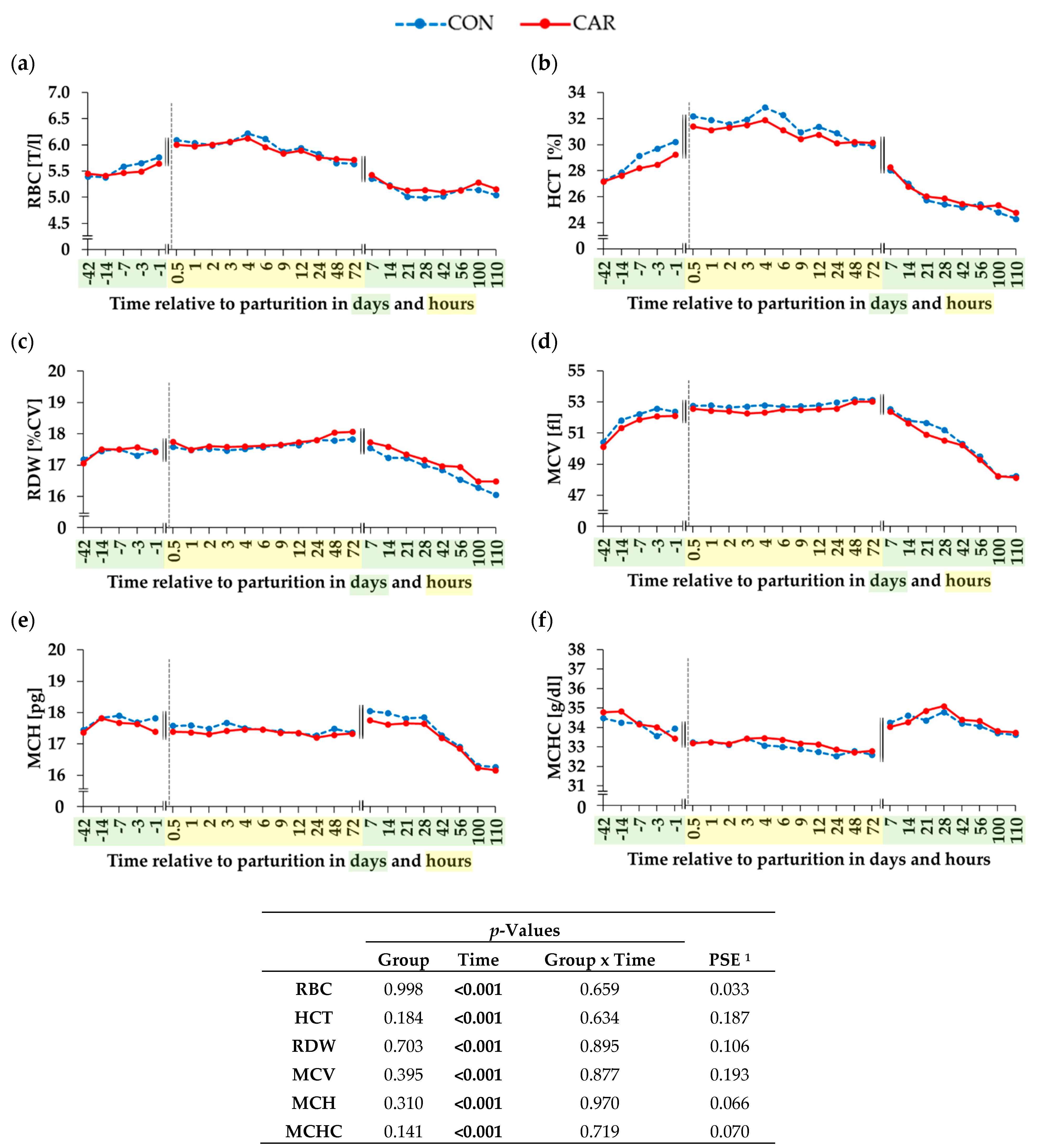
3.1. Erythrogram
3.2. Platelets
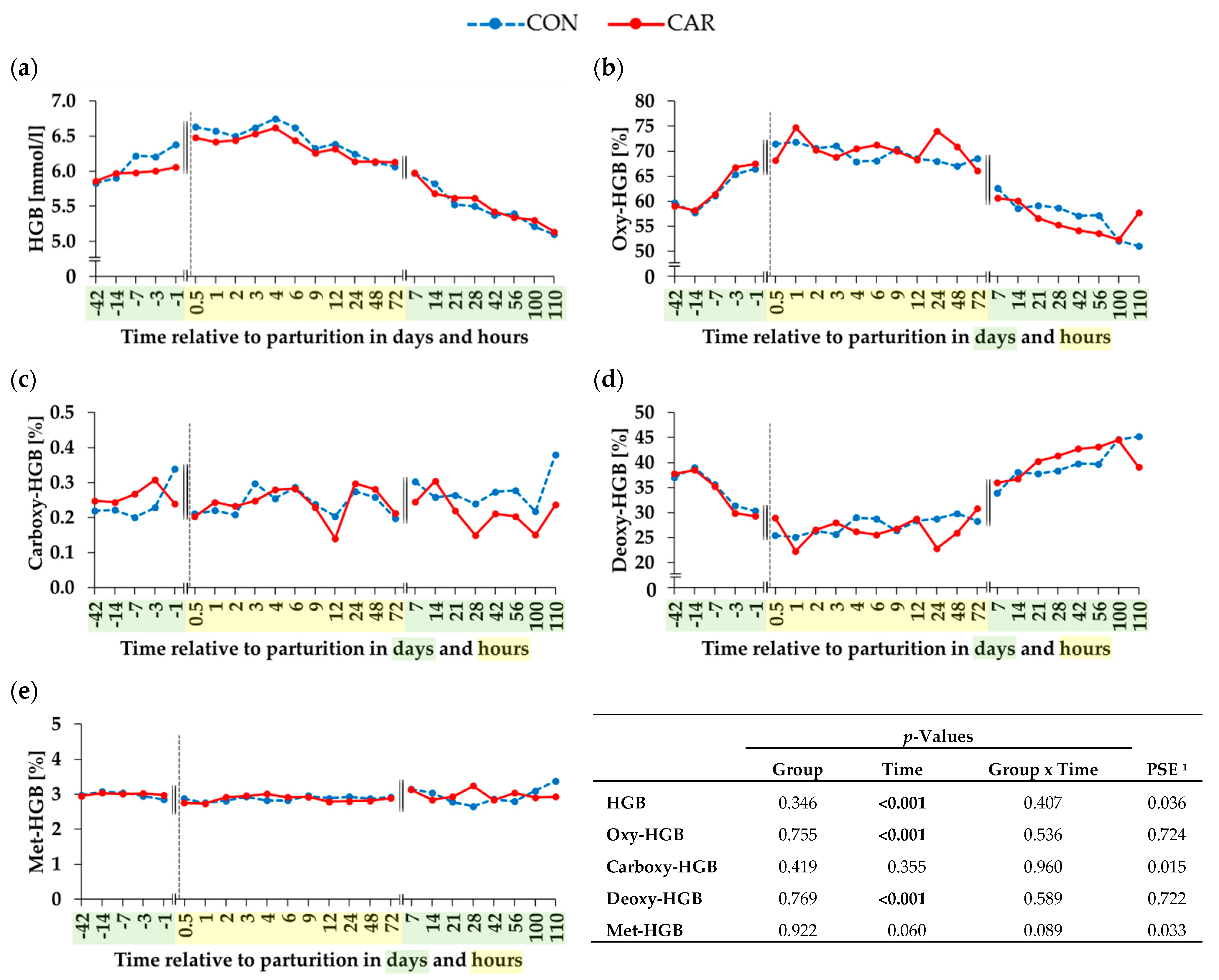
3.3. Hemoglobin and Derivatives of Hemoglobin
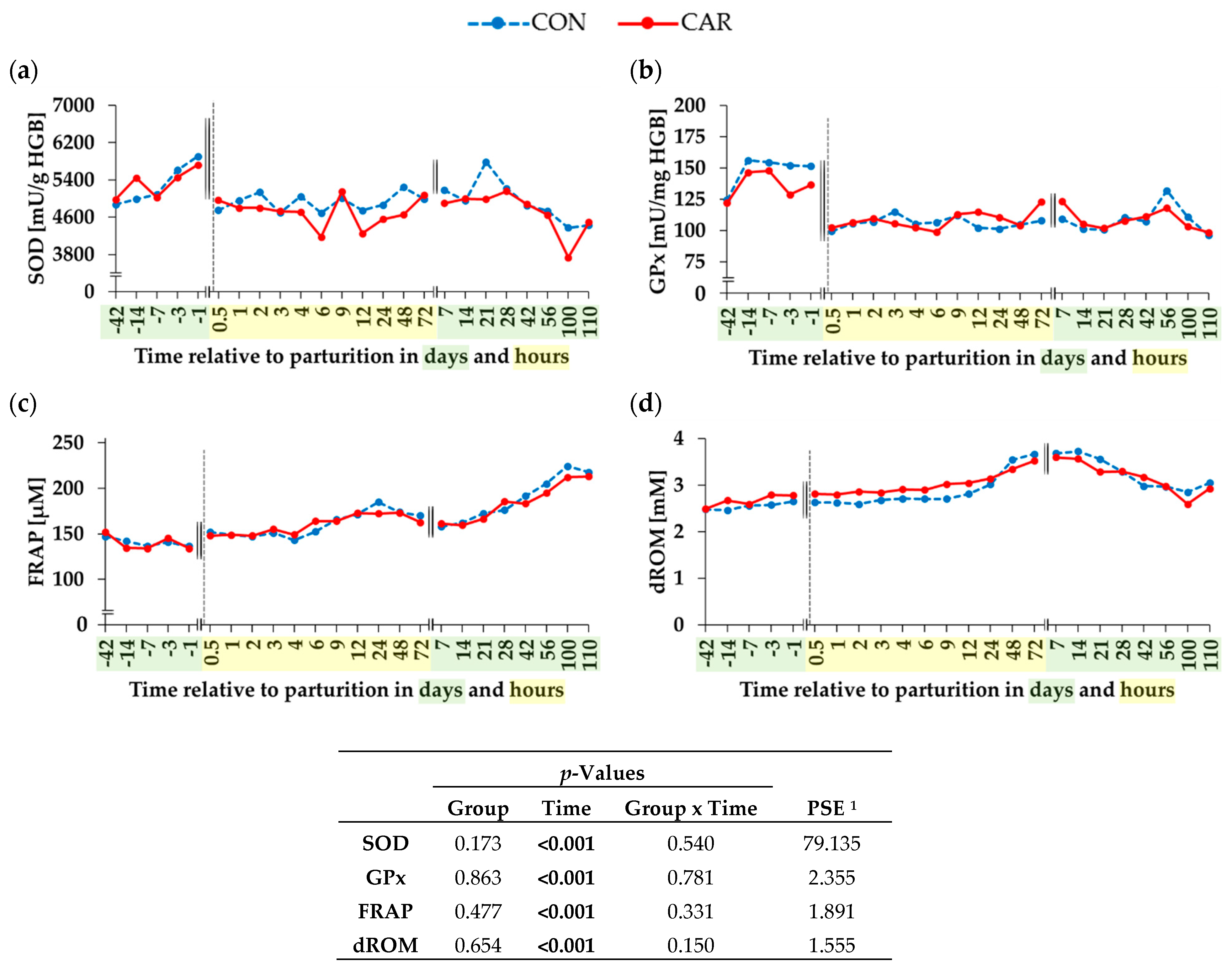
3.4. Indicators for the Oxidative and Antioxidative Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Găvan, C.; Retea, C.; Motorga, V. Changes in the hematological profile of Holstein primiparous in periparturient period and in early to mid lactation. Sci. Pap. Anim. Sci. Biotechnol. 2010, 43, 244–246. [Google Scholar]
- Konvičná, J.; Vargová, M.; Paulíková, I.; Kovcáč, G.; Kostecká, Z. Oxidative stress and antioxidant status in dairy cows during prepartal and postpartal periods. Acta Vet. Brno 2015, 84, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Drackley, J.K.; Overton, T.R.; Douglas, G.N. Adaptations of glucose and long-chain fatty acid metabolism in liver of dairy cows during the periparturient period. J. Dairy Sci. 2001, 84, E100–E112. [Google Scholar] [CrossRef]
- Jonsson, N.N.; Fortes, M.R.S.; Piper, E.K.; Vankan, D.M.; de Cisneros, J.P.J.; Wittek, T. Comparison of metabolic, hematological, and peripheral blood leukocyte cytokine profiles of dairy cows and heifers during the periparturient period. J. Dairy Sci. 2013, 96, 2283–2292. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.B.; Forte, C.A.; Jones, D. Carnitine and acetylcarnitine in red blood cells. Biochim. Et Biophys. Acta Lipids 1988, 959, 100–105. [Google Scholar] [CrossRef]
- Arduini, A.; Rossi, M.; Mancinelli, G.; Belfiglio, M.; Scurti, R.; Radatti, G.; Shohet, S.B. Effect of L-carnitine and acetyl-L-carnitibe on the human erythrocyte membrane stability and deformability. Life Sci. 1990, 47, 2395–2400. [Google Scholar] [CrossRef]
- Kraft, W.; Dürr, U.M. Klinische Labordiagnostik in der Tiermedizin; Schattauer: Stuttgart, Germany, 2005. [Google Scholar]
- Budak, Y.U.; Polat, M.; Huysal, K. The use of platelet indices, plateletcrit, mean platelet volume and platelet distribution width in emergency non-traumatic abdominal surgery: A systematic review. Biochem. Med. 2016, 26, 178–193. [Google Scholar] [CrossRef]
