
Ecological Forecasting and Operational Information Systems Support Sustainable Ocean Management
 , ,
, ,
Abstract
:1. Introduction
2. Case Studies
2.1. Ecological Forecasts for Fisheries
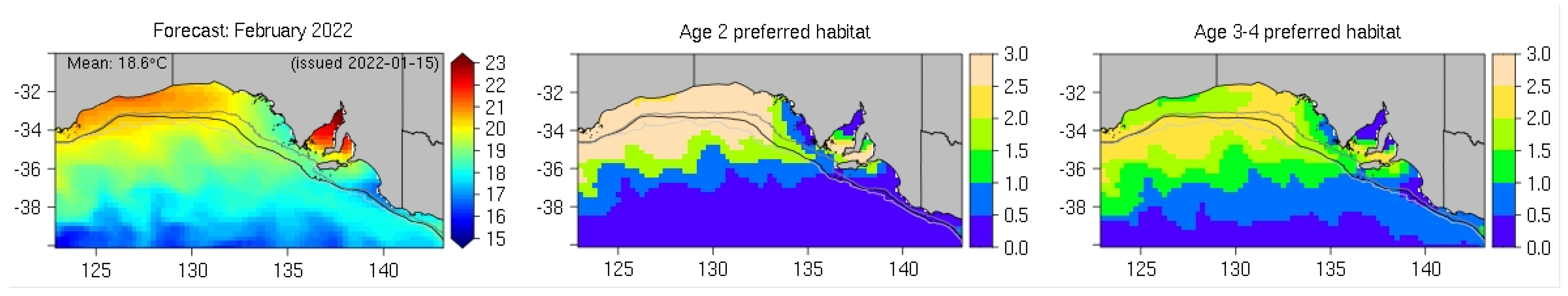
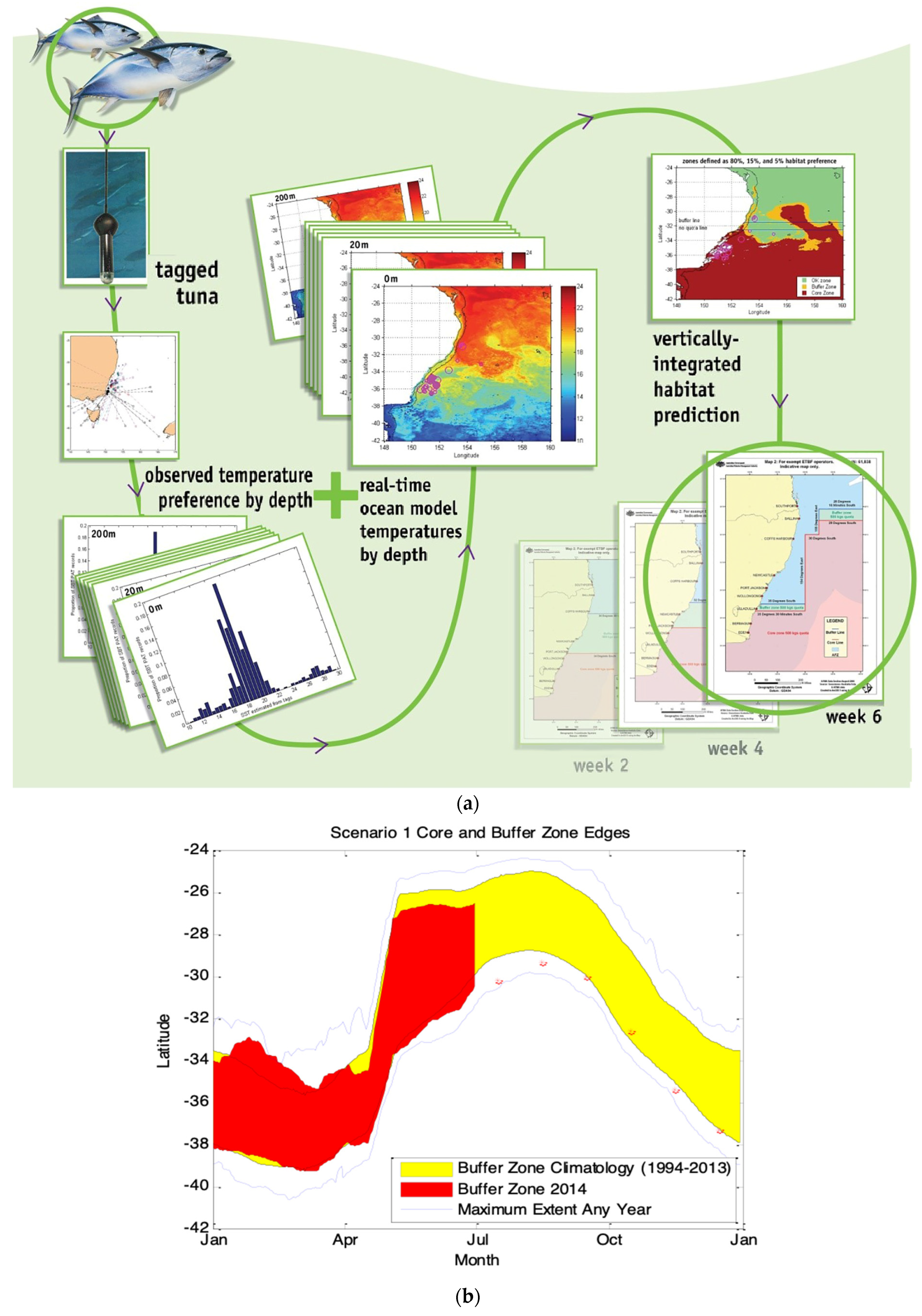
2.1.1. Case Study 1—Southern Bluefin Tuna in the Great Australia Bight
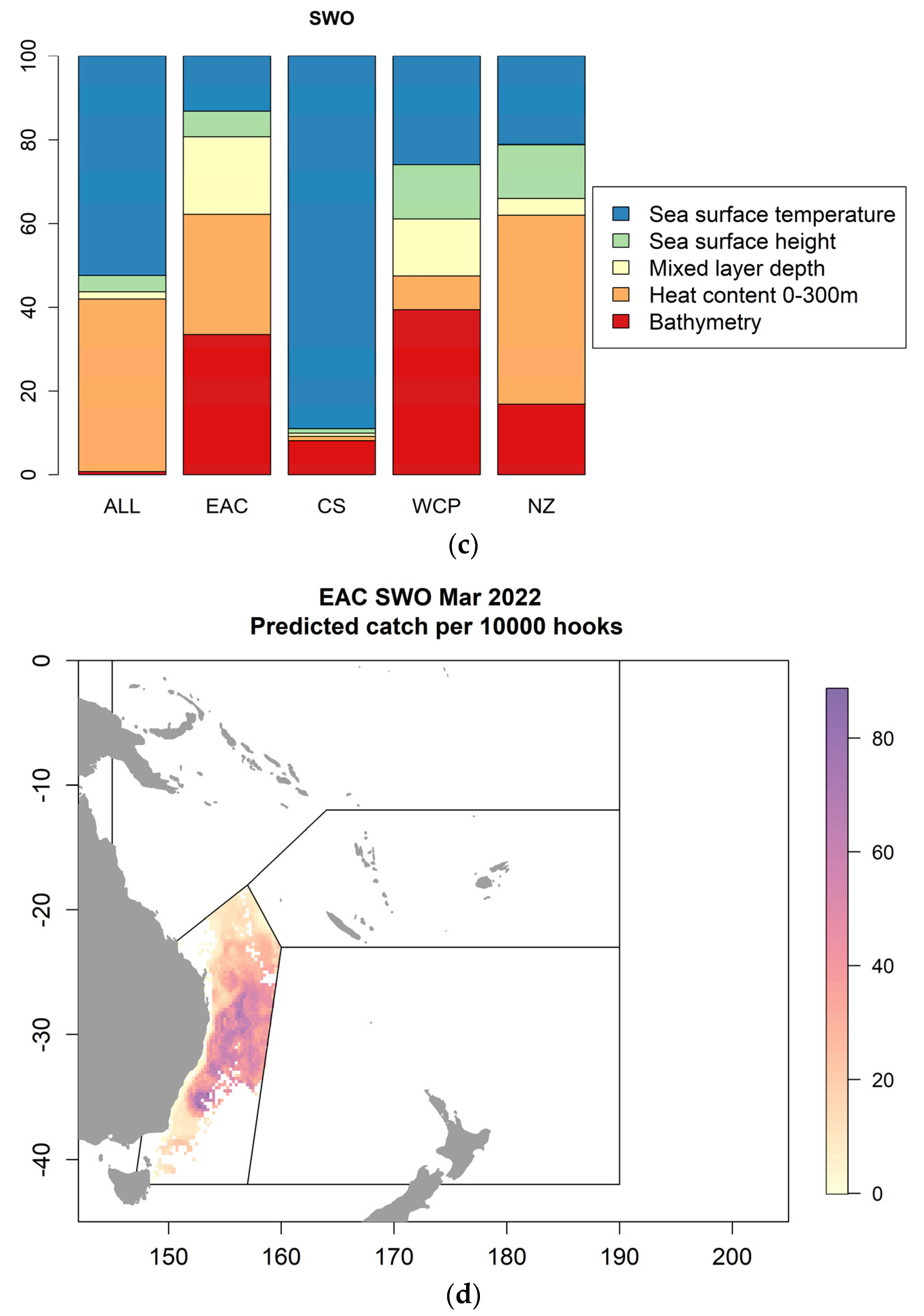
2.1.2. Case Study 2—Forecasts in a Multispecies Longline Fishery in Eastern Australia
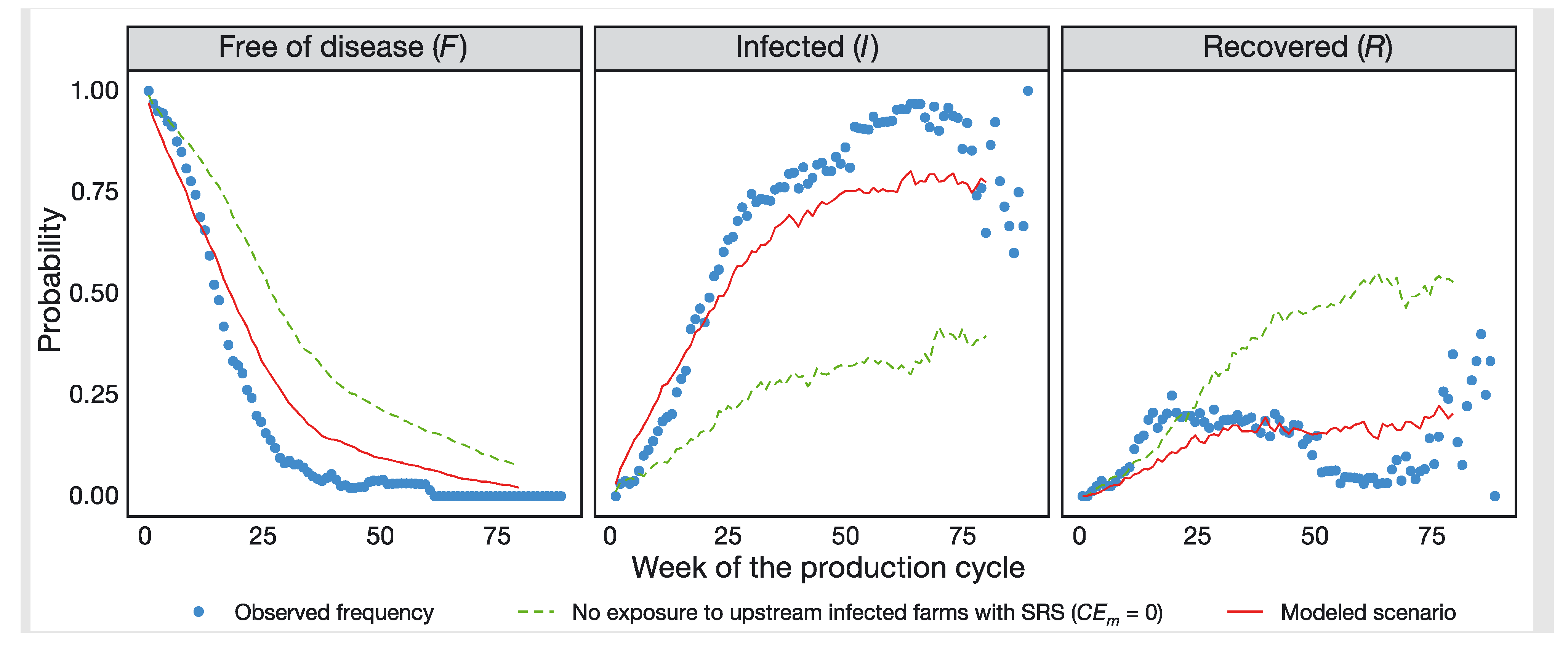
