
Does Motor Memory Reactivation through Practice and Post-Learning Sleep Modulate Consolidation?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Demographic Data
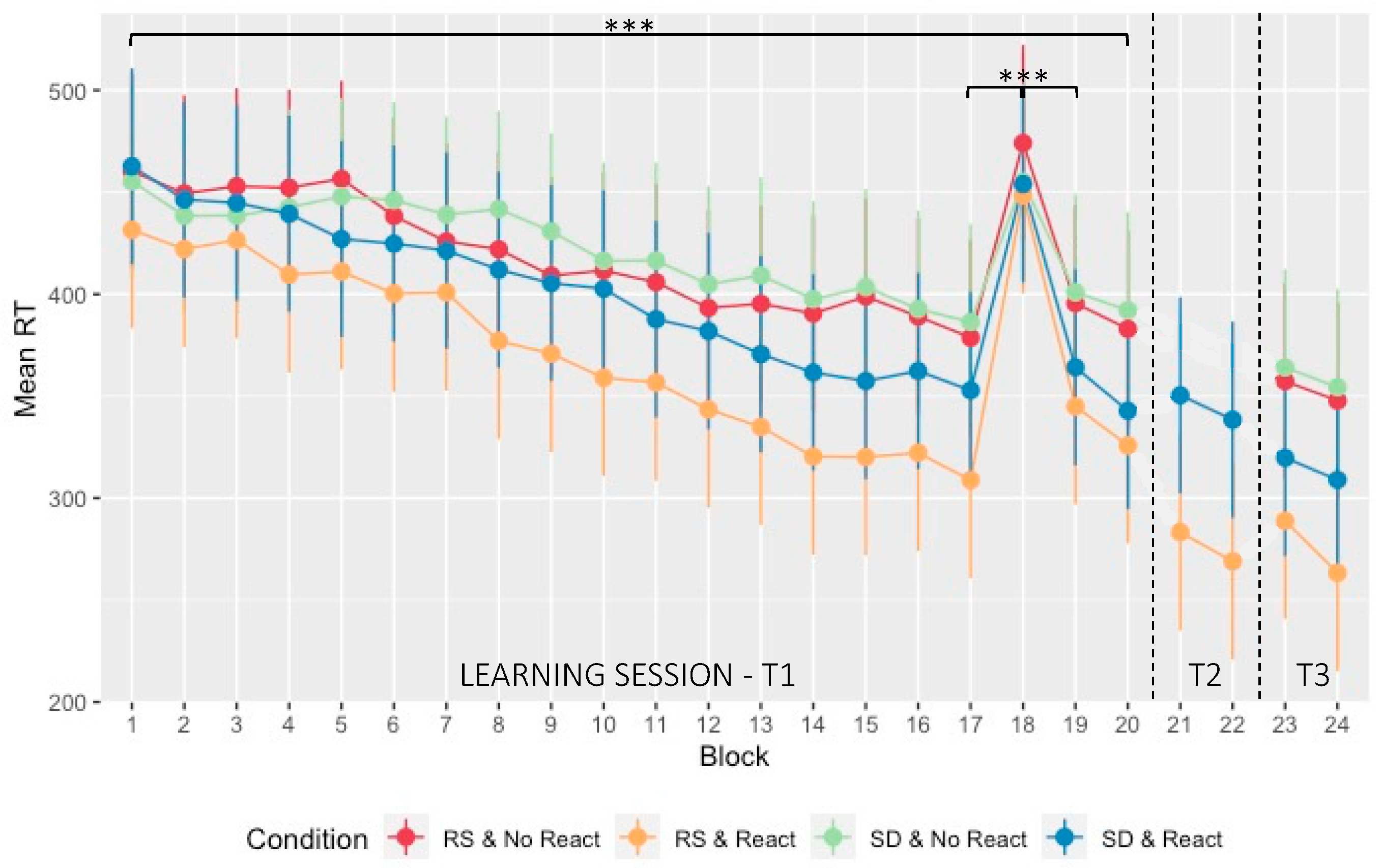
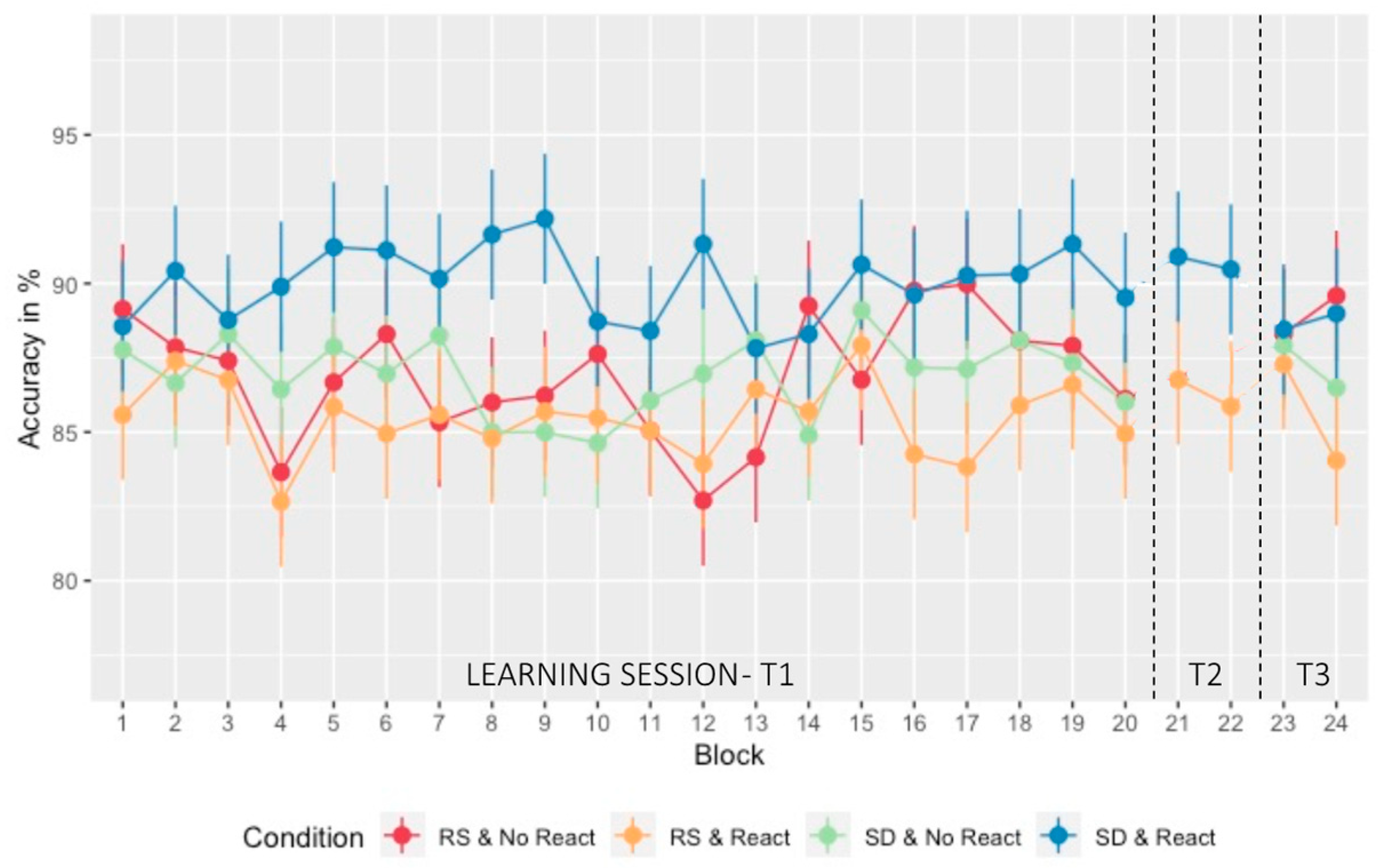
2.2. Learning Session
2.3. Difference in Performance at T2
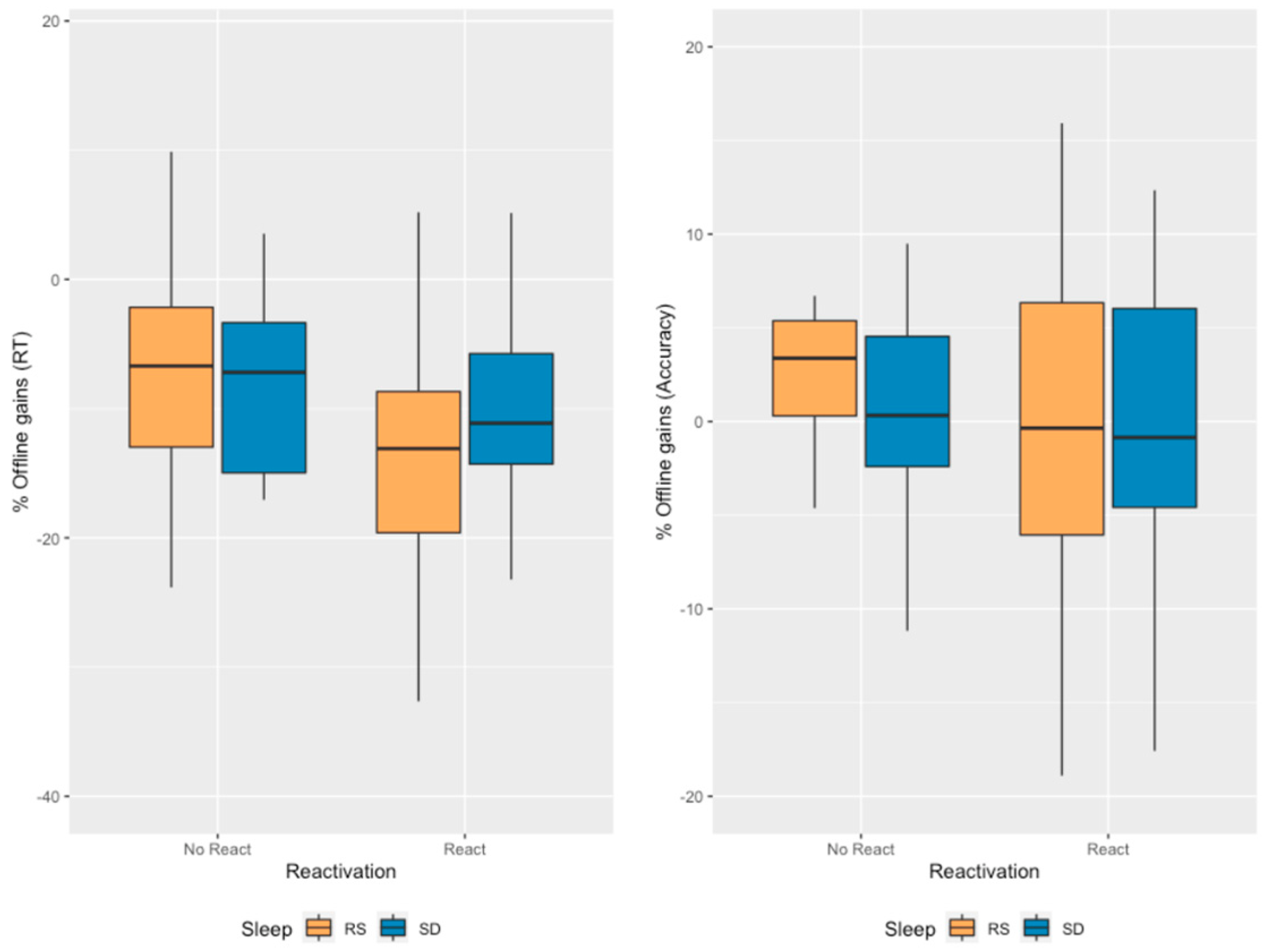
2.4. Motor Memory Consolidation Modulated by Post-Training Sleep and Reactivation Conditions
2.5. Additional Control Analyses
2.5.1. Alertness before Learning and Testing Sessions
2.5.2. Sleep Quality and Duration on the Experimental RS Night
2.5.3. Sleep Quality and Duration during the Three Recovery Nights
2.5.4. Vigilance and Sleepiness throughout the Experimental SD Night
3. Discussion
4. Materials and Methods
4.1. Participants
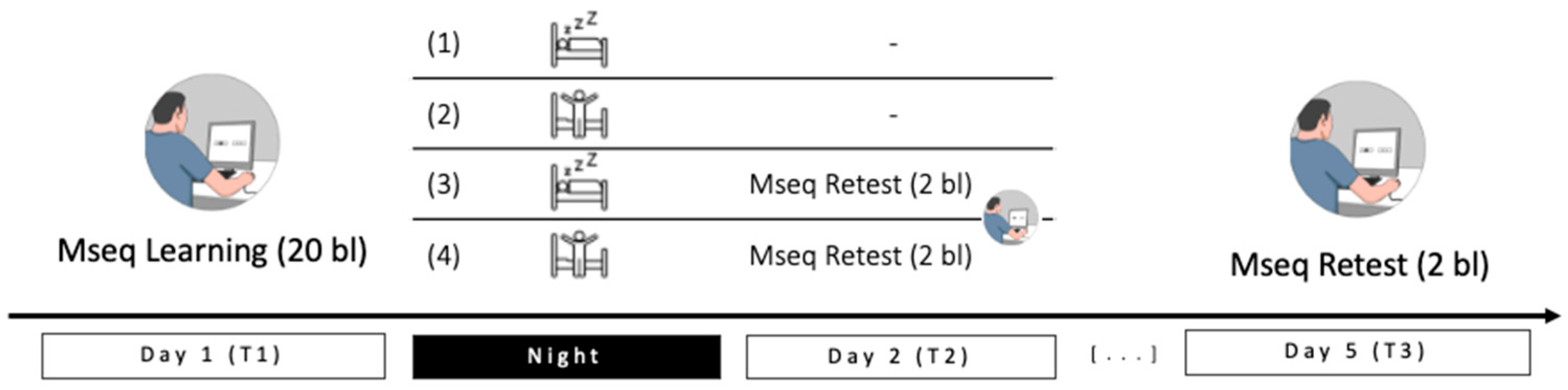
4.2. General Procedure
4.3. Motor Task
4.4. Performance Assessment and Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lechner, H.A.; Squire, L.R.; Byrne, J.H. 100 Years of Consolidation—Remembering Müller and Pilzecker. Learn. Mem. 1999, 6, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Stern, S.A.; Alberini, C.M. Mechanisms of memory enhancement. Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 5, 37–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G.M. Synaptic plasticity and memory: An evaluation of the hypothesis. Annu. Rev. Neurosci. 2000, 23, 649–711. [Google Scholar] [CrossRef] [Green Version]
- Frankland, P.W.; Bontempi, B. The organization of recent and remote memories. Nat. Rev. Neurosci. 2005, 6, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Dudai, Y.; Karni, A.; Born, J. The Consolidation and Transformation of Memory. Neuron 2015, 88, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.B.; Soares, A.B. Reconsolidation of human motor memory: From boundary conditions to behavioral interventions-How far are we from clinical applications? Behav. Brain Res. 2018, 353, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Nader, K.; Hardt, O. A single standard for memory: The case for reconsolidation. Nat. Rev. Neurosci. 2009, 10, 224–234. [Google Scholar] [CrossRef]