- Jenne, C.N.; Urrutia, R.; Kubes, P. Platelets: Bridging hemostasis, inflammation, and immunity. Int. J. Lab. Hematol. 2013, 35, 254–261. [Google Scholar] [CrossRef]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Johnson, M.S.; Hardy, R.W.; Ballinger, S.W.; Darley-Usmar, V.M. Methods for defining distinct bioenergetic profiles in platelets, lymphocytes, monocytes, and neutrophils, and the oxidative burst from human blood. Lab. Investig. 2013, 93, 690–700. [Google Scholar] [CrossRef]
- Iuliano, L.; Colavita, A.R.; Leo, R.; Praticò, D.; Violi, F. Oxygen Free Radicals and Platelet Activation. Free Radic. Biol. Med. 1997, 22, 999–1006. [Google Scholar] [CrossRef]
- Strasser, A.; Dedoyard, A.; Lohninger, A.; Niedermüller, H. l-Carnitine l-tartrate (LCLT) and dehydroepiandrosterone sulfate (DHEAS) affect red and white blood cells in aged Sprague-Dawley rats. Arch. Gerontol. Geriatr. 2007, 44, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, G.; Keller, J.; Hirche, F.; Geissler, S.; Schwarz, F.J.; Ringseis, R.; Stangl, G.I.; Eder, K. Expression of genes involved in hepatic carnitine synthesis and uptake in dairy cows in the transition period and at different stages of lactation. BMC Vet. Res. 2012, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringseis, R.; Keller, J.; Eder, K. Regulation of carnitine status in ruminants and efficacy of carnitine supplementation on performance and health aspects of ruminant livestock: A review. Arch. Anim. Nutr. 2018, 72, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant and antiradical activities of L-carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef]
- Sweeney, J.D.; Arduini, A. L-Carnitine and Its Possible Role in Red Cell and Platelet Storage. Transfus. Med. Rev. 2004, 18, 58–65. [Google Scholar] [CrossRef]