2.2. Ecological Forecasts in Aquaculture
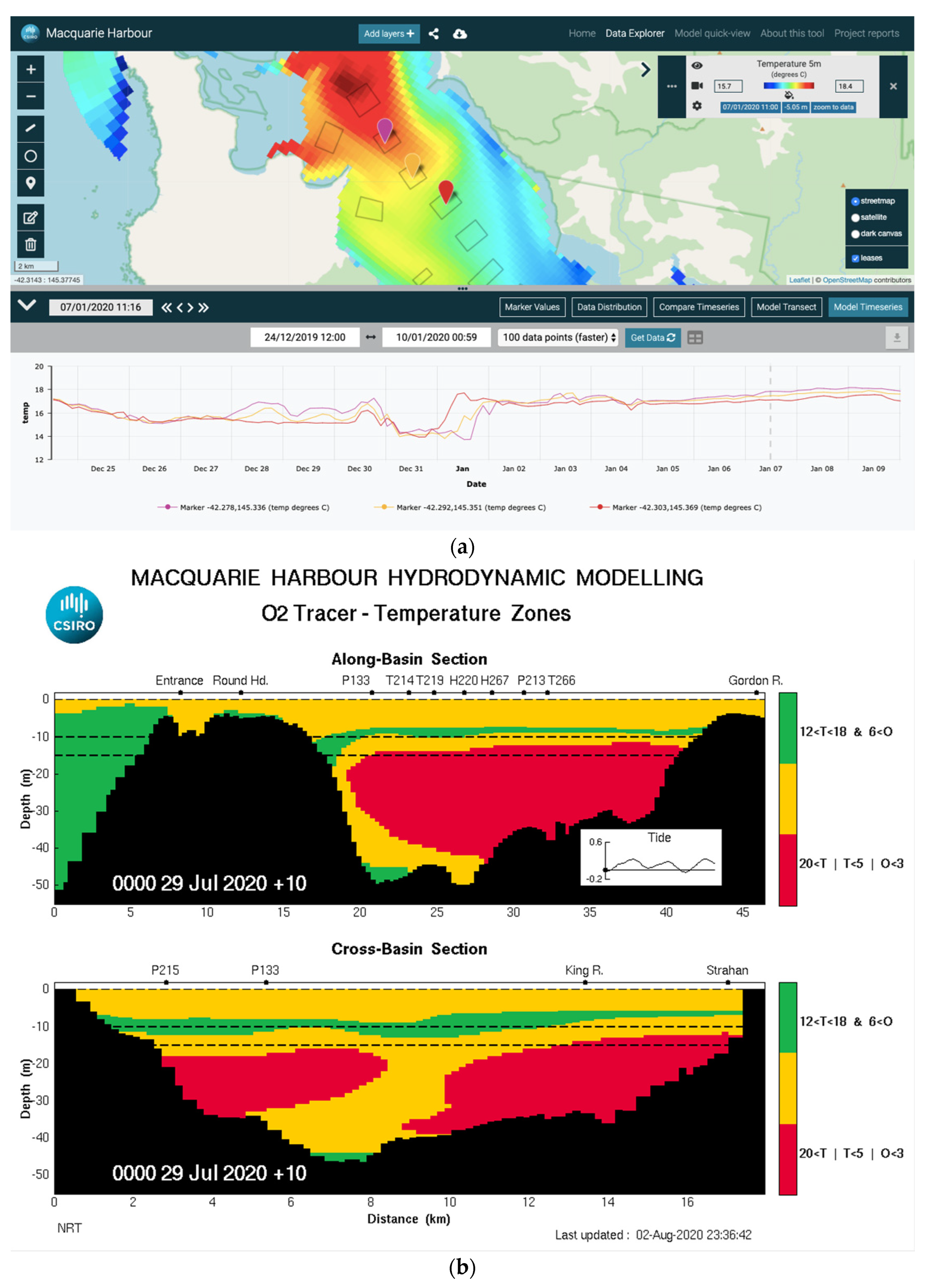
2.2.1. Case Study 1—Dissolved Oxygen Forecasts in Tasmania
2.2.2. Case Study 2—An Operational Information System for Managing the Chilean Aquaculture Industry
2.3. Ecological Forecasts of Harmful Algal Blooms
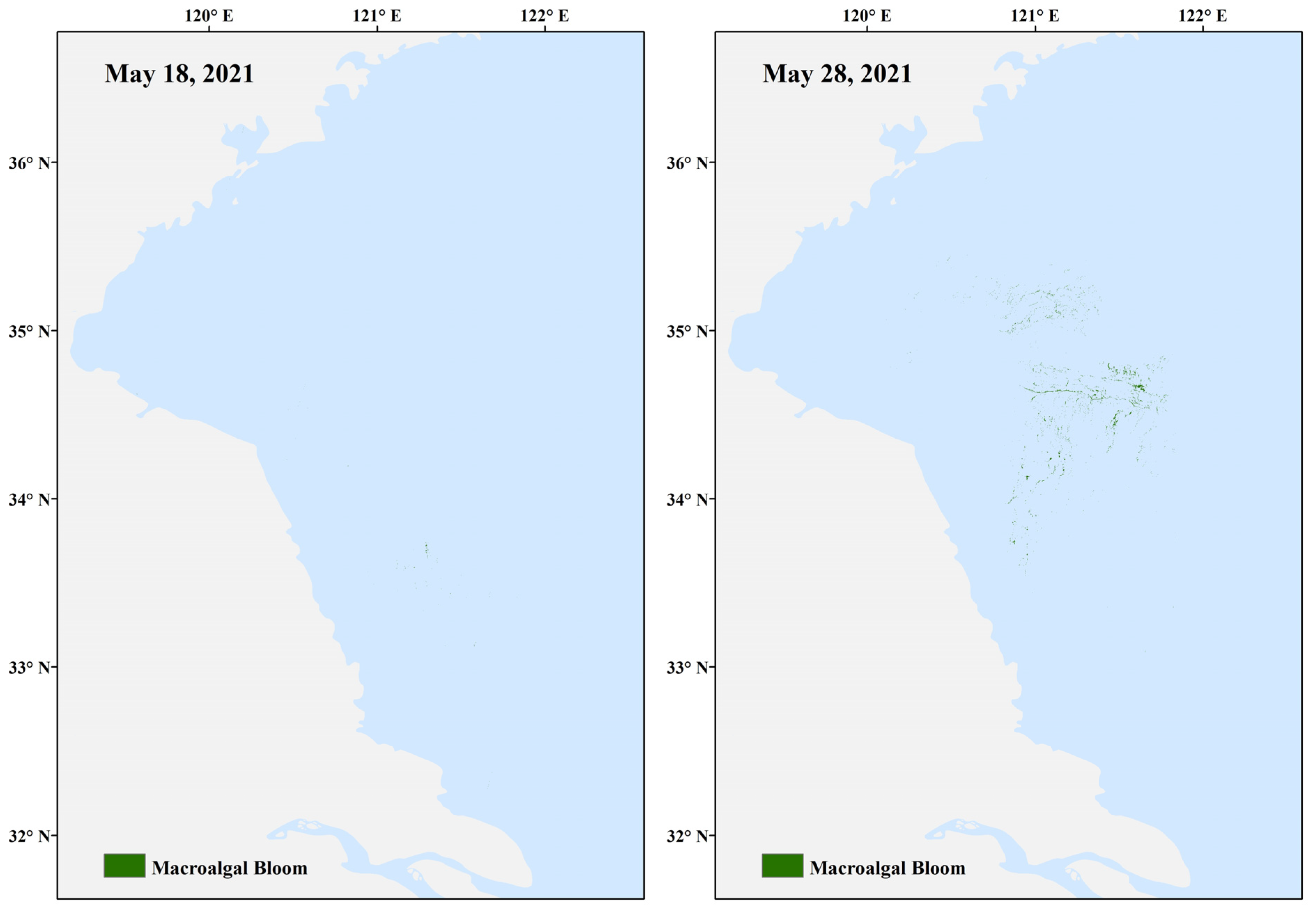
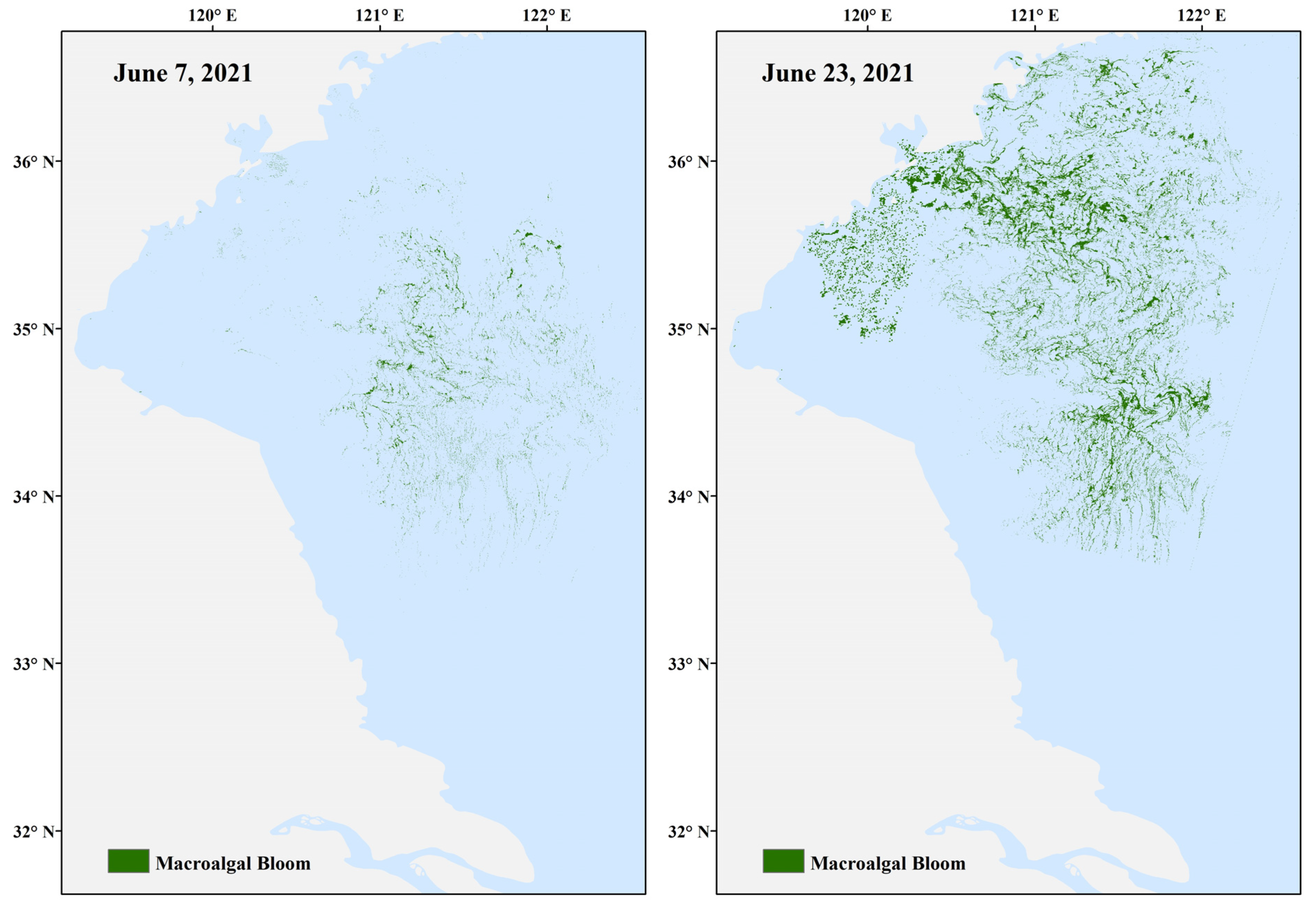
2.3.1. Case Study 1—Real-Time Forecasting of Harmful Algal Blooms in the Yellow SEA
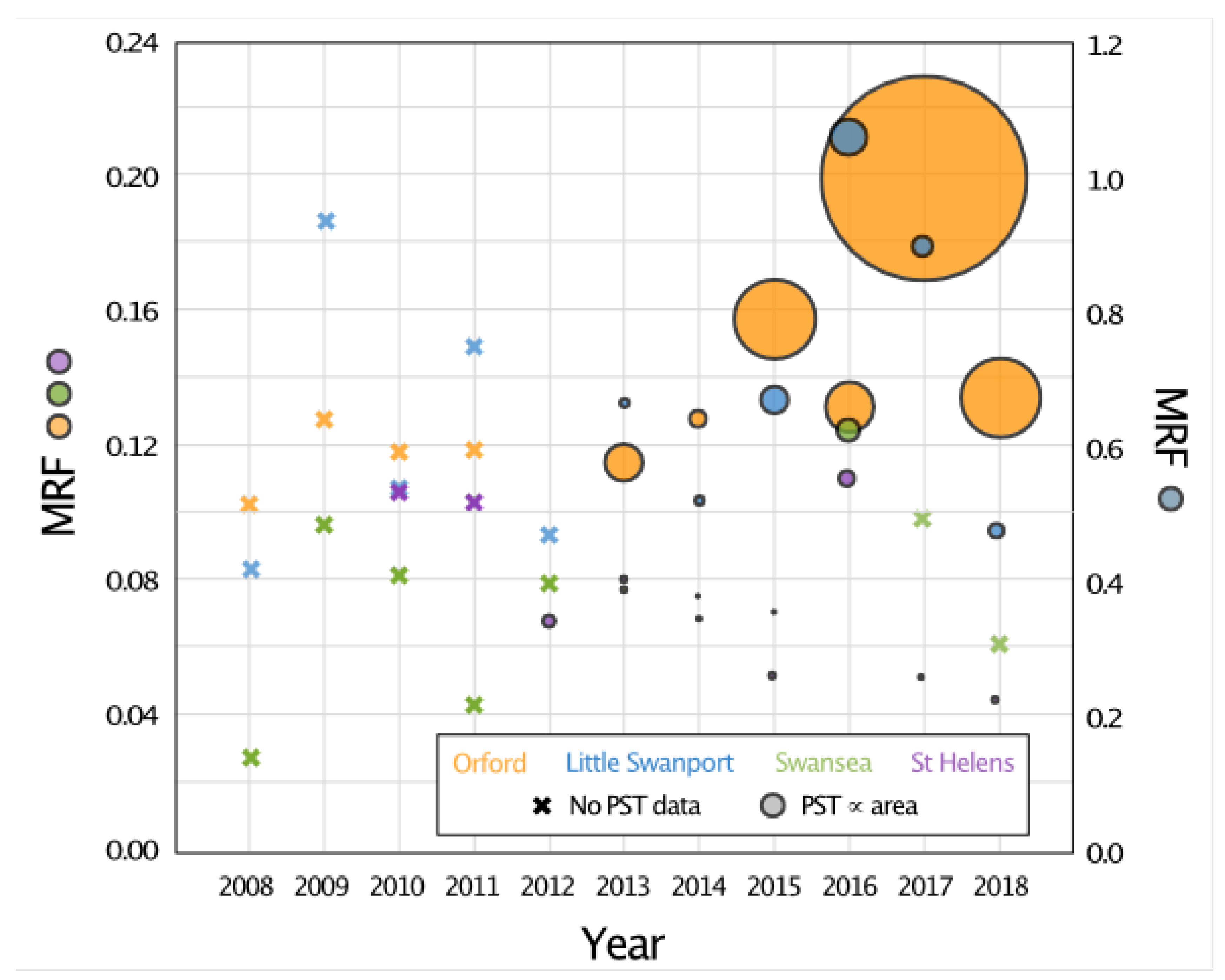