- Nader, K.; Schafe, G.E.; le Doux, J.E. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature 2000, 406, 722–726. [Google Scholar] [CrossRef]
- Tronson, N.C.; Taylor, J.R. Molecular mechanisms of memory reconsolidation. Nat. Rev. Neurosci. 2007, 8, 262–275. [Google Scholar] [CrossRef]
- Lee, J.L.C. Reconsolidation: Maintaining memory relevance. Curr. Biol. 2012, 22, R66–R71. [Google Scholar] [CrossRef] [Green Version]
- Forcato, C.; Fernandez, R.S.; Pedreira, M.E. The role and dynamic of strengthening in the reconsolidation process in a human declarative memory: What decides the fate of recent and older memories? PLoS ONE 2013, 8, e61688. [Google Scholar] [CrossRef] [Green Version]
- Sandrini, M.; Censor, N.; Mishoe, J.; Cohen, L.G. Causal Role of Prefrontal Cortex in Strengthening of Episodic Memories through Reconsolidation. Curr. Biol. 2013, 23, 2181–2184. [Google Scholar] [CrossRef] [Green Version]
- Herszage, J.; Sharon, H.; Censor, N. Reactivation-induced motor skill learning. Proc. Natl. Acad. Sci. USA 2021, 118, 2102242118. [Google Scholar] [CrossRef]
- Wymbs, N.F.; Bastian, A.J.; Celnik, P.A. Motor skills are strengthened through reconsolidation. Curr. Biol. 2016, 26, 338. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.P.; Brakefield, T.; Hobson, J.A.; Stickgold, R. Dissociable stages of human memory consolidation and reconsolidation. Nature 2003, 425, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Censor, N.; Horovitz, S.G.; Cohen, L.G. Interference with existing memories alters offline intrinsic functional brain connectivity. Neuron 2014, 81, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Censor, N.; Dimyan, M.A.; Cohen, L.G. Modification of Existing Human Motor Memories Is Enabled by Primary Cortical Processing during Memory Reactivation. Curr. Biol. 2010, 20, 1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haubrich, J.; Nader, K. Memory Reconsolidation. Curr. Top. Behav. Neurosci. 2018, 37, 151–176. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, P. Sleep and memory in the making. Are current concepts sufficient in children? Arch. Ital. Biol. 2014, 152, 79–92. [Google Scholar] [CrossRef]