- Pignatelli, P.; Lenti, L.; Sanguigni, V.; Frati, G.; Simeoni, I.; Gazzaniga, P.P.; Pulcinelli, F.M.; Violi, F. Carnitine inhibits arachidonic acid turnover, platelet function, and oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H41–H48. [Google Scholar] [CrossRef]
- Saluk-Juszczak, J.; Olas, B.; Wachowicz, B.; Glowacki, R.; Bald, E. L-carnitine modulates blood platelet oxidative stress. Cell Biol. Toxicol. 2010, 26, 355–365. [Google Scholar] [CrossRef]
- Uluisik, D.; Keskin, E. The effects of L-carnitine on some hematological parameters in rats fed a cholesterol-rich diet. Biotech. Histochem. 2014, 89, 393–397. [Google Scholar] [CrossRef]
- Goff, J.P.; Stabel, J.R. Decreased Plasma Retinol, α-Tocopherol, and Zinc Concentration During the Periparturient Period: Effect of Milk Fever. J. Dairy Sci. 1990, 73, 3195–3199. [Google Scholar] [CrossRef]
- Gessner, D.K.; Most, E.; Schlegel, G.; Kupczyk, K.; Schwarz, F.J.; Eder, K. Concentrations of retinol and tocopherols in the milk of cows supplemented with conjugated linoleic acid. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.; Daniels, S.U.; Grindler, S.; Tröscher-Mußotter, J.; Alaedin, M.; Frahm, J.; Hüther, L.; Kluess, J.; Kersten, S.; von Soosten, D.; et al. Effects of a Dietary L-Carnitine Supplementation on Performance, Energy Metabolism and Recovery from Calving in Dairy Cows. Animals 2020, 10, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regenhard, P.; Nakov, D.; Sauerwein, H. Applicability of a spectrophotometric method for assessment of oxidative stress in poultry. Maced. Vet. Rev. 2014, 37, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Akers, R.M. A 100-Year Review: Mammary development and lactation. J. Dairy Sci. 2017, 100, 10332–10352. [Google Scholar] [CrossRef] [PubMed]
- Fandrey, J. Oxygen-dependent and tissue-specific regulation of erythropoietin gene expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R977–R988. [Google Scholar] [CrossRef]
- Moritz, K.M.; Lim, G.B.; Wintour, E.M. Developmental regulation of erythropoietin and erythropoiesis. Am. J. Physiol. 1997, 273, R1829–R1844. [Google Scholar] [CrossRef]