2.3.2. Case Study 2—Toxic Algal Blooms in Tasmanian Coastal Waters
2.4. Ecological Forecasting for Risks to Iconic Habitats or Their Users
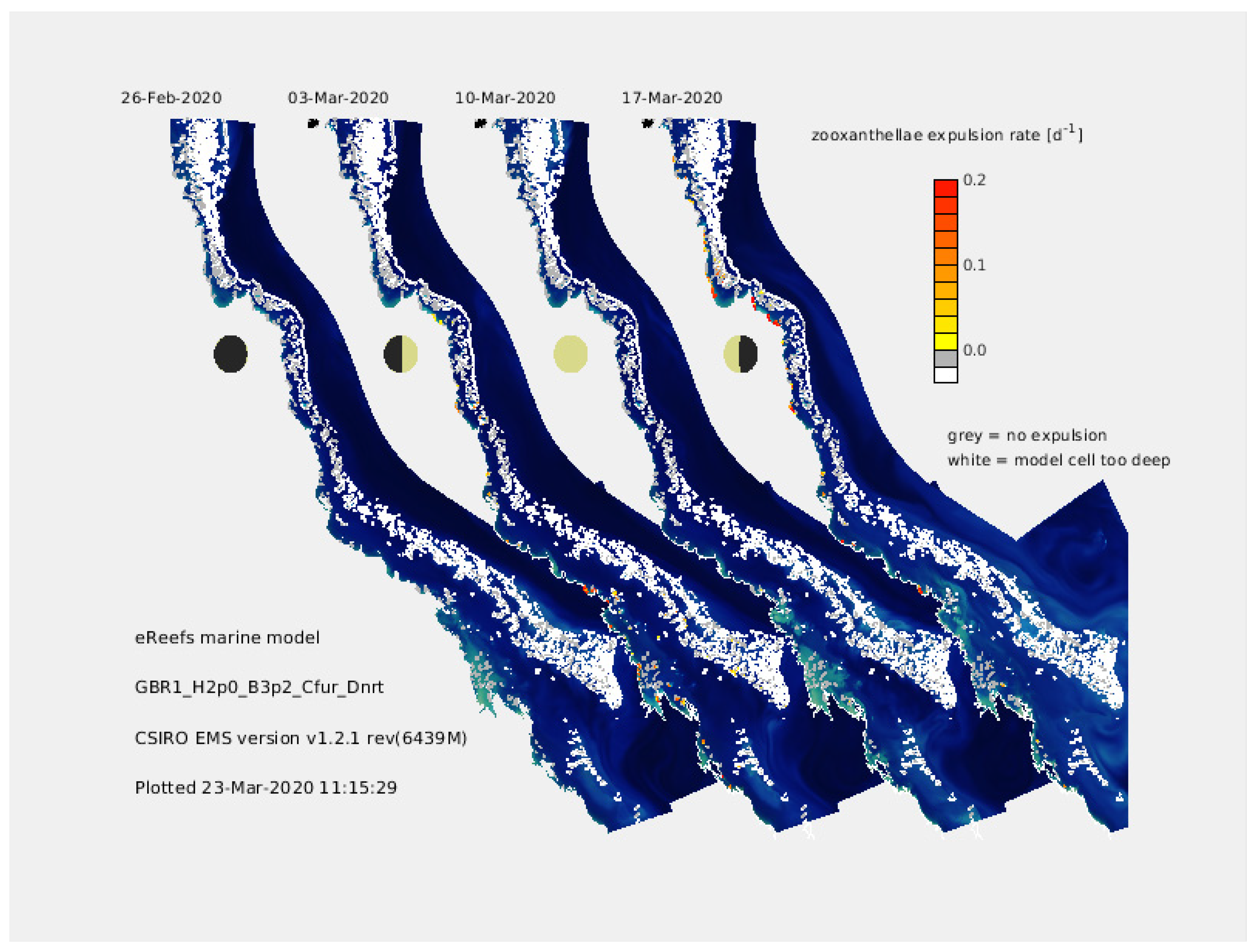
2.4.1. Case Study 1—Near Real-Time Forecasts of Coral Bleaching on the Great Barrier Reef
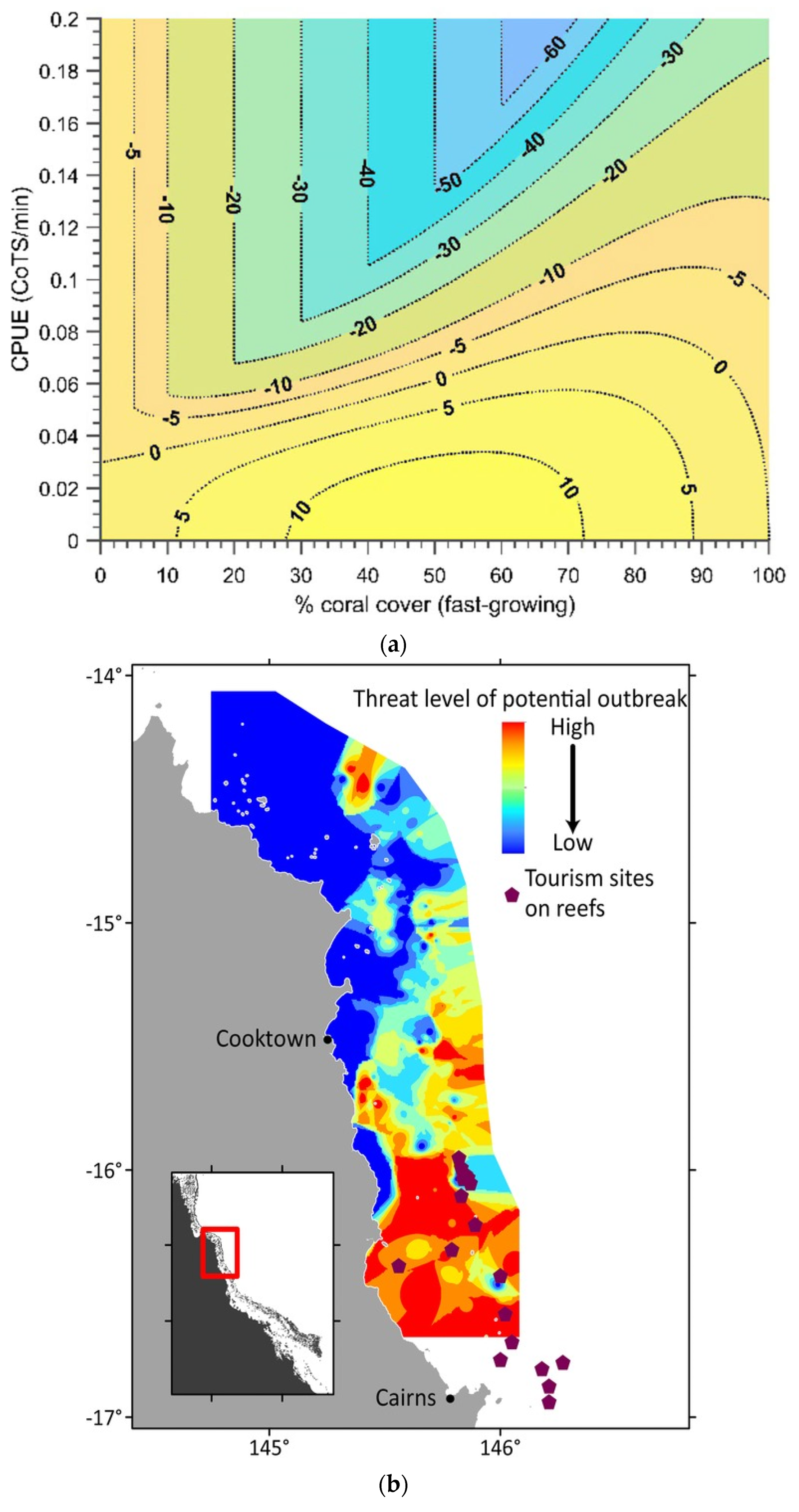
2.4.2. Case Study 2—Forecasting to Inform Suppression of Crown-of-Thorns Starfish on the Great Barrier Reef
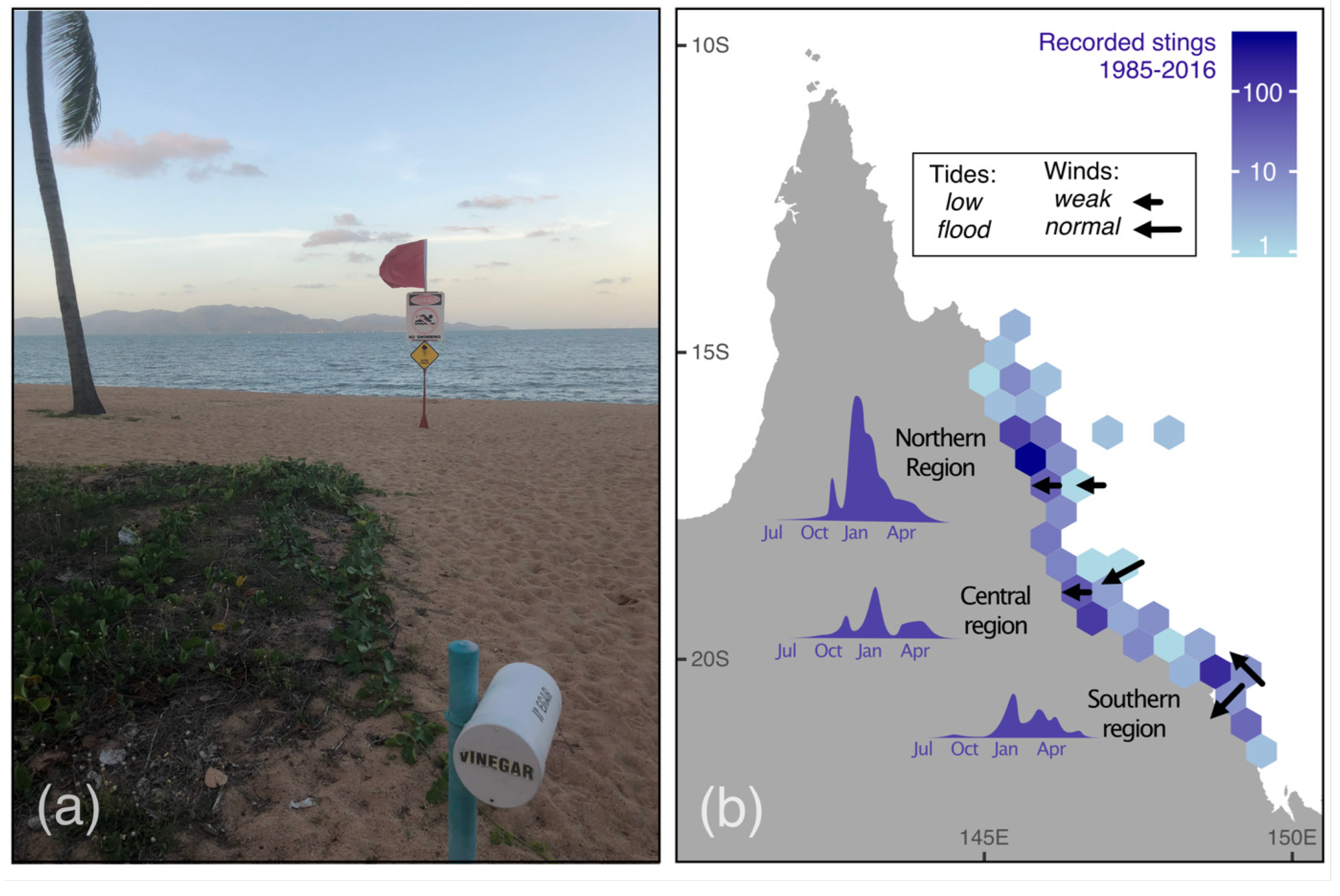
2.4.3. Case Study 3—Real-Time Forecasting to Manage Risks Posed by Irukandji Jellyfish on the Great Barrier Reef