- Havekes, R.; Meerlo, P.; Abel, T. Animal studies on the role of sleep in memory: From behavioral performance to molecular mechanisms. Curr. Top. Behav. Neurosci. 2015, 25, 183–206. [Google Scholar] [CrossRef]
- Rasch, B.; Born, J. About sleep’s role in memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub, J.B.; Biswal, S.; Peled, N.; Rivilis, N.; Golby, A.J.; Lee, J.W.; Westover, M.B.; Halgren, E.; Cash, S.S. Reactivation of Motor-Related Gamma Activity in Human NREM Sleep. Front. Neurosci. 2020, 14, 449. [Google Scholar] [CrossRef] [PubMed]
- Maquet, P.; Laureys, S.; Peigneux, P.; Fuchs, S.; Petiau, C.; Phillips, C.; Cleeremans, A. Experience-dependent changes in cerebral activation during human REM sleep. Nat. Neurosci. 2000, 3, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, P.; Laureys, S.; Fuchs, S.; Destrebecqz, A.; Collette, F.; Delbeuck, X.; Phillips, C.; Aerts, J.; Del Fiore, G.; Degueldre, C.; et al. Learned material content and acquisition level modulate cerebral reactivation during posttraining rapid-eye-movements sleep. Neuroimage 2003, 20, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Fogel, S.; Albouy, G.; King, B.R.; Lungu, O.; Vien, C.; Bore, A.; Pinsard, B.; Benali, H.; Carrier, J.; Doyon, J. Reactivation or transformation? Motor memory consolidation associated with cerebral activation time-locked to sleep spindles. PLoS ONE 2017, 12, e0174755. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, D.S.; Gulati, T.; Ganguly, K. Sleep-Dependent Reactivation of Ensembles in Motor Cortex Promotes Skill Consolidation. PLoS Biol. 2015, 13, e1002263. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Lai, C.S.W.; Cichon, J.; Ma, L.; Li, W.; Gan, W.B. Sleep promotes branch-specific formation of dendritic spines after learning. Science 2014, 344, 1173–1178. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.; Goyal, V.; Mello, C.; Pavlides, C. Brain Gene Expression During REM Sleep Depends on Prior Waking Experience. Learn. Mem. 1999, 6, 500. [Google Scholar] [CrossRef] [Green Version]