- Haase, V.H. Hypoxic regulation of erythropoiesis and iron metabolism. Am. J. Physiol. Ren. Physiol. 2010, 299, F1–F13. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Zhao, F.-Q. Emerging evidence of the physiological role of hypoxia in mammary development and lactation. J. Anim. Sci. Biotechnol. 2014, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Götze, A.; Honnens, A.; Flachowsky, G.; Bollwein, H. Variability of mammary blood flow in lactating Holstein-Friesian cows during the first twelve weeks of lactation. J. Dairy Sci. 2010, 93, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Fleet, I.R.; Peaker, M. Mammary function and its control at the cessation of lactation in the goat. J. Physiol. 1978, 279, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K.; Aikman, P.C.; Lupoli, B.; Humphries, D.J.; Beever, D.E. Splanchnic Metabolism of Dairy Cows During the Transition From Late Gestation Through Early Lactation. J. Dairy Sci. 2003, 86, 1201–1217. [Google Scholar] [CrossRef]
- Moretti, P.; Probo, M.; Cantoni, A.; Paltrinieri, S.; Giordano, A. Fluctuation of neutrophil counts around parturition in Holstein dairy cows with and without retained placenta. Res. Vet. Sci. 2016, 107, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Tharwat, M.; Ali, A.; Al-Sobayil, F. Hematological and biochemical profiles in goats during the transition period. Comp. Clin. Pathol. 2015, 24, 1–7. [Google Scholar] [CrossRef]
- Olmos, G.; Boyle, L.; Horan, B.; Berry, D.P.; Sayers, R.; Hanlon, A.; Mee, J.F. Effect of holstein-friesian genetic group on peripartum and early lactation haematological and acute phase proteins profiles, health and fertility. Animal 2009, 3, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belić, B.; Cincović, M.R.; Davidov, I.; Lako, B.; Potkonjak, A.; Stančić, I. Periparturient hematological finding in dairy cows with uterus and udder inflammation. Serb. J. Agric. Sci. 2012, 61, 111. [Google Scholar]