2.5. Common Features amongst Case Studies
3. Discussion
3.1. Insights from the Case Studies
3.1.1. A Clearly Identified Management Problem Benefits from Environmental Intelligence
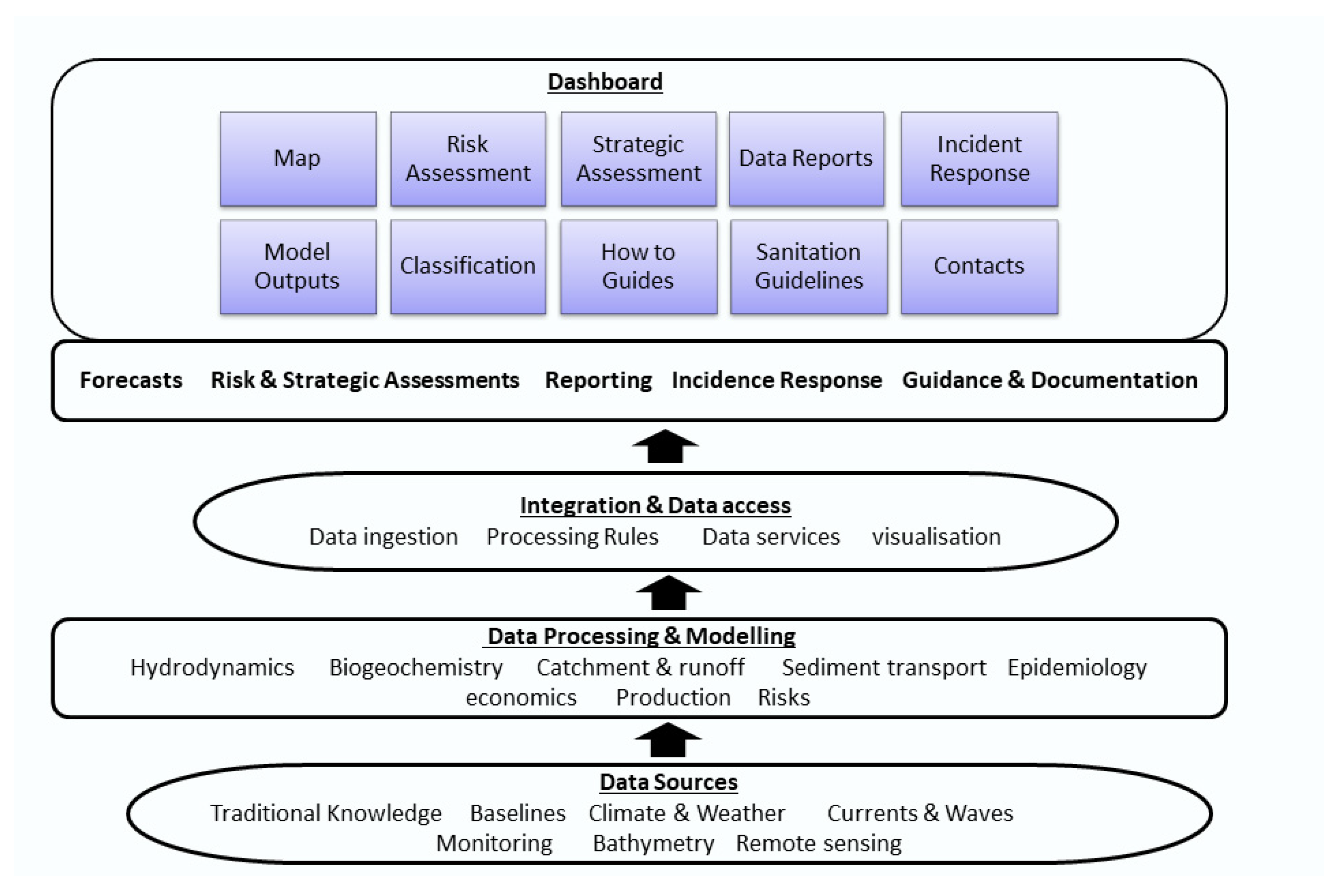
3.1.2. Operational Information Systems can Enhance Forecast Value
3.1.3. An Enduring Funding Model Is Needed to Sustain and Develop the Field
3.2. Gaps and Improvements
3.3. Recommendations
- Information systems can help end users. To integrate ecoforecasting into decision making, an operational information system that synthesises observations and modelling to provide easy-access information delivered through cyberinfrastructure can be used as a powerful tool to communicate a complex forecast. Not all forecasts need to be developed by mechanism-based models, which tend to be more expensive to run. Sometimes empirical models relying on correlations of past events (which can be run very quickly) can be effective. The trade-off between these two types of models needs to be considered for optimal outcome. Similarly, not all forecasts need to be delivered by operational information systems, and in some situations, a simple alert or warning might be sufficient. However, in complex situations or when the model results need expert interpretation, an easy-to-use information system will be essential. Significant and ongoing investment in cyberinfrastructure is needed to support operational forecasting systems.
- Active engagement with end users is essential to ensure forecasts are reliable and useful, with their assumptions, uncertainties, and results clearly communicated, and the needs of decision makers addressed [16,23]. Effective partnerships should be formed between scientists and stakeholders. These could aim to develop effective communication tools that recognise stakeholders’ level of forecasting knowledge, priorities, and interests related to the forecast [39]. Ethical issues associated with forecasting should be considered to allow societal, ecological, and economic benefits (e.g., [15]).
- National ecoforecasting agencies are best able to support long-term delivery. The project-based funding model needs to be backed by strategic funding or commercial investment to execute ongoing delivery and operationalisation. Research projects and teams have delivered excellent forecast systems, but dedicated national programs to provide marine ecoforecasts (e.g., [24]) are needed to bring together scientists and resource managers together to solve resource management challenges in a rapidly changing world, and deliver consistent, timely, and reliable forecasts to a wide range of users. The Ecological Forecasting Initiative is a grass roots coordination approach connecting forecast developers in the USA, Canada, and Oceania regions, but is still supported by project-based funding. We proposed that funding agencies consider supporting a national agency responsible for coordinating existing monitoring, modelling, and dissemination capabilities for nationally important priority areas of ecoforecasting.
- Real-time data access will require new technologies. New technologies need to be developed to provide real-time in situ observation data and fit-for-purpose models (e.g., hydrodynamic, ecological, disease). For example, DNA-based techniques and data could inform ecological models, especially when cryptic, sporadic, remote, organisms are involved.
3.4. The Future of Forecasts in Supporting Sustainable Ocean Management
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hobday, A.J.; Pecl, G.T. Identification of global marine hotspots: Sentinels for change and vanguards for adaptation action. Rev. Fish Biol. Fish. 2014, 24, 415–425. [Google Scholar] [CrossRef]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Sen Gupta, A.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. 2019, 9, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Smith, R.D.; Brown, C.J.; Ceccarelli, D.M.; Edgar, G.J. Ecosystem restructuring along the Great Barrier Reef following mass coral bleaching. Nature 2018, 560, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babcock, R.C.; Bustamante, R.H.; Fulton, E.A.; Fulton, D.J.; Haywood, M.D.E.; Hobday, A.J.; Kenyon, R.; Matear, R.J.; Plagányi, E.E.; Richardson, A.J.; et al. Severe Continental-Scale Impacts of Climate Change Are Happening Now: Extreme Climate Events Impact Marine Habitat Forming Communities Along 45% of Australia’s Coast. Front. Mar. Sci. 2019, 6, 411. [Google Scholar] [CrossRef]
- Wernberg, T.; Russell, B.D.; Thomsen, M.S.; Gurgel, C.F.D.; Bradshaw, C.J.; Poloczanska, E.S.; Connell, S.D. Seaweed Communities in Retreat from Ocean Warming. Curr. Biol. 2011, 21, 1828–1832. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Stuart-Smith, R.D. Climate change: Large-scale abundance shifts in fishes. Curr. Biol. 2021, 31, R1445–R1447. [Google Scholar] [CrossRef]
- Halpern, B.S.; Frazier, M.; Afflerbach, J.; Lowndes, J.S.; Micheli, F.; O’Hara, C.; Scarborough, C.; Selkoe, K.A. Recent pace of change in human impact on the world’s ocean. Sci. Rep. 2019, 9, 11609. [Google Scholar] [CrossRef] [Green Version]
- Dietze, M.C.; Fox, A.; Beck-Johnson, L.M.; Betancourt, J.L.; Hooten, M.B.; Jarnevich, C.S.; Keitt, T.H.; Kenney, M.A.; Laney, C.M.; Larsen, L.G.; et al. Iterative near-term ecological forecasting: Needs, opportunities, and challenges. Proc. Natl. Acad. Sci. USA 2018, 115, 1424–1432. [Google Scholar] [CrossRef]
- Tulloch, A.I.T.; Hagger, V.; Greenville, A.C. Ecological forecasts to inform near-term management of threats to biodiversity. Glob. Chang. Biol. 2020, 26, 5816–5828. [Google Scholar] [CrossRef] [PubMed]
- Hobday, A.J.; Spillman, C.M.; Eveson, J.P.; Hartog, J.R.; Zhang, X.; Brodie, S. A Framework for Combining Seasonal Forecasts and Climate Projections to Aid Risk Management for Fisheries and Aquaculture. Front. Mar. Sci. 2018, 5, 137. [Google Scholar] [CrossRef] [Green Version]
- Dietze, M.C. Ecological Forcasting; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Tommasi, D.; Stock, C.A.; Hobday, A.J.; Methot, R.; Kaplan, I.C.; Eveson, J.P.; Holsman, K.; Miller, T.J.; Gaichas, S.; Gehlen, M.; et al. Managing living marine resources in a dynamic environment: The role of seasonal to decadal climate forecasts. Prog. Oceanogr. 2017, 152, 15–49. [Google Scholar] [CrossRef] [Green Version]
- Hobday, A.J.; Hartog, J.R.; Manderson, J.P.; Mills, K.E.; Oliver, M.J.; Pershing, A.J.; Siedlecki, S. Ethical considerations and unanticipated consequences associated with ecological forecasting for marine resources. ICES J. Mar. Sci. 2019, 76, 1244–1256. [Google Scholar] [CrossRef]
- Bodner, K.; Rauen Firkowski, C.; Bennett, J.R.; Brookson, C.; Dietze, M.; Green, S.; Hughes, J.; Kerr, J.; Kunegel-Lion, M.; Leroux, S.J.; et al. Bridging the divide between ecological forecasts and environmental decision making. Ecosphere 2021, 12, e03869. [Google Scholar] [CrossRef]
- Lagabrielle, E.; Allibert, A.; Kiszka, J.J.; Loiseau, N.; Kilfoil, J.P.; Lemahieu, A. Environmental and anthropogenic factors affecting the increasing occurrence of shark-human interactions around a fast-developing Indian Ocean island. Sci. Rep. 2018, 8, 3676. [Google Scholar] [CrossRef] [Green Version]
- Powell, G.; Versluys, T.M.M.; Williams, J.J.; Tiedt, S.; Pooley, S. Using environmental niche modelling to investigate abiotic predictors of crocodilian attacks on people. Oryx 2020, 54, 639–647. [Google Scholar] [CrossRef]
- Rowley, O.C.; Courtney, R.; Northfield, T.; Seymour, J. Environmental drivers of the occurrence and abundance of the Irukandji jellyfish (Carukia barnesi). PLoS ONE 2022, 17, e0272359. [Google Scholar] [CrossRef]
- HOBDAY, A.J.; HARTMANN, K. Near real-time spatial management based on habitat predictions for a longline bycatch species. Fish. Manag. Ecol. 2006, 13, 365–380. [Google Scholar] [CrossRef]
- Howell, E.; Kobayashi, D.; Parker, D.; Balazs, G.; Polovina, J. TurtleWatch: A tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. Endanger. Species Res. 2008, 5, 267–278. [Google Scholar] [CrossRef]
- Payne, M.R.; Hobday, A.J.; MacKenzie, B.R.; Tommasi, D.; Dempsey, D.P.; Fässler, S.M.M.; Haynie, A.C.; Ji, R.; Liu, G.; Lynch, P.D.; et al. Lessons from the First Generation of Marine Ecological Forecast Products. Front. Mar. Sci. 2017, 4, 289. [Google Scholar] [CrossRef]
- Hobday, A.J.; Spillman, C.M