- Karni, A.; Meyer, G.; Rey-Hipolito, C.; Jezzard, P.; Adams, M.M.; Turner, R.; Ungerleider, L.G. The acquisition of skilled motor performance: Fast and slow experience-driven changes in primary motor cortex. Proc. Natl. Acad. Sci. USA 1998, 95, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Karni, A.; Meyer, G.; Jezzard, P.; Adams, M.M.; Turner, R.; Ungerleider, L.G. Functional MRI evidence for adult motor cortex plasticity during motor skill learning. Nature 1995, 377, 155–158. [Google Scholar] [CrossRef]
- Schmid, D.; Erlacher, D.; Klostermann, A.; Kredel, R.; Hossner, E.J. Sleep-dependent motor memory consolidation in healthy adults: A meta-analysis. Neurosci. Biobehav. Rev. 2020, 118, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.P.; Brakefield, T.; Seidman, J.; Morgan, A.; Hobson, J.A.; Stickgold, R. Sleep and the time course of motor skill learning. Learn. Mem. 2003, 10, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Nissen, C.; Kloepfer, C.; Feige, B.; Piosczyk, H.; Spiegelhalder, K.; Voderholzer, U.; Riemann, D. Sleep-related memory consolidation in primary insomnia. J. Sleep. Res. 2011, 20, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Schönauer, M.; Grätsch, M.; Gais, S. Evidence for two distinct sleep-related long-term memory consolidation processes. Cortex 2015, 63, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Debas, K.; Carrier, J.; Orban, P.; Barakat, M.; Lungu, O.; Vandewalle, G.; Tahar, A.H.; Bellec, P.; Karni, A.; Ungerleider, L.G.; et al. Brain plasticity related to the consolidation of motor sequence learning and motor adaptation. Proc. Natl. Acad. Sci. USA 2010, 107, 17839–17844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blischke, K.; Erlacher, D.; Kresin, H.; Brueckner, S.; Malangré, A. Benefits of sleep in motor learning—Prospects and limitations. J. Hum. Kinet. 2008, 20, 23–36. [Google Scholar] [CrossRef]