- Schäfers, S.; von Soosten, D.; Meyer, U.; Drong, C.; Frahm, J.; Tröscher, A.; Pelletier, W.; Sauerwein, H.; Dänicke, S. Influence of conjugated linoleic acids and vitamin E on biochemical, hematological, and immunological variables of dairy cows during the transition period. J. Dairy Sci. 2018, 101, 1585–1600. [Google Scholar] [CrossRef] [Green Version]
- Klinkon, M.; Zadnik, T. Dynamics of red and white blood picture in dairy cows during the periparturient period. Comp. Haematol. Int. 1999, 9, 156–161. [Google Scholar] [CrossRef]
- Chandra, S.; Tripathi, A.K.; Mishra, S.; Amzarul, M.; Vaish, A.K. Physiological changes in hematological parameters during pregnancy. Indian J. Hematol. Blood Transfus. 2012, 28, 144–146. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kumar, P.; Singh, M.; Vasishta, N.K. Haemato-biochemical and endocrine profiling of north western Himalayan Gaddi sheep during various physiological/reproductive phases. Open Vet. J. 2015, 5, 103–107. [Google Scholar] [PubMed]
- Abugo, O.O.; Rifkind, J.M. Oxidation of hemoglobin and the enhancement produced by nitroblue tetrazolium. J. Biol. Chem. 1994, 269, 24845–24853. [Google Scholar] [PubMed]
- Nagababu, E.; Gulyani, S.; Earley, C.J.; Cutler, R.G.; Mattson, M.P.; Rifkind, J.M. Iron-deficiency anaemia enhances red blood cell oxidative stress. Free Radic. Res. 2008, 42, 824–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Influence of Body Condition Score on Relationships Between Metabolic Status and Oxidative Stress in Periparturient Dairy Cows. J. Dairy Sci. 2005, 88, 2017–2026. [Google Scholar] [CrossRef] [Green Version]
- Bühler, S.; Frahm, J.; Tienken, R.; Kersten, S.; Meyer, U.; Huber, K.; Dänicke, S. Effects of energy supply and nicotinic acid supplementation on serum anti-oxidative capacity and on expression of oxidative stress-related genes in blood leucocytes of periparturient primi- and pluriparous dairy cows. J. Anim. Physiol. Anim. Nutr. 2018, 102, e87–e98. [Google Scholar] [CrossRef] [Green Version]
