
The Wave Model of Sleep Dynamics and an Invariant Relationship between NonREM and REM Sleep


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Modeling Sleep–Wake Homeostasis through Interacting Waves
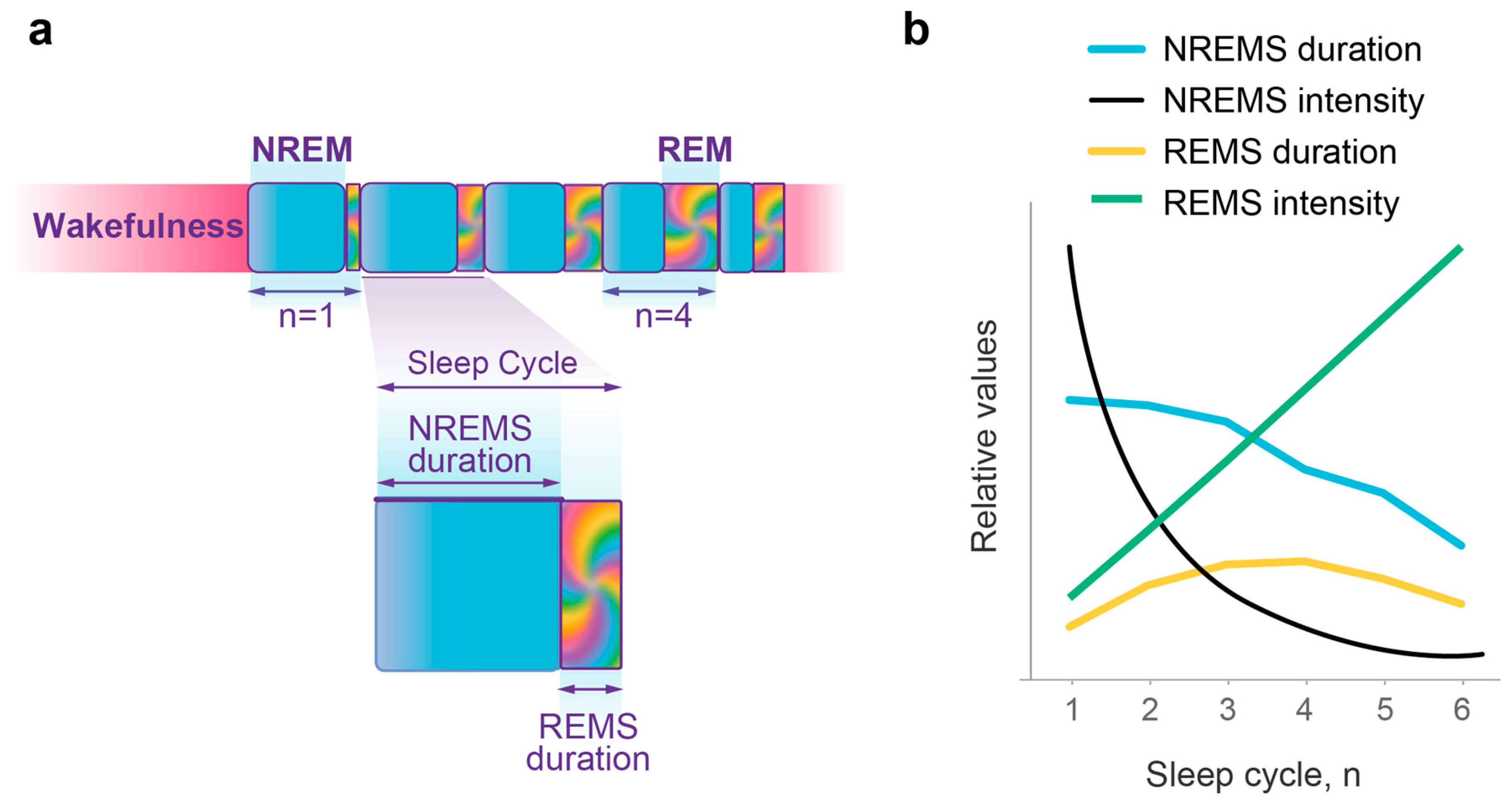
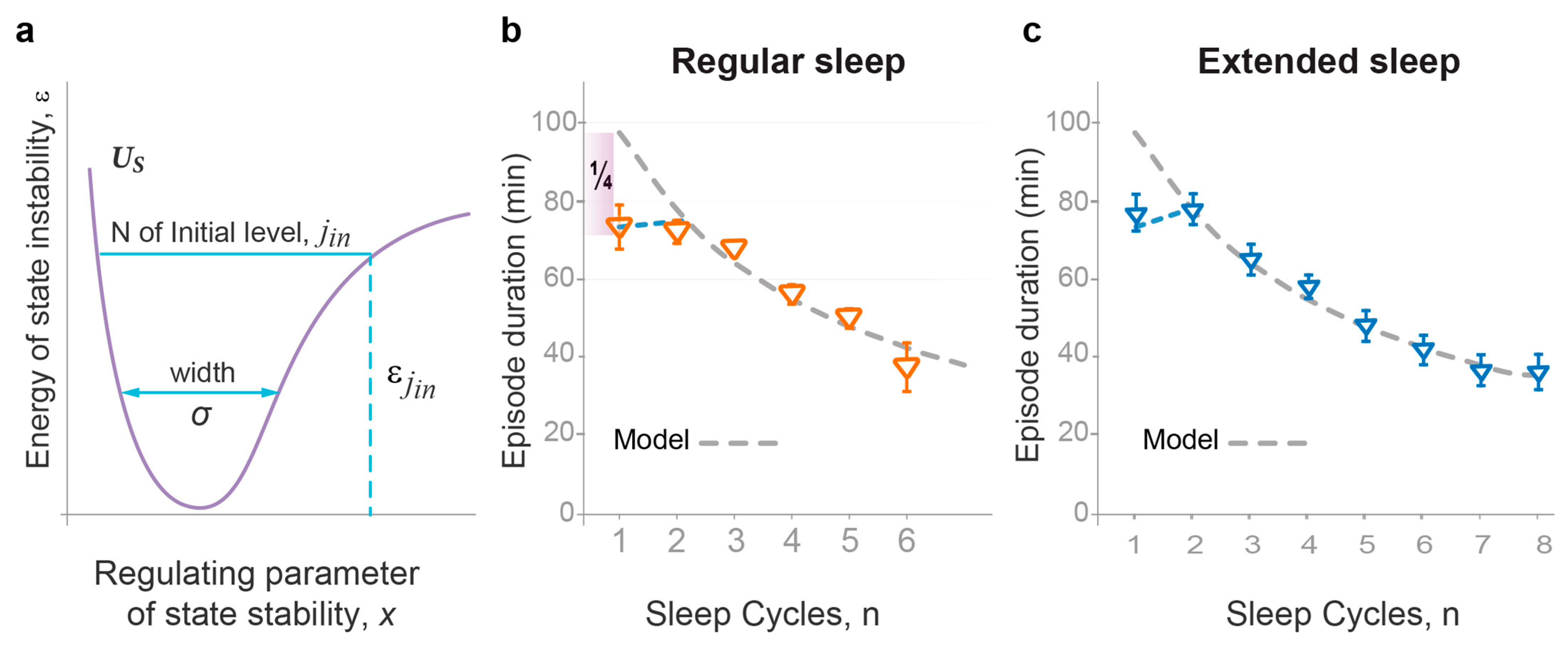
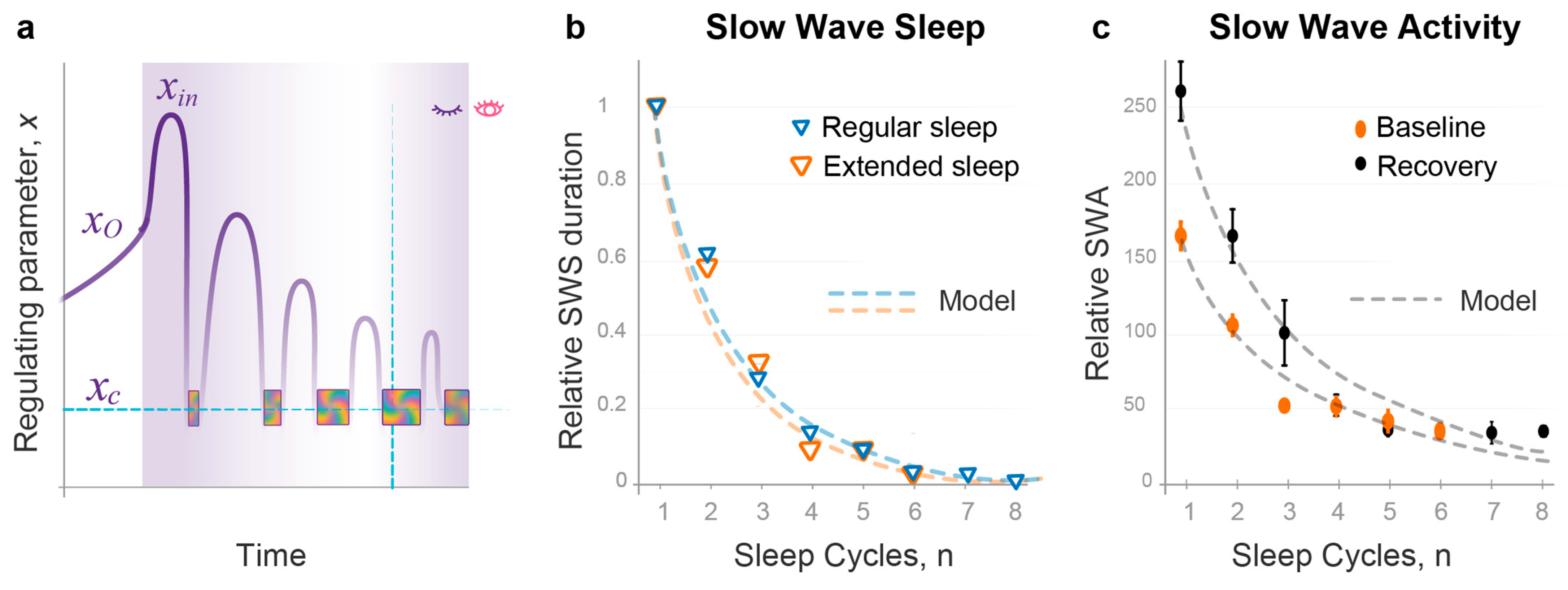
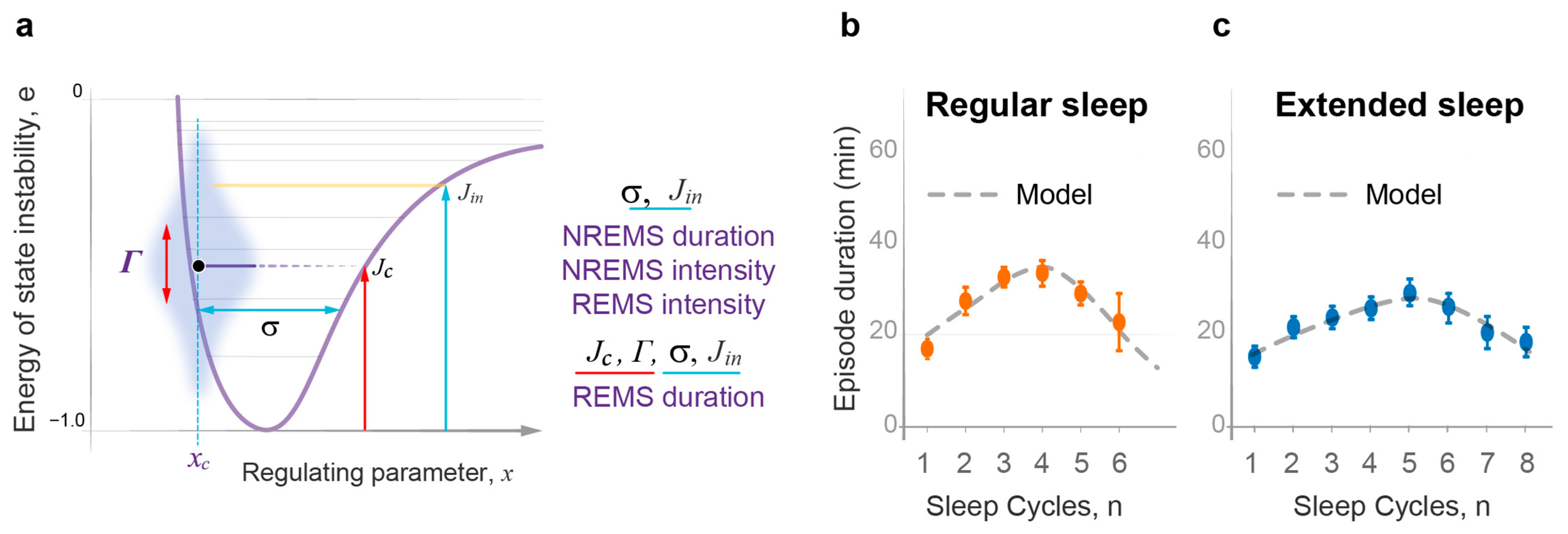
2.2. The Wave Model Accurately Describes the Dynamics of NREMS Episode Duration
2.3. Quantitative Predictions of the Dynamics of NREMS Intensity
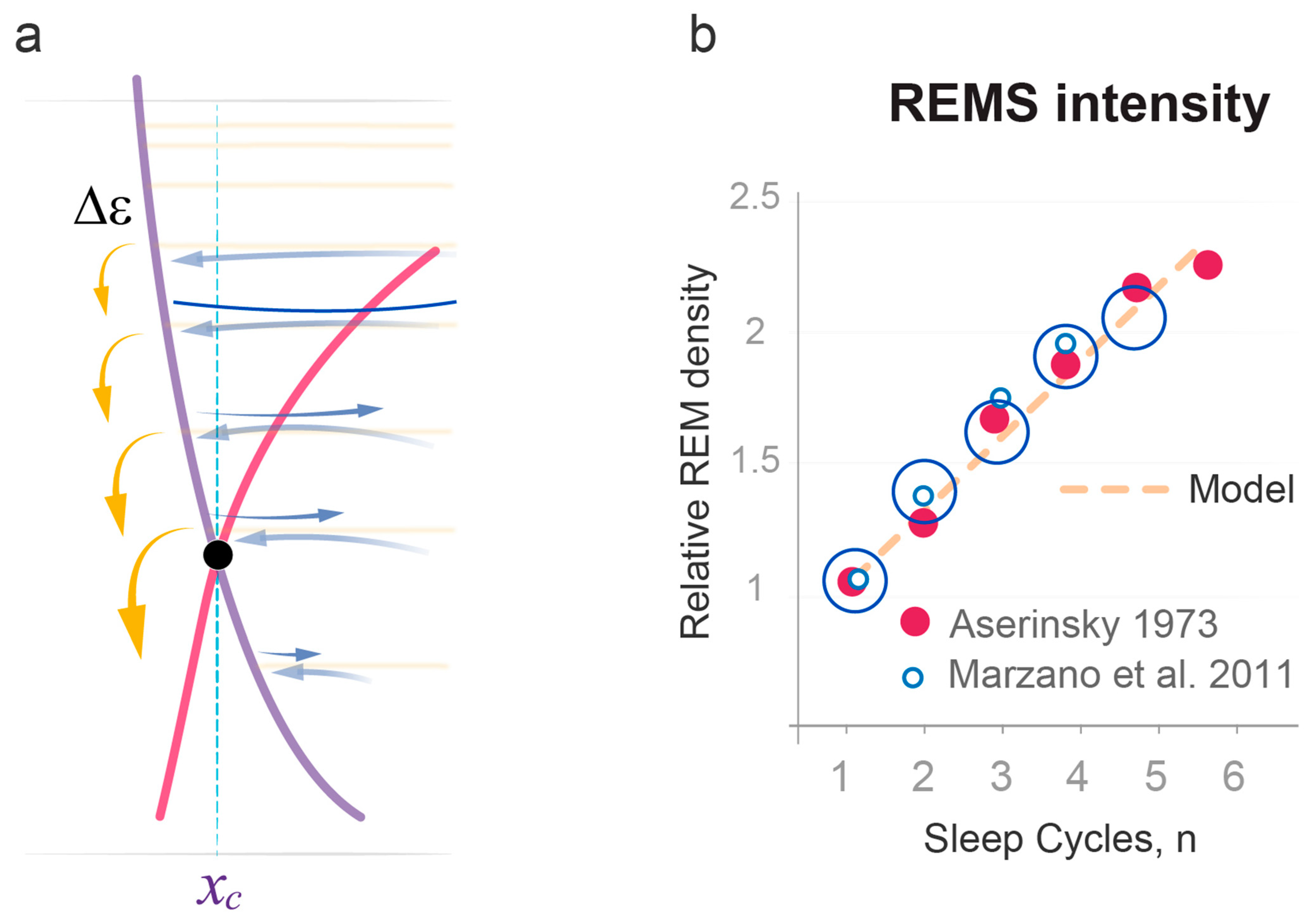
2.4. Quantitative Predictions of the Dynamics of REMS Intensity
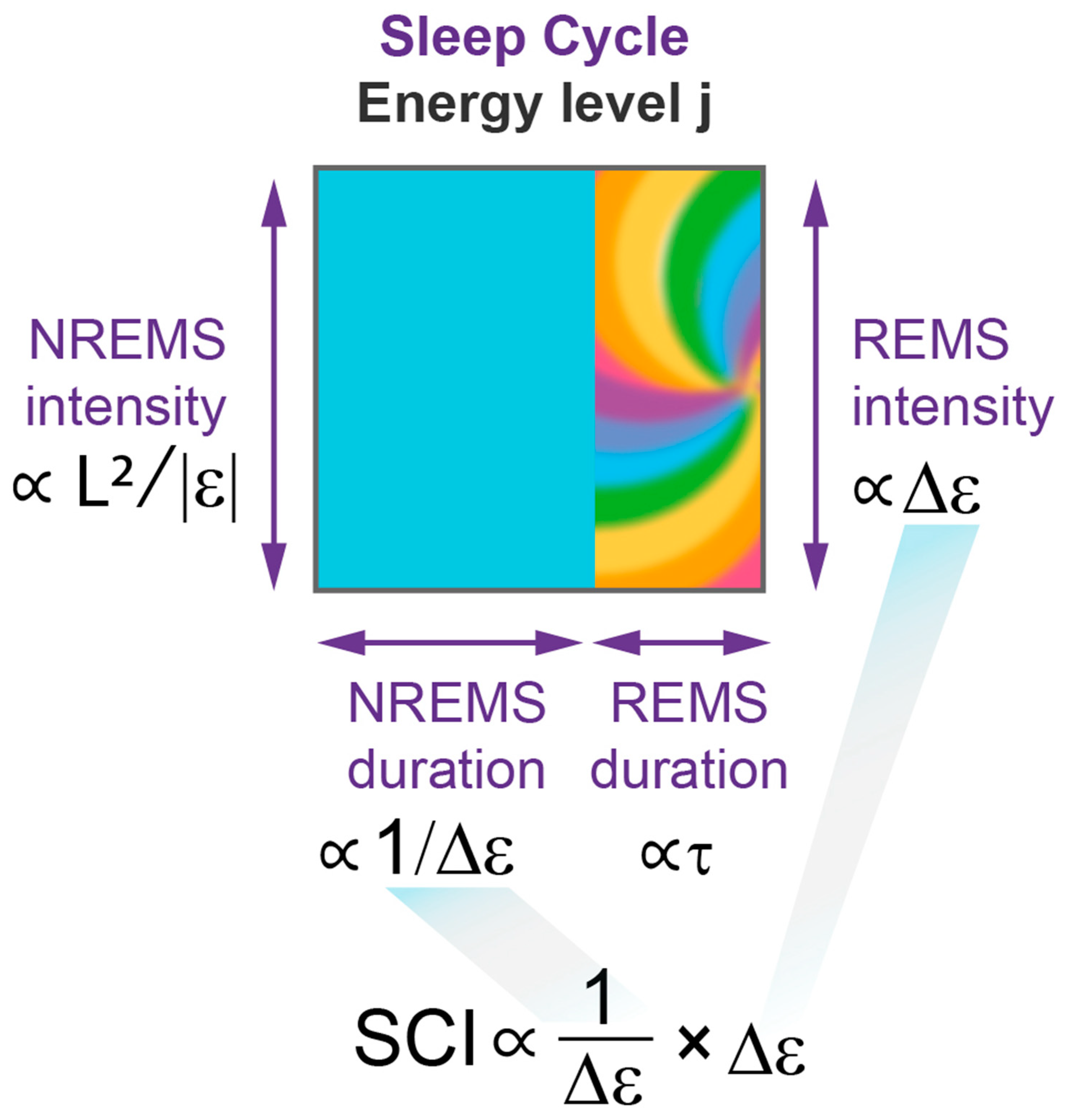
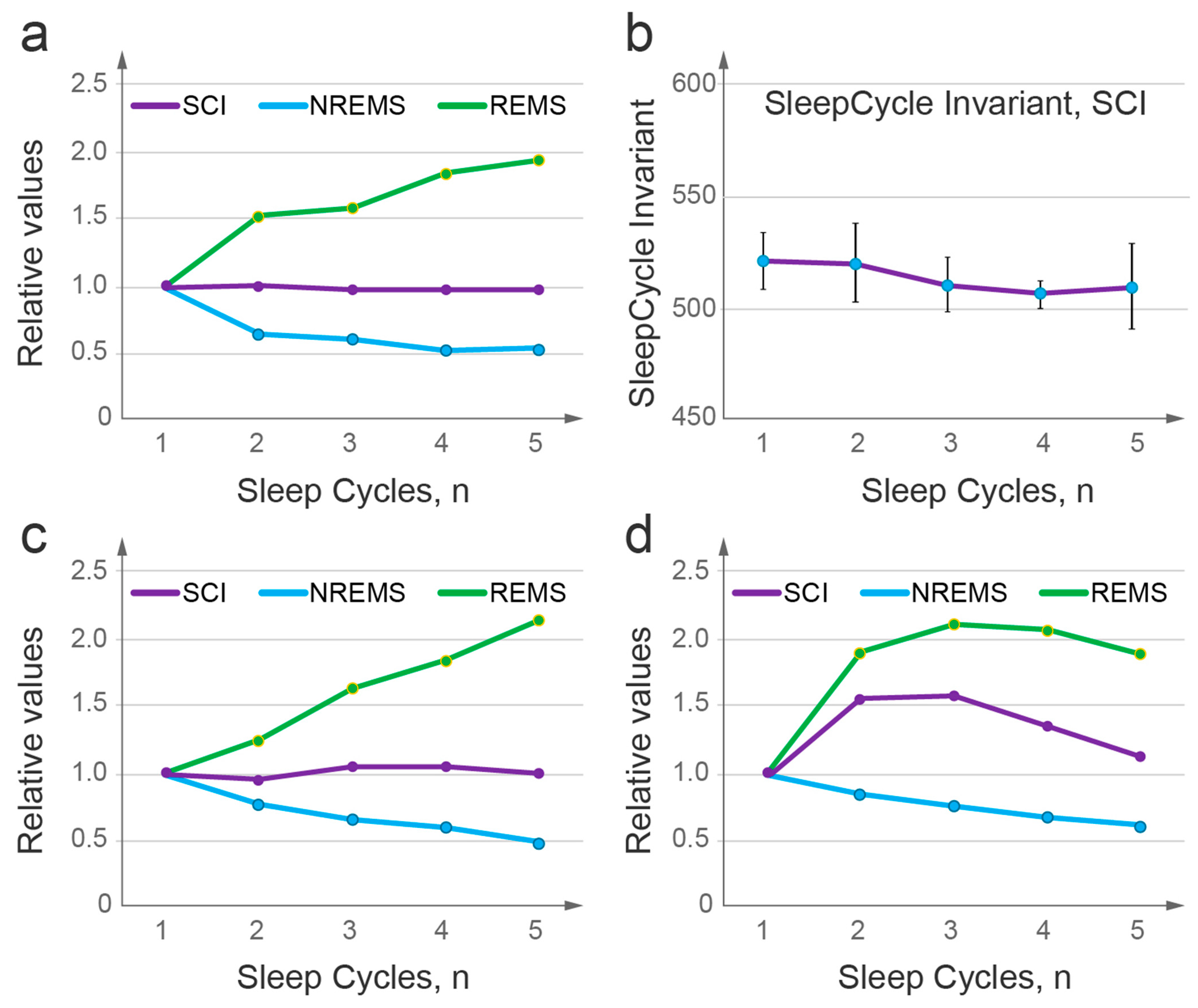
2.5. Sleep Cycle Invariant: Theoretical Prediction and Experimental Demonstration
2.6. Quantitative Predictions of the Dynamics of REMS Duration
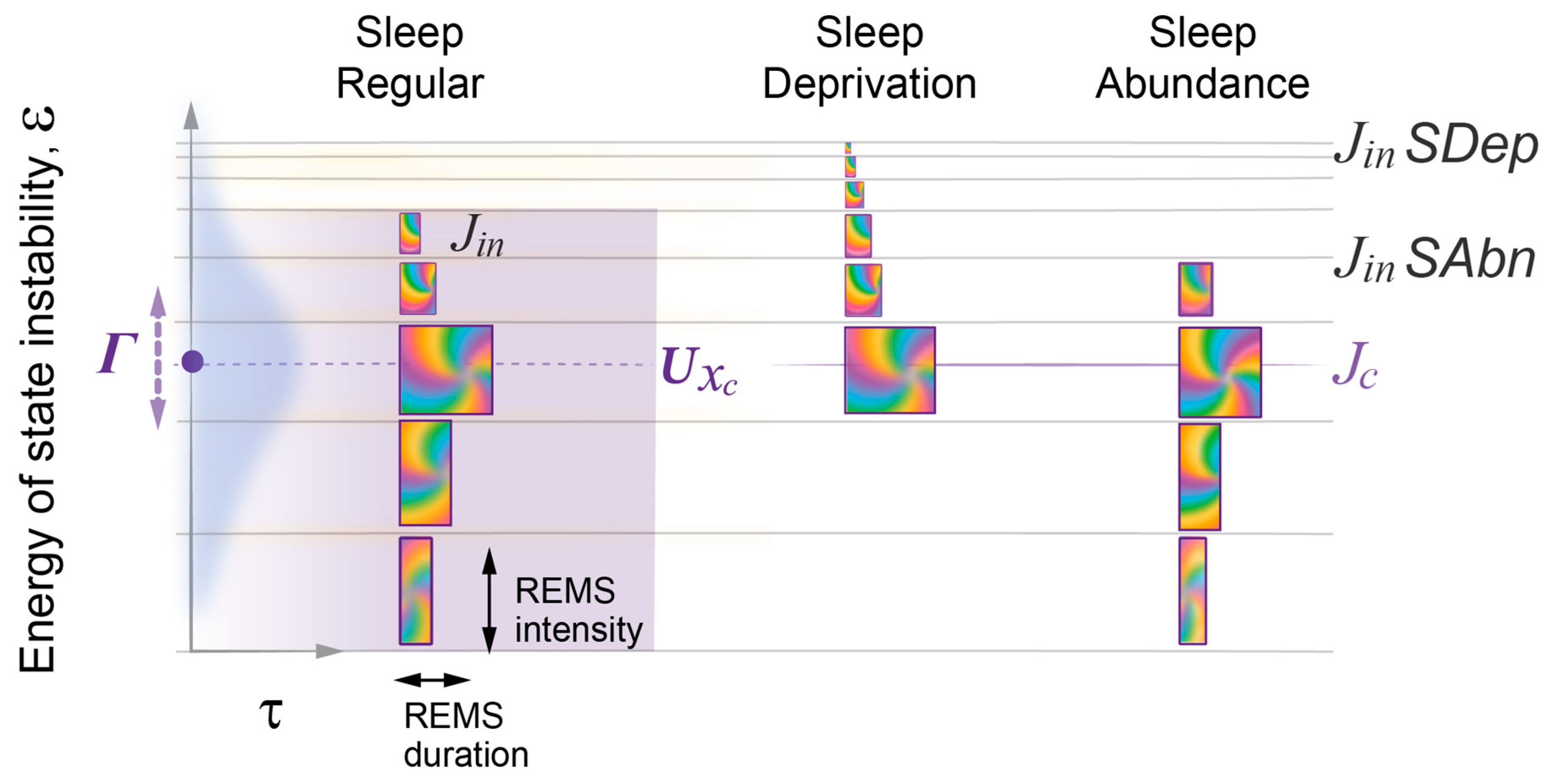
2.7. Model Predicts the Effects of Sleep Deprivation or Abundance on Sleep Architecture
3. Discussion
3.1. Interaction of Two Probability Waves Defines Dynamic Sleep Architecture
3.2. REM Sleep Intensity and the Role of Sleep Cycle Invariant
3.3. State Instability and Strength of Interaction Define the Sleep–Wake Cycle
3.4. Future Directions
4. Methods
4.1. Analogies between the Wave Model of Sleep and the Model of a Diatomic Molecule
- (a)
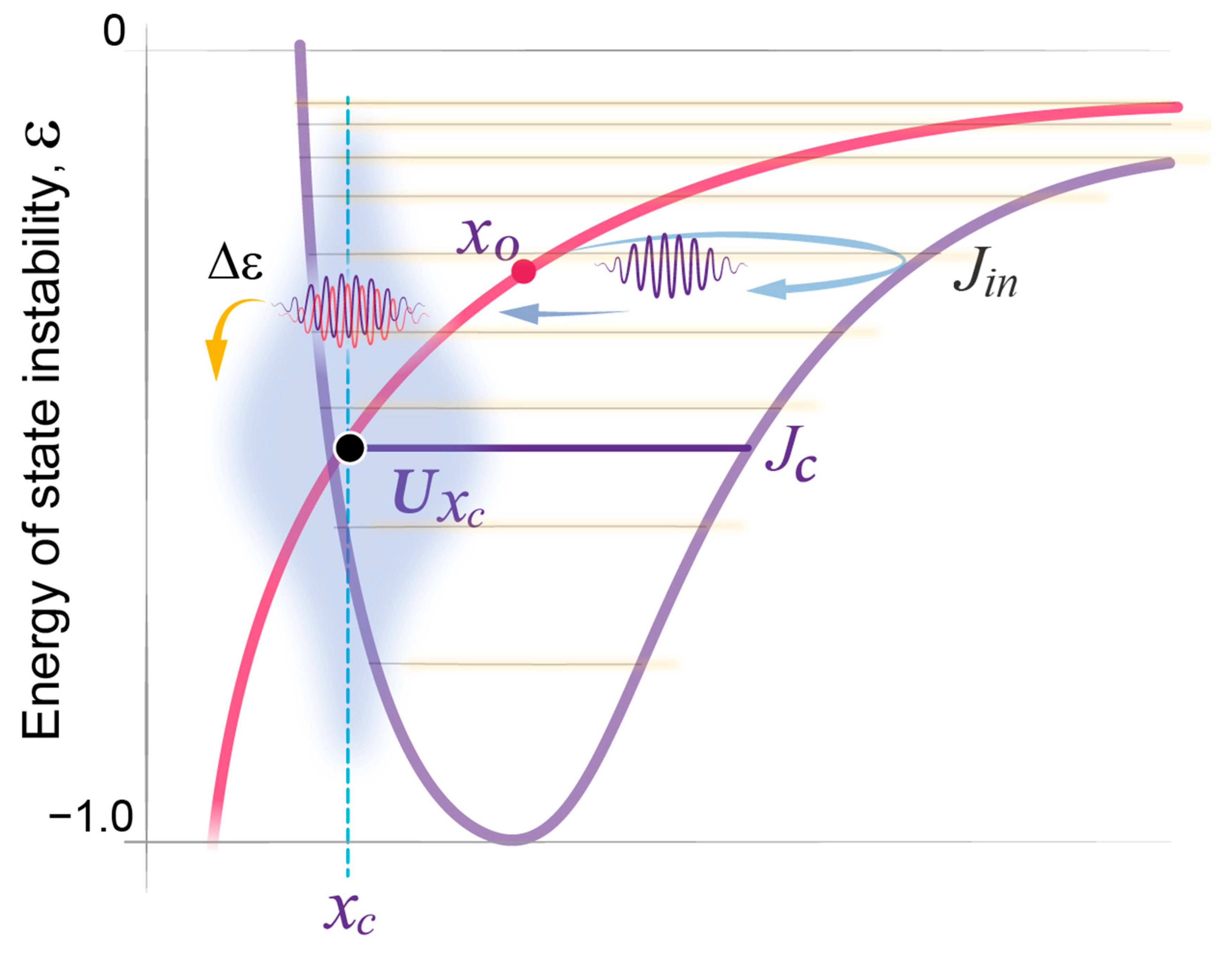
- Fast and slow components: In a diatomic molecule, the dynamics encompass reciprocal rapid changes in electronic states alongside the slower adjustments of the regulating parameter, the distance between the molecule’s two nuclei (R). In our model, we suggest that changes in the biochemical and electrochemical processes that form sleep and wake states are relatively fast, whereas variations in state stability, reflected in the regulating parameter of state stability, denoted as x, occur more gradually, akin to alterations in R. In both types of systems, there is a reciprocal relationship between their fast and slow components.As noted in Section 3, the sleep state engages in reciprocal interactions with a multitude of physiological functions and it remains unknown whether a single molecular or systemic parameter central to the sleep state exists, or if the regulating parameter reflects the combined effects of multiple processes on state stability that are integrated by the neuronal structures involved in state transitions [1]. For the purpose of the mathematical modeling of sleep dynamics presented here, the regulating parameter x is regarded as a one-dimensional reduction of numerous regulatory physiological parameters that collectively define the system’s capacity to maintain stability.
- (b)
- State stability of the fast component. In the molecular system, the electron component can be in a stable (ground) or unstable (excited) state, which are states of different symmetry at a given value of R. Changes in R can result in the swap of symmetry of state, such that a former ground state becomes unstable, while a former excited state becomes stable. Similarly, in our model, the stability of sleep and wake states is determined by the regulating parameter x, and changes depend on variations in x value. Low x values favor the wake state and high x values favor the sleep state.
- (c)
- The interaction and feedback relationship between the fast and slow components. For different electronic states in a diatomic molecule, the energy of the fast (electronic) component depends on R, and this dependence creates a potential energy U(R) for the slow nuclear motion. We expect a similar relationship within sleep and wake dynamics, where parameter regulates the stability of the underlying fast processes, e.g., individual homeostatic loops. In turn, those fast processes modulate the dynamics of the regulating parameter , creating distinct potential energies and respectively.
- (d)
- Probability waves. The wave nature of probabilistic processes can be illustrated by probability waves, or de Broglie waves, which describe the dynamics of the electronic and nuclear components in a diatomic molecule. The nondeterministic nature of the sleep and wake processes, as well as the coherent dynamics of their slow and fast components, suggests the use of quantum mechanical analogies in the description of sleep architecture.
- (e)
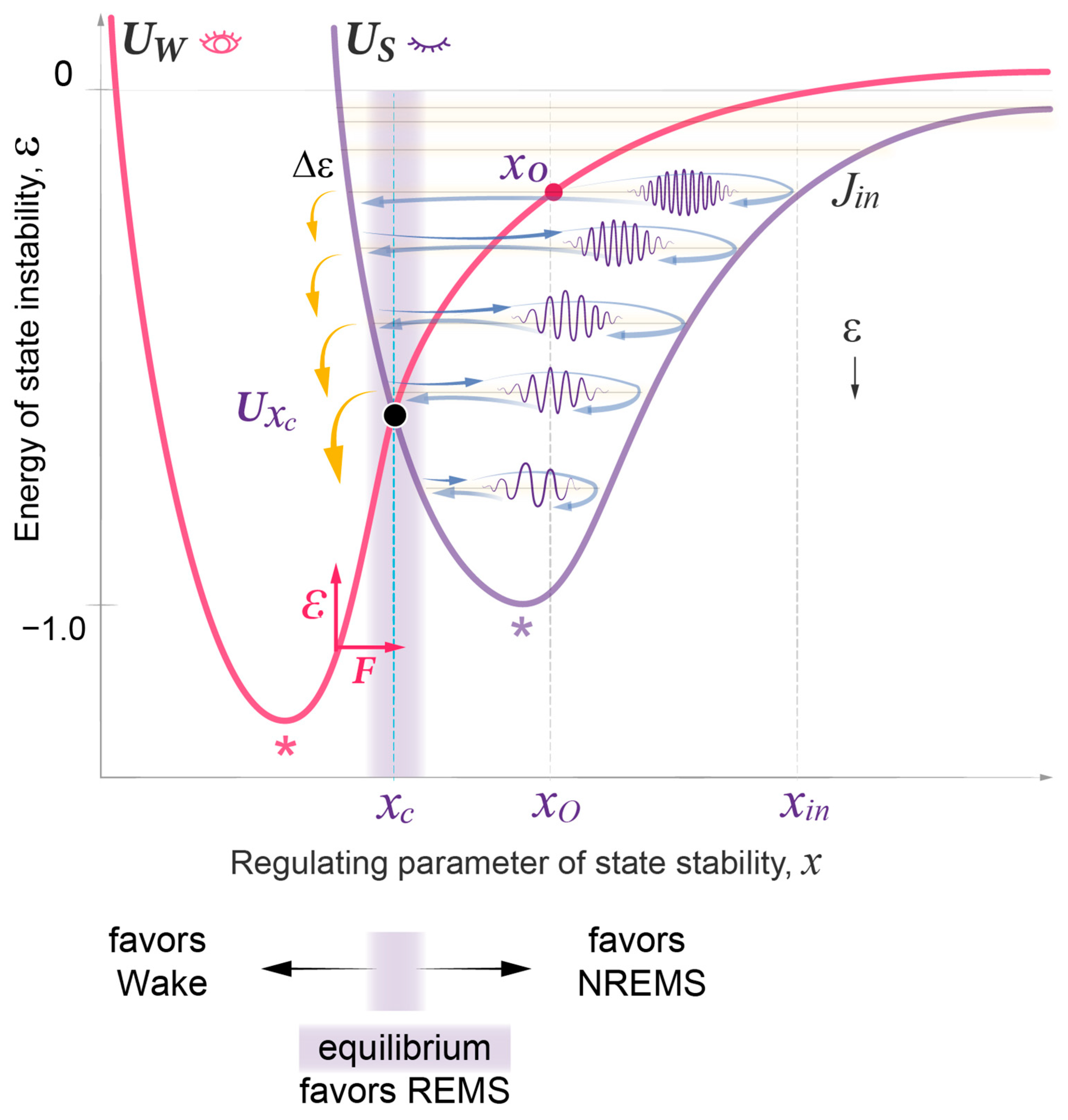
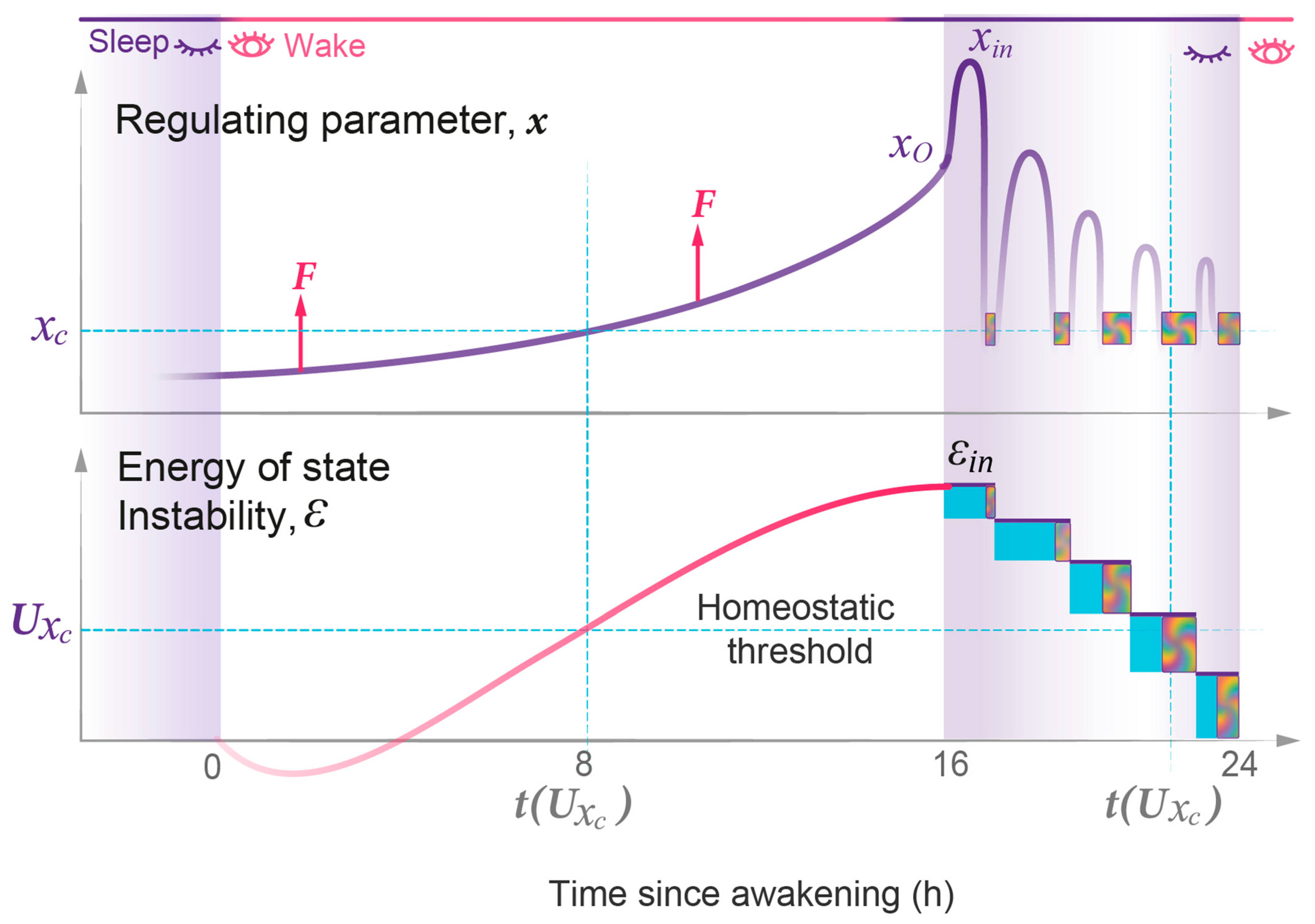
- State transitions. The probabilistic transitions between electronic states of different symmetry, that is the swapping of stable and metastable (excited) states, can occur within certain restricted regions of the R parameter where the electronic energies of different states have close values. We predict that transitions between the sleep and wake states will have similar behavior in the region of crossing or pseudo-crossing of the and potential curves. xc represents the point of sleep–wake homeostatic equilibrium, with Uxc as the homeostatic energy threshold or setpoint (Figure 2).
- (f)
- Discrete energy spectra of the stationary probability waves. In the molecular system, stationary probability waves have a discrete spectrum of energy for both the electronic and nuclear components, the latter being represented by R-vibrations. Analogously, in our model, we introduce the energy parameter ε, which represents the measure of instability for either the wake or sleep state. An increase in state instability leads to an increase in ε.
4.2. Mathematical Apparatus of the Wave Model of Sleep Dynamics
4.2.1. Wave Equations for the Sleep and Wake States
4.2.2. S and W Stationary Waves
4.2.3. Energy Spectra of the S waves in the Morse Potential
4.2.4. Energy Relaxation and Structure of the S Wavepacket
4.2.5. Quasi-Classical Motion of the Wavepacket
4.2.6. Interaction between S and W Stationary Waves and Their Coherent Superposition
4.2.7. The Coherent Superposition of S and W Waves and Wavepacket Dynamics
4.2.8. Short Time-Delay Induced in Landau–Zener Transition
4.2.9. Resonance Enhancement of REMS Episode Duration and Energy Release
4.2.10. Mechanisms of Resonance Formation
4.2.11. Lorentz Resonance Curve
4.2.12. NREMS and REMS Episode Durations in Absolute Time Units
4.2.13. Predictions of NREMS Intensity as a Function of Initial Energy and Amplitude of X-Oscillations
4.2.14. Dynamics of REMS Intensity Depends on Energy Release
4.2.15. Sleep Cycle Invariant
4.2.16. Datasets
4.2.17. Statistical Testing
5. Conclusions
- The wave model of sleep dynamics represents the sleep and wake states as interacting probability waves. By applying the mathematical apparatus of wave mechanics developed for the analysis of probability waves and state transitions on the molecular level, the model provides a precise quantitative description of the typical dynamics of four principal measures of normal sleep architecture, including the durations and intensities of consecutive episodes of NREM and REM sleep, as documented in experimental groups of young healthy adults.
- The model demonstrates that the duration and intensity of consecutive episodes of NREM sleep, as well as the intensity of REM sleep episodes, only reflect the behavior of the sleep probability wave. The variability in these sleep measures between experimental groups can be described by two model parameters: first, the width of the potential well containing the sleep wave, which depends on the habitual sleep duration; and second, the energy level of state instability reached by the time of sleep initiation, which depends on the duration of prior wakefulness.
- The model describes REM sleep as a coherent superposition of sleep and wake waves. Accordingly, an accurate description of the duration of REM sleep episodes depends on the strength of interaction between these two waves, which is maximal in the region of their homeostatic equilibrium and further enhanced by the resonance between them.
- The model establishes an invariant relationship between NREM and REM sleep by predicting that the product of NREM sleep duration and REM sleep intensity should normally remain constant over consecutive sleep cycles. The analysis of experimental group data collected in young adults with high quality sleep confirms this prediction, indicating an intrinsic connection between NREM and REM sleep as part of the same two-step sleep process.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scammell, T.E.; Arrigoni, E.; Lipton, J.O. Neural Circuitry of Wakefulness and Sleep. Neuron 2017, 93, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A.; Achermann, P. Sleep homeostasis and models of sleep regulation. J. Biol. Rhythms. 1999, 14, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.M.; Frank, M.G.; Wisor, J.P.; Roy, S. Sleep function: Toward elucidating an enigma. Sleep Med. Rev. 2016, 28, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Jaggard, J.B.; Wang, G.X.; Mourrain, P. Non-REM and REM/paradoxical sleep dynamics across phylogeny. Curr. Opin. Neurobiol. 2021, 71, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Aserinsky, E.; Kleitman, N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science 1953, 118, 273–274. [Google Scholar] [CrossRef]
- Peever, J.; Fuller, P.M. The Biology of REM Sleep. Curr. Biol. 2017, 27, R1237–R1248. [Google Scholar] [CrossRef]
- Goel, N.; Rao, H.; Durmer, J.S.; Dinges, D.F. Neurocognitive consequences of sleep deprivation. Semin. Neurol. 2009, 29, 320–339. [Google Scholar] [CrossRef]
- Doran, S.M.; Van Dongen, H.P.A.; Dinges, D.F. Sustained attention performance during sleep deprivation: Evidence of state instability. Arch. Ital. Biol. 2001, 139, 253–267. [Google Scholar]
- Zhou, X.; Ferguson, S.A.; Matthews, R.W.; Sargent, C.; Darwent, D.; Kennaway, D.J.; Roach, G.D. Dynamics of neurobehavioral performance variability under forced desynchrony: Evidence of state instability. Sleep 2011, 34, 57–63. [Google Scholar] [CrossRef]
- Cajochen, C.; Khalsa, S.B.S.; Wyatt, J.K.; Czeisler, C.A.; Dijk, D.J. EEG and ocular correlates of circadian melatonin phase and human performance decrements during sleep loss. Am. J. Physiol. 1999, 277, R640. [Google Scholar] [CrossRef]
- Glos, M.; Fietze, I.; Blau, A.; Baumann, G.; Penzel, T. Cardiac autonomic modulation and sleepiness: Physiological consequences of sleep deprivation due to 40 h of prolonged wakefulness. Physiol. Behav. 2014, 125, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Sauvet, F.; Leftheriotis, G.; Gomez-Merino, D.; Langrume, C.; Drogou, C.; Van Beers, P.; Bourrilhon, C.; Florence, G.; Chennaoui, M. Effect of acute sleep deprivation on vascular function in healthy subjects. J. Appl. Physiol. 2010, 108, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Hilton, H.J.; Gates, G.J.; Jelic, S.; Stern, Y.; Bartels, M.N.; Demeersman, R.E.; Basner, R.C. Increased sympathetic and decreased parasympathetic cardiovascular modulation in normal humans with acute sleep deprivation. J. Appl. Physiol. 2005, 98, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Durmer, J.S.; Dinges, D.F. Neurocognitive consequences of sleep deprivation. Semin. Neurol. 2005, 25, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Achermann, P.; Borbély, A.A. Mathematical models of sleep regulation. Front. Biosci. 2003, 8, s683–s693. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Robinson, P.A.; Klerman, E.B. Arousal state feedback as a potential physiological generator of the ultradian REM/NREM sleep cycle. J. Theor. Biol. 2013, 319, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Le Bon, O. Relationships between REM and NREM in the NREM-REM sleep cycle: A review on competing concepts. Sleep Med. 2020, 70, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Heller, H.C. Question what is “known”. Neurobiol. Sleep Circadian Rhythms. 2021, 10, 100062. [Google Scholar] [CrossRef]
- Benington, J.H.; Heller, H.C. Does the function of REM sleep concern non-REM sleep or waking? Prog. Neurobiol. 1994, 44, 433–449. [Google Scholar] [CrossRef]
- Massaquoi, S.G.; McCarley, R.W. Extension of the limit cycle reciprocal interaction model of REM cycle control. An integrated sleep control model. J. Sleep Res. 1992, 1, 138–143. [Google Scholar] [CrossRef]
- Ashby, W.R. Design for a Brain: The Origin of Adaptive Behavior, 2nd ed.; Chapman: London, UK, 1960. [Google Scholar]
- Kelso, J.A.S. Dynamic Patterns: The Self-Organization of Brain and Behavior; MIT Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Deneke, V.E.; Di Talia, S. Chemical waves in cell and developmental biology. J. Cell Biol. 2018, 217, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Landau, L.D.; Lifshitz, E.M. Quantum Mechanics. Non-Relativistic Theory; Pergamon Press: Oxford, UK, 1977; ISBN 9780080209401. [Google Scholar]
- Bush, J.; Oza, A. Hydrodynamic quantum analogs. Rep. Prog. Phys. 2020, 84, 017001. [Google Scholar] [CrossRef] [PubMed]
- Couder, Y.; Fort, E. Single-Particle Diffraction and Interference at a Macroscopic Scale. Phys. Rev. Lett. 2006, 97, 154101. [Google Scholar] [CrossRef] [PubMed]
- Eddi, A.; Moukhtar, J.; Perrard, S.; Fort, E.; Couder, Y. Level Splitting at Macroscopic Scale. Phys. Rev. Lett. 2012, 108, 264503. [Google Scholar] [CrossRef]
- Vandewalle, N. Bouncing droplets mimic spin systems. Nature 2021, 596, 41. [Google Scholar] [CrossRef]
- Perrard, S.; Labousse, M.; Miskin, M.; Fort, E.; Couder, Y. Self-organization into quantized eigenstates of a classical wave-driven particle. Nat. Commun. 2014, 5, 3219. [Google Scholar] [CrossRef]
- Barbato, G.; Wehr, T.A. Homeostatic regulation of REM sleep in humans during extended sleep. Sleep 1998, 21, 267–276. [Google Scholar] [CrossRef]
- Barbato, G.; Barker, C.; Bender, C.; Giesen, H.A.; Wehr, T.A. Extended sleep in humans in 14 hour nights (LD 10:14): Relationship between REM density and spontaneous awakening. Electroencephalogr. Clin. Neurophysiol. 1994, 90, 291–297. [Google Scholar] [CrossRef]
- Dijk, D.J.; Brunner, D.P.; Borbély, A.A. Time course of EEG power density during long sleep in humans. Am. J. Physiol. 1990, 258 Pt 2, R650–R661. [Google Scholar] [CrossRef]
- Aserinsky, E. The maximal capacity for sleep: Rapid eye movement density as an index of sleep satiety. Biol. Psychiatry 1969, 1, 147–159. [Google Scholar]
- Aserinsky, E. Relationship of rapid eye movement density to the prior accumulation of sleep and wakefulness. Psychophysiology 1973, 10, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Marzano, C.; De Simoni, E.; Tempesta, D.; Ferrara, M.; De Gennaro, L. Sleep deprivation suppresses the increase of rapid eye movement density across sleep cycles. J. Sleep Res. 2011, 20, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.B.; Conroy, D.A.; Duffy, J.F.; Czeisler, C.A.; Dijk, D.J. Sleep- and circadian-dependent modulation of REM density. J. Sleep Res. 2002, 11, 53–59. [Google Scholar] [CrossRef]
- Arfken, C.L.; Joseph, A.; Sandhu, G.R.; Roehrs, T.; Douglass, A.B.; Boutros, N.N. The status of sleep abnormalities as a diagnostic test for major depressive disorder. J. Affect Disord. 2014, 156, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; Diederich, N.J.; Schroeder, L.A.; Schmidt, M.; Rüegg, S.; Khatami, R. Time to reconsider REM density in sleep research. Clin. Neurophysiol. 2022, 137, 63–65. [Google Scholar] [CrossRef]
- Aserinsky, E. Rapid eye movement density and pattern in the sleep of normal young adults. Psychophysiology 1971, 8, 361–375. [Google Scholar] [CrossRef]
- Skorucak, J.; Arbon, E.L.; Dijk, D.J.; Achermann, P. Response to chronic sleep restriction, extension, and subsequent total sleep deprivation in humans: Adaptation or preserved sleep homeostasis? Sleep 2018, 41, zsy078. [Google Scholar] [CrossRef]
- Wineland, D.J. Nobel Lecture: Superposition, entanglement, and raising Schrödinger’s cat. Rev. Mod. Phys. 2013, 85, 1103. [Google Scholar] [CrossRef]
- Jouvet, M. Paradoxical sleep-A study of its nature and mechanisms. Prog. Brain Res. 1965, 18, 20–62. [Google Scholar] [CrossRef]
- Chow, H.M.; Horovitz, S.G.; Carr, W.S.; Picchioni, D.; Coddington, N.; Fukunaga, M.; Xu, Y.; Balkin, T.J.; Duyn, J.H.; Braun, A.R. Rhythmic alternating patterns of brain activity distinguish rapid eye movement sleep from other states of consciousness. Proc. Natl. Acad. Sci. USA 2013, 110, 10300–10305. [Google Scholar] [CrossRef]
- Benca, R.M.; Okawa, M.; Uchiyama, M.; Ozaki, S.; Nakajima, T.; Shibui, K.; Obermeyer, W.H. Sleep and mood disorders. Sleep Med. Rev. 1997, 1, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Zangani, C.; Casetta, C.; Saunders, A.S.; Donati, F.; Maggioni, E.; D’Agostino, A. Sleep abnormalities across different clinical stages of Bipolar Disorder: A review of EEG studies. Neurosci. Biobehav. Rev. 2020, 118, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Li, R.; Zhang, M.Q.; Zhang, Z.; Qu, W.M.; Huang, Z.L. The Neurobiological Mechanisms and Treatments of REM Sleep Disturbances in Depression. Curr. Neuropharmacol. 2015, 13, 543–553. [Google Scholar] [CrossRef]
- Steiger, A.; Pawlowski, M. Depression and Sleep. Int. J. Mol. Sci. 2019, 20, 607. [Google Scholar] [CrossRef] [PubMed]
- Buysse, D.J.; Tu, X.M.; Cherry, C.R.; Begley, A.E.; Kowalski, J.; Kupfer, D.J.; Frank, E. Pretreatment REM sleep and subjective sleep quality distinguish depressed psychotherapy remitters and nonremitters. Biol. Psychiatry 1999, 45, 205–213. [Google Scholar] [CrossRef]
- Lechinger, J.; Koch, J.; Weinhold, S.L.; Seeck-Hirschner, M.; Stingele, K.; Kropp-Näf, C.; Braun, M.; Drews, H.J.; Aldenhoff, J.; Huchzermeier, C.; et al. REM density is associated with treatment response in major depression: Antidepressant pharmacotherapy vs. psychotherapy. J. Psychiatr. Res. 2021, 133, 67–72. [Google Scholar] [CrossRef]
- Modell, S.; Ising, M.; Holsboer, F.; Lauer, C.J. The Munich vulnerability study on affective disorders: Premorbid polysomnographic profile of affected high-risk probands. Biol. Psychiatry 2005, 58, 694–699. [Google Scholar] [CrossRef]
- Kowatch, R.A.; Schnoll, S.S.; Knisely, J.S.; Green, D.; Elswick, R.K. Electroencephalographic sleep and mood during cocaine withdrawal. J. Addict. Dis. 1992, 11, 21–45. [Google Scholar] [CrossRef]
- Clark, C.P.; Gillin, J.C.; Golshan, S.; Demodena, A.; Smith, T.L.; Danowski, S.; Irwin, M.; Schuckit, M. Increased REM sleep density at admission predicts relapse by three months in primary alcoholics with a lifetime diagnosis of secondary depression. Biol. Psychiatry 1998, 43, 601–607. [Google Scholar] [CrossRef]
- Schroeder, L.A.; Rufra, O.; Sauvageot, N.; Fays, F.; Pieri, V.; Diederich, N.J. Reduced Rapid Eye Movement Density in Parkinson Disease: A Polysomnography-Based Case-Control Study. Sleep 2016, 39, 2133–2139. [Google Scholar] [CrossRef]
- Czeisler, C.A.; Gooley, J.J. Sleep and circadian rhythms in humans. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 579–597. [Google Scholar] [CrossRef] [PubMed]
- Kleitman, N. Sleep and Wakefulness; University of Chicago Press: Chicago, IL, USA, 1963. [Google Scholar]
- Lavie, P. Ultradian rhythms in arousal—The problem of masking. Chronobiol. Int. 1989, 6, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Mott, N.F.; Massey, H.S. Theory of Atomic Collisions; Clarendon Press: Oxford, UK, 1965; ISBN 0198512422. [Google Scholar]
- Migdal, A.B. Qualitative Methods in Quantum Theory; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar] [CrossRef]
- Bates, D.R.; Lynn, N. Electron Capture of the Accidental Resonance Type. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1959, 253, 141–153. [Google Scholar]
- Goldberger, M.L.; Watson, K.M. Collision Theory; Dover Publications, Inc.: Mineola, NY, USA, 2004. [Google Scholar]
- Zhu, C. Analytical semiclassical theory for general non-adiabatic transition and tunneling. Phys. Scr. 2009, 80, 048114. [Google Scholar] [CrossRef]
- Sun, G.; Xueda Wen, X.; Gong, M.; Zhang, D.-W.; Yu, Y.; Zhu, S.-L.; Chen, J.; Wu, P.; Han, S. Observation of coherent oscillation in single-passage Landau-Zener transitions. Sci. Rep. 2015, 5, 8463. [Google Scholar] [CrossRef]
- Tayebirad, G.; Zenesini, A.; Ciampini, D.; Mannella, R.; Morsch, O.; Arimondo, E.; Lörch, N.; Wimberger, S. Time-resolved measurement of Landau-Zener tunneling in different bases. Phys. Rev. A 2010, 82, 013633. [Google Scholar] [CrossRef]
- Rubbmark, J.R.; Kash, M.M.; Littman, M.G.; Kleppner, D. Dynamical effects at avoided level crossings: A study of the Landau-Zener effect using Rydberg atoms. Phys. Rev. A 1981, 23, 3107. [Google Scholar] [CrossRef]
- Cote, R. Role of resonances at ultracold temperatures. In Cold Chemistry: Molecular Scattering and Reactivity Near Absolute Zero; Dulieu, O., Osterwalder, A., Eds.; Royal Society of Chemistry: London, UK, 2018; pp. 313–388. ISBN 9781782625971. [Google Scholar]
- Coveney, P.V.; Child, M.S.; Barany, A. The two-state S matrix for the Landau-Zener potential curve crossing model: Predissociation and resonant scattering. J. Phys. B At. Mol. Phys. 1985, 18, 4557–4580. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.; Harding, S.M.; Lloyd, R.M.; Quan, S.F.; Vaughn, B.V. AASM Scoring Manual Updates for 2017 (Version 2.4). J. Clin. Sleep. Med. 2017, 13, 665–666. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kharchenko, V.; Zhdanova, I.V. The Wave Model of Sleep Dynamics and an Invariant Relationship between NonREM and REM Sleep. Clocks & Sleep 2023, 5, 686-716. https://doi.org/10.3390/clockssleep5040046
Kharchenko V, Zhdanova IV. The Wave Model of Sleep Dynamics and an Invariant Relationship between NonREM and REM Sleep. Clocks & Sleep. 2023; 5(4):686-716. https://doi.org/10.3390/clockssleep5040046
Chicago/Turabian StyleKharchenko, Vasili, and Irina V. Zhdanova. 2023. "The Wave Model of Sleep Dynamics and an Invariant Relationship between NonREM and REM Sleep" Clocks & Sleep 5, no. 4: 686-716. https://doi.org/10.3390/clockssleep5040046