
Molecular Dynamics Simulations of Shockwave Affected STMV Virus to Measure the Frequencies of the Oscillatory Response


Abstract
:1. Introduction
2. Materials and Methods
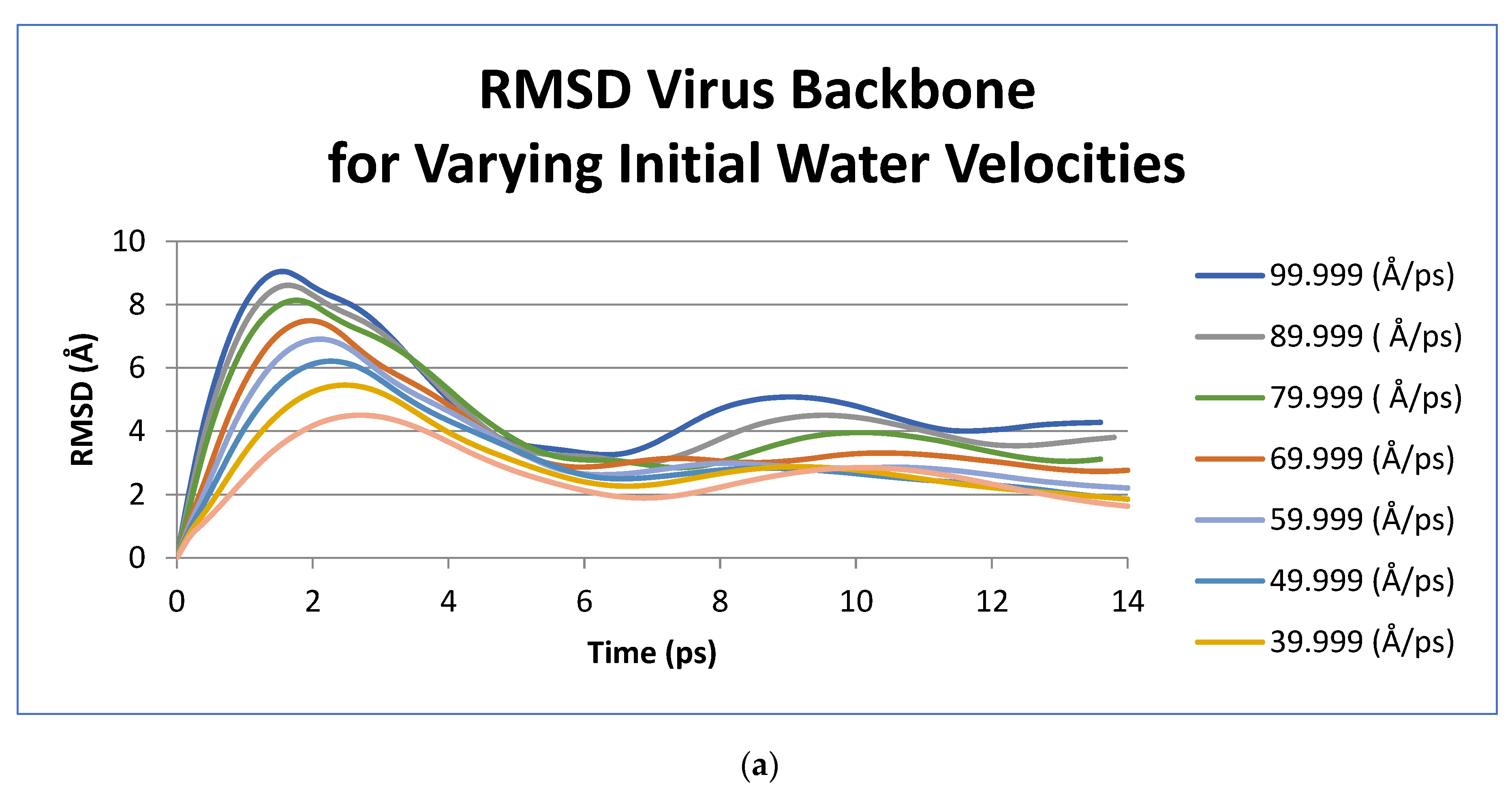
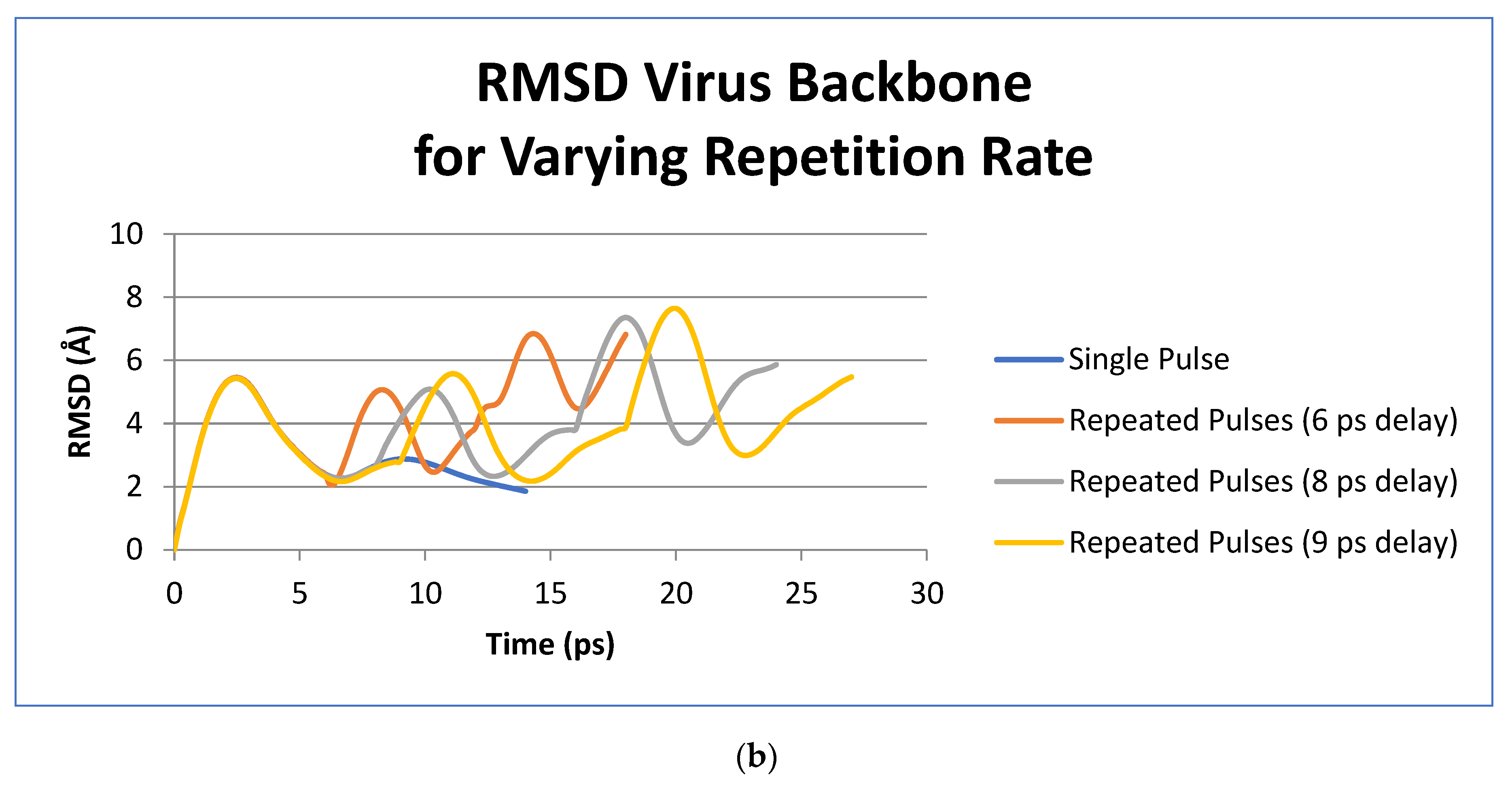
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Spheroidal Mode Number | Frequency (GHz) |
|---|---|
| L0 | 92.47 |
| L1 | 60.60 |
| L2 | 44.67 |
| L3 | 66.57 |
References
- Hamrick, P.E.; Cleary, S.F. Laser-Induced Acoustic Breakage of Tobacco Mosaic Virus. Nature 1968, 220, 909–910. [Google Scholar] [CrossRef]
- Smith, R.J.; Cleary, S.F. Investigation of Structural Bonding Forces in Bacteriophage T2. J. Acoust. Soc. Am. 1974, 56, 1883–1889. [Google Scholar] [CrossRef]
- Tsen, S.-W.D.; Kingsley, D.H.; Poweleit, C.; Achilefu, S.; Soroka, D.S.; Wu, T.C.; Tsen, K.-T. Studies of Inactivation Mechanism of Non-Enveloped Icosahedral Virus by a Visible Ultrashort Pulsed Laser. Virol. J. 2014, 11, 20. [Google Scholar] [CrossRef]
- Nazari, M.; Xi, M.; Lerch, S.; Alizadeh, M.; Ettinger, C.; Akiyama, H.; Gillespie, C.; Gummuluru, S.; Erramilli, S.; Reinhard, B.M. Plasmonic Enhancement of Selective Photonic Virus Inactivation. Sci. Rep. 2017, 7, 11951. [Google Scholar] [CrossRef] [Green Version]
- Babincová, M.; Sourivong, P.; Babinec, P. Resonant absorption of ultrasound energy as a method of HIV destruction. Med. Hypotheses 2000, 55, 450. [Google Scholar] [CrossRef]
- Yang, S.-C.; Lin, H.-C.; Liu, T.-M.; Lu, J.-T.; Hung, W.-T.; Huang, Y.-R.; Tsai, Y.-C.; Kao, C.-L.; Chen, S.-Y.; Sun, C.-K. Efficient Structure Resonance Energy Transfer from Microwaves to Confined Acoustic Vibrations in Viruses. Sci. Rep. 2015, 5, 18030. [Google Scholar] [CrossRef] [Green Version]
- Hempel, E. Irradiation Device for Influencing a Biological Structure in a Subject with Electromagnetic Radiation 2010. U.S. Patent US7648498B2, 19 January 2010. [Google Scholar]
- Ford, L. Estimate of the Vibrational Frequencies of Spherical Virus Particles. Phys. Rev. E 2003, 67, 051924. [Google Scholar] [CrossRef] [Green Version]
- Saviot, L.; Murray, D.B.; Mermet, A.; Duval, E. Comment on “Estimate of the Vibrational Frequencies of Spherical Virus Particles”. Phys. Rev. E 2004, 69, 023901. [Google Scholar] [CrossRef] [Green Version]
- Fonoberov, V.A.; Balandin, A.A. Low-Frequency Vibrational Modes of Viruses Used for Nanoelectronic Self-Assemblies. Phys. Status Solidi (B) 2004, 241, R67–R69. [Google Scholar] [CrossRef]
- Talati, M.; Jha, P.K. Acoustic Phonon Quantization and Low-Frequency Raman Spectra of Spherical Viruses. Phys. Rev. E 2006, 73, 011901. [Google Scholar] [CrossRef]
- Yang, Z.; Bahar, I.; Widom, M. Vibrational Dynamics of Icosahedrally Symmetric Biomolecular Assemblies Compared with Predictions Based on Continuum Elasticity. Biophys. J. 2009, 96, 4438–4448. [Google Scholar] [CrossRef] [Green Version]
- Tama, F.; Brooks III, C.L. Diversity and Identity of Mechanical Properties of Icosahedral Viral Capsids Studied with Elastic Network Normal Mode Analysis. J. Mol. Biol. 2005, 345, 299–314. [Google Scholar] [CrossRef]
- Dykeman, E.C.; Sankey, O.F. Normal Mode Analysis and Applications in Biological Physics. J. Phys. Condens. Matter 2010, 22, 423202. [Google Scholar] [CrossRef]
- Lee, B.H.; Jo, S.; Choi, M.; Kim, M.H.; Choi, J.B.; Kim, M.K. Normal Mode Analysis of Zika Virus. Comput. Biol. Chem. 2018, 72, 53–61. [Google Scholar] [CrossRef]
- Dykeman, E.C.; Sankey, O.F. Atomistic Modeling of the Low-Frequency Mechanical Modes and Raman Spectra of Icosahedral Virus Capsids. Phys. Rev. E 2010, 81, 021918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsen, K.-T.; Dykeman, E.C.; Sankey, O.F.; Tsen, S.-W.D.; Lin, N.-T.; Kiang, J.G. Raman Scattering Studies of the Low-Frequency Vibrational Modes of Bacteriophage M13 in Water—Observation of an Axial Torsion Mode. Nanotechnology 2006, 17, 5474. [Google Scholar] [CrossRef]