- Gaál, T.; Ribiczeyné-Szabó, P.; Stadler, K.; Jakus, J.; Reiczigel, J.; Kövér, P.; Mézes, M.; Sümeghy, L. Free radicals, lipid peroxidation and the antioxidant system in the blood of cows and newborn calves around calving. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 143, 391–396. [Google Scholar] [CrossRef]
- Urh, C.; Denißen, J.; Gerster, E.; Kraus, N.; Stamer, E.; Heitkönig, B.; Spiekers, H.; Sauerwein, H. Pro-and antioxidative indicators in serum of dairy cows during late pregnancy and early lactation: Testing the effects of parity, different dietary energy levels, and farm. J. Dairy Sci. 2019, 102, 6672–6678. [Google Scholar] [CrossRef]
- Li, J.-L.; Wang, Q.-Y.; Luan, H.-Y.; Kang, Z.-C.; Wang, C.-B. Effects of L-carnitine against oxidative stress in human hepatocytes: Involvement of peroxisome proliferator-activated receptor alpha. J. Biomed. Sci. 2012, 19, 32. [Google Scholar] [CrossRef] [Green Version]
- Binienda, Z.K.; Ali, S.F. Neuroprotective role of L-carnitine in the 3-nitropropionic acid induced neurotoxicity. Toxicol. Lett. 2001, 125, 67–73. [Google Scholar] [CrossRef]
- Lee, B.J.; Lin, J.S.; Lin, Y.C.; Lin, P.T. Effects of L-carnitine supplementation on oxidative stress and antioxidant enzymes activities in patients with coronary artery disease: A randomized, placebo-controlled trial. Nutr. J. 2014, 13, 79. [Google Scholar] [CrossRef] [Green Version]
- Thangasamy, T.; Subathra, M.; Sittadjody, S.; Jeyakumar, P.; Joyee, A.G.; Mendoza, E.; Chinnakkanu, P. Role of L-carnitine in the modulation of immune response in aged rats. Clin. Chim. Acta 2008, 389, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Asadi, H.; Sadeghi, A.; Eila, N.; Aminafshar, M. Carcass traits and immune response of broiler chickens fed dietary L-Carnitine, coenzyme Q10 and ractopamine. Braz. J. Poult. Sci. 2016, 18, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Sotirakopoulos, N.; Athanasiou, G.; Tsitsios, T.; Mavromatidis, K. The influence of l-carnitine supplementation on hematocrit and hemoglobin levels in patients with end stage renal failure on CAPD. Ren. Fail. 2002, 24, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Arduini, A.; Gorbunov, N.; Arrigoni-Martelli, E.; Dottori, S.; Molajoni, F.; Russo, F.; Federici, G. Effects of l-carnitine and its acetate and propionate esters on the molecular dynamics of human erythrocyte membrane. Biochim. Et Biophys. Acta (BBA) Biomembr. 1993, 1146, 229–235. [Google Scholar] [CrossRef]
- Fritz, I.B.; Wong, K.; Burdzy, K. Clustering of erythrocytes by fibrinogen is inhibited by carnitine: Evidence that sulfhydryl groups on red blood cell membranes are involved in carnitine actions. J. Cell. Physiol. 1991, 149, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Orrù, L.; Abeni, F.; Catillo, G.; Grandoni, F.; Crisà, A.; De Matteis, G.; Carmela Scatà, M.; Napolitano, F.; Moioli, B. Leptin gene haplotypes are associated with change in immunological and hematological variables in dairy cow during the peripartum period. J. Anim. Sci. 2012, 90, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Schoene, N.W.; Guidry, C.A. Dietary soy isoflavones inhibit activation of rat platelets. J. Nutr. Biochem. 1999, 10, 421–426. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kononov, S.U.; Meyer, J.; Frahm, J.; Kersten, S.; Kluess, J.; Meyer, U.; Huber, K.; Dänicke, S. Effects of Dietary L-Carnitine Supplementation on Platelets and Erythrogram of Dairy Cows with Special Emphasis on Parturition. Dairy 2021, 2, 1-13. https://doi.org/10.3390/dairy2010001
Kononov SU, Meyer J, Frahm J, Kersten S, Kluess J, Meyer U, Huber K, Dänicke S. Effects of Dietary L-Carnitine Supplementation on Platelets and Erythrogram of Dairy Cows with Special Emphasis on Parturition. Dairy. 2021; 2(1):1-13. https://doi.org/10.3390/dairy2010001
Chicago/Turabian StyleKononov, Susanne Ursula, Jennifer Meyer, Jana Frahm, Susanne Kersten, Jeannette Kluess, Ulrich Meyer, Korinna Huber, and Sven Dänicke. 2021. "Effects of Dietary L-Carnitine Supplementation on Platelets and Erythrogram of Dairy Cows with Special Emphasis on Parturition" Dairy 2, no. 1: 1-13. https://doi.org/10.3390/dairy2010001