- Fogel, S.M.; Albouy, G.; Vien, C.; Popovicci, R.; King, B.R.; Hoge, R.; Jbabdi, S.; Benali, H.; Karni, A.; Maquet, P.; et al. fMRI and sleep correlates of the age-related impairment in motor memory consolidation. Hum. Brain. Mapp. 2014, 35, 3625–3645. [Google Scholar] [CrossRef]
- Simmons, A.L.; Duke, R.A. Effects of Sleep on Performance of a Keyboard Melody. J. Res. Music. Educ. 2016, 54, 257–269. [Google Scholar] [CrossRef]
- Landry, S.; Anderson, C.; Conduit, R. The effects of sleep, wake activity and time-on-task on offline motor sequence learning. Neurobiol. Learn. Mem. 2016, 127, 56–63. [Google Scholar] [CrossRef]
- King, B.R.; Hoedlmoser, K.; Hirschauer, F.; Dolfen, N.; Albouy, G. Sleeping on the motor engram: The multifaceted nature of sleep-related motor memory consolidation. Neurosci. Biobehav. Rev. 2017, 80, 1–22. [Google Scholar] [CrossRef]
- Birbaumer, N. Memory: Reconsolidation Allows Modification of Motor Memories. Curr. Biol. 2010, 20, R709–R710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrini, M.; Cohen, L.G.; Censor, N. Modulating reconsolidation: A link to causal systems-level dynamics of human memories. Trends Cogn. Sci. 2015, 19, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Beukelaar, T.T.; Woolley, D.G.; Wenderoth, N. Gone for 60 seconds: Reactivation length determines motor memory degradation during reconsolidation. Cortex 2014, 59, 138–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Beukelaar, T.T.; Woolley, D.G.; Alaerts, K.; Swinnen, S.P.; Wenderoth, N. Reconsolidation of motor memories is a time-dependent process. Front. Hum. Neurosci. 2016, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Gabitov, E.; Boutin, A.; Pinsard, B.; Censor, N.; Fogel, S.M.; Albouy, G.; King, B.R.; Benali, H.; Carrier, J.; Cohen, L.G.; et al. Re-stepping into the same river: Competition problem rather than a reconsolidation failure in an established motor skill. Sci. Rep. 2017, 7, 9406. [Google Scholar] [CrossRef] [PubMed]
- Gabitov, E.; Boutin, A.; Pinsard, B.; Censor, N.; Fogel, S.M.; Albouy, G.; King, B.R.; Carrier, J.; Cohen, L.G.; Karni, A.; et al. Susceptibility of consolidated procedural memory to interference is independent of its active task-based retrieval. PLoS ONE 2019, 14, e0210876. [Google Scholar] [CrossRef] [Green Version]
- Reis, J.; Schambra, H.M.; Cohen, L.G.; Buch, E.R.; Fritsch, B.; Zarahn, E.; Celnik, P.A.; Krakauer, J.W. Noninvasive cortical stimulation enhances motor skill acquisition over multiple days through an effect on consolidation. Proc. Natl. Acad. Sci. USA 2009, 106, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Nissen, M.J.; Bullemer, P. Attentional requirements of learning: Evidence from performance measures. Cogn. Psychol. 1987, 19, 1–32. [Google Scholar] [CrossRef]
- Basner, M.; Mcguire, S.; Goel, N.; Rao, H.; Dinges, D.F. A new likelihood ratio metric for the psychomotor vigilance test and its sensitivity to sleep loss. J. Sleep. Res. 2015, 24, 702–713. [Google Scholar] [CrossRef]
- Rodriguez-Ortiz, C.J.; Bermúdez-Rattoni, F. Determinants to trigger memory reconsolidation: The role of retrieval and updating information. Neurobiol. Learn. Mem. 2017, 142, 4–12. [Google Scholar] [CrossRef]
- Schultz, W.; Dickinson, A. Neuronal coding of prediction errors. Annu. Rev. Neurosci. 2000, 23, 473–500. [Google Scholar] [CrossRef] [Green Version]
- López, M.A.; Santos, M.J.; Cortasa, S.; Fernández, R.S.; Tano, M.C.; Pedreira, M.E. Different dimensions of the prediction error as a decisive factor for the triggering of the reconsolidation process. Neurobiol. Learn. Mem. 2016, 136, 210–219. [Google Scholar] [CrossRef]
- Hardwicke, T.E.; Taqi, M.; Shanks, D.R. Postretrieval new learning does not reliably induce human memory updating via reconsolidation. Proc. Natl. Acad. Sci. USA 2016, 113, 5206–5211. [Google Scholar] [CrossRef] [Green Version]
- Urbain, C.; Schmitz, R.; Schmidt, C.; Cleeremans, A.; Van Bogaert, P.; Maquet, P.; Peigneux, P. Sleep-dependent neurophysiological processes in implicit sequence learning. J. Cogn. Neurosci. 2013, 25, 2003–2014. [Google Scholar] [CrossRef] [Green Version]
- Borragán, G.; Urbain, C.; Schmitz, R.; Mary, A.; Peigneux, P. Sleep and memory consolidation: Motor performance and proactive interference effects in sequence learning. Brain. Cogn. 2015, 95, 54–61. [Google Scholar] [CrossRef]
- Fitzroy, A.B.; Kainec, K.A.; Seo, J.; Spencer, R.M.C. Encoding and consolidation of motor sequence learning in young and older adults. Neurobiol. Learn. Mem. 2021, 185, 107508. [Google Scholar] [CrossRef]
- Meier, B.; Cock, J. Offline consolidation in implicit sequence learning. Cortex 2014, 57, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Robertson, E.M.; Pascual-Leone, A.; Press, D.Z. Awareness modifies the skill-learning benefits of sleep. Curr. Biol. 2004, 14, 208–212. [Google Scholar] [CrossRef]
- Genzel, L.; Kiefer, T.; Renner, L.; Wehrle, R.; Kluge, M.; Grözinger, M.; Steiger, A.; Dresler, M. Sex and modulatory menstrual cycle effects on sleep related memory consolidation. Psychoneuroendocrinology 2012, 37, 987–998. [Google Scholar] [CrossRef]
- Alonso, A.; Genzel, L.; Gomez, A. Sex and Menstrual Phase Influences on Sleep and Memory. Curr. Sleep. Med. Rep. 2021, 7, 1–14. [Google Scholar] [CrossRef]
- Yonkers, K.A.; O’Brien, P.S.; Eriksson, E. Premenstrual syndrome. Lancet 2008, 371, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms—PubMed. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar] [PubMed]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, K.; Sato, D.; Iguchi, K.; Baba, Y.; Yamashiro, K. Menstrual Cycle Modulates Motor Learning and Memory Consolidation in Humans. Brain Sci. 2020, 10, 696. [Google Scholar] [CrossRef]
- Tucker, M.A.; Hirota, Y.; Wamsley, E.J.; Lau, H.; Chaklader, A.; Fishbein, W. A daytime nap containing solely non-REM sleep enhances declarative but not procedural memory. Neurobiol. Learn Mem. 2006, 86, 241–247. [Google Scholar] [CrossRef]
- Åkerstedt, T.; Gillberg, M. Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef]
- Kaida, K.; Takahashi, M.; Åkerstedt, T.; Nakata, A.; Otsuka, Y.; Haratani, T.; Fukasawa, K. Validation of the Karolinska sleepiness scale against performance and EEG variables. Clin. Neurophysiol. 2006, 117, 1574–1581. [Google Scholar] [CrossRef]
- Dinges, D.I.; Powell, J.W. Microcomputer analysis of performance on a portable, simple visual RT task sustained operations. Behav. Res. Methods Instrum. Comput. 1985, 17, 652–655. [Google Scholar] [CrossRef]
- Borbély, A.A.; Baumann, F.; Brandeis, D.; Strauch, I.; Lehmann, D. Sleep deprivation: Effect on sleep stages and EEG power density in man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 483–493. [Google Scholar] [CrossRef]
- Dinges, D.F. Sleep Deprivation and Vigilant Attention. Mol. Biophys. Mech. Arousal Alertness Atten. 2008, 1129, 305–322. [Google Scholar] [CrossRef]
- Sakai, K.; Kitaguchi, K.; Hikosaka, O. Chunking during human visuomotor sequence learning. Exp. Brain Res. 2003, 152, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Fonollosa, J.; Neftci, E.; Rabinovich, M. Learning of Chunking Sequences in Cognition and Behavior. PLoS Comput. Biol. 2015, 11, e1004592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleeremans, A.; McClelland, J.L. Learning the Structure of Event Sequences. J. Exp. Psychol. Gen. 1991, 120, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Dienes, Z. Bayesian Versus Orthodox Statistics: Which Side Are You On? Perspect. Psychol. Sci. 2011, 6, 274–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stee, W.; Peigneux, P. Does Motor Memory Reactivation through Practice and Post-Learning Sleep Modulate Consolidation? Clocks & Sleep 2023, 5, 72-84. https://doi.org/10.3390/clockssleep5010008
Stee W, Peigneux P. Does Motor Memory Reactivation through Practice and Post-Learning Sleep Modulate Consolidation? Clocks & Sleep. 2023; 5(1):72-84. https://doi.org/10.3390/clockssleep5010008
Chicago/Turabian StyleStee, Whitney, and Philippe Peigneux. 2023. "Does Motor Memory Reactivation through Practice and Post-Learning Sleep Modulate Consolidation?" Clocks & Sleep 5, no. 1: 72-84. https://doi.org/10.3390/clockssleep5010008