- Stephanidis, B.; Adichtchev, S.; Gouet, P.; McPherson, A.; Mermet, A. Elastic Properties of Viruses. Biophys. J. 2007, 93, 1354–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartschuh, R.; Wargacki, S.; Xiong, H.; Neiswinger, J.; Kisliuk, A.; Sihn, S.; Ward, V.; Vaia, R.; Sokolov, A. How Rigid Are Viruses. Phys. Rev. E 2008, 78, 021907. [Google Scholar] [CrossRef]
- Tcherniega, N.; Pershin, S.; Bunkin, A.; Donchenko, E.; Karpova, O.; Kudryavtseva, A.; Lednev, V.; Mironova, T.; Shevchenko, M.; Strokov, M.; et al. Laser Excitation of Gigahertz Vibrations in Cauliflower Mosaic Viruses’ Suspension. Laser Phys. Lett. 2018, 15, 095603. [Google Scholar] [CrossRef] [Green Version]
- Sirotkin, S.; Mermet, A.; Bergoin, M.; Ward, V.; Van Etten, J.L. Viruses as Nanoparticles: Structure versus Collective Dynamics. Phys. Rev. E 2014, 90, 022718. [Google Scholar] [CrossRef] [Green Version]
- Burkhartsmeyer, J.; Wang, Y.; Wong, K.S.; Gordon, R. Optical Trapping, Sizing, and Probing Acoustic Modes of a Small Virus. Appl. Sci. 2020, 10, 394. [Google Scholar] [CrossRef] [Green Version]
- Mattarelli, M.; Vassalli, M.; Caponi, S. Relevant Length Scales in Brillouin Imaging of Biomaterials: The Interplay between Phonons Propagation and Light Focalization. ACS Photonics 2020, 7, 2319–2328. [Google Scholar] [CrossRef]
- Duval, E. Far-Infrared and Raman Vibrational Transitions of a Solid Sphere: Selection Rules. Phys. Rev. B 1992, 46, 5795. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Larson, S.B.; Heitsch, C.E.; McPherson, A.; Harvey, S.C. A Model for the Structure of Satellite Tobacco Mosaic Virus. J. Struct. Biol. 2012, 180, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freddolino, P.L.; Arkhipov, A.S.; Larson, S.B.; McPherson, A.; Schulten, K. Molecular Dynamics Simulations of the Complete Satellite Tobacco Mosaic Virus. Structure 2006, 14, 437–449. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Mori, T.; Kobayashi, C.; Matsunaga, Y.; Yoda, T.; Feig, M.; Sugita, Y. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2015, 5, 310. [Google Scholar] [CrossRef]
- Kumar, S.; Sun, Y.; Kalé, L.V. Acceleration of an Asynchronous Message Driven Programming Paradigm on IBM Blue Gene/Q. In Proceedings of the 2013 IEEE 27th International Symposium on Parallel and Distributed Processing, Cambridge, MA, USA, 20–24 May 2013; IEEE: Piscataway, NJ, USA, 2013; pp. 689–699. [Google Scholar]
- Solov’yov, A.V.; Surdutovich, E.; Scifoni, E.; Mishustin, I.; Greiner, W. Physics of Ion Beam Cancer Therapy: A Multiscale Approach. Phys. Rev. E 2009, 79, 011909. [Google Scholar] [CrossRef] [Green Version]
- Surdutovich, E.; Yakubovich, A.V.; Solov’yov, A.V. Biodamage via Shock Waves Initiated by Irradiation with Ions. Sci. Rep. 2013, 3, 1289. [Google Scholar] [CrossRef] [Green Version]
- Surdutovich, E.; Solov’yov, A. Multiscale Physics of Ion-Beam Cancer Therapy. In Proceedings of the Journal of Physics: Conference Series; IOP Publishing: Bristol, UK, 2012; Volume 373, p. 012001. [Google Scholar]
- Adhikari, U.; Goliaei, A.; Berkowitz, M.L. Mechanism of Membrane Poration by Shock Wave Induced Nanobubble Collapse: A Molecular Dynamics Study. J. Phys. Chem. B 2015, 119, 6225–6234. [Google Scholar] [CrossRef]
- Koshiyama, K.; Kodama, T.; Yano, T.; Fujikawa, S. Molecular Dynamics Simulation of Structural Changes of Lipid Bilayers Induced by Shock Waves: Effects of Incident Angles. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable Molecular Dynamics on CPU and GPU Architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Holian, B.L.; Lomdahl, P.S. Plasticity Induced by Shock Waves in Nonequilibrium Molecular-Dynamics Simulations. Science 1998, 280, 2085–2088. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD–Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Lamb, H. On the Vibrations of an Elastic Sphere. Proc. Lond. Math. Soc. 1881, 1, 189–212. [Google Scholar] [CrossRef] [Green Version]
- Murray, D.B.; Saviot, L. Phonons in an Inhomogeneous Continuum: Vibrations of an Embedded Nanoparticle. Phys. Rev. B 2004, 69, 094305. [Google Scholar] [CrossRef] [Green Version]
- MacKerell Jr, A.D.; Bashford, D.; Bellott, M.; Dunbrack Jr, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]



| Initial Velocity | Period of Oscillation (ps) | Frequency (GHz) |
|---|---|---|
| Water Slice (Å/ps) | ||
| 99.999 | 15.2 | 65.79 |
| 89.999 | 16 | 62.5 |
| 79.999 | 16.4 | 60.98 |
| 69.999 | 16.4 | 60.98 |
| 59.999 | 16.8 | 59.52 |
| 49.999 | 14 | 71.43 |
| 39.999 | 14 | 71.43 |
| 29.999 | 14.8 | 67.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burkhartsmeyer, J.; Wong, K.S. Molecular Dynamics Simulations of Shockwave Affected STMV Virus to Measure the Frequencies of the Oscillatory Response. Acoustics 2022, 4, 268-275. https://doi.org/10.3390/acoustics4010016
Burkhartsmeyer J, Wong KS. Molecular Dynamics Simulations of Shockwave Affected STMV Virus to Measure the Frequencies of the Oscillatory Response. Acoustics. 2022; 4(1):268-275. https://doi.org/10.3390/acoustics4010016
Chicago/Turabian StyleBurkhartsmeyer, Jeffrey, and Kam Sing Wong. 2022. "Molecular Dynamics Simulations of Shockwave Affected STMV Virus to Measure the Frequencies of the Oscillatory Response" Acoustics 4, no. 1: 268-275. https://doi.org/10.3390/acoustics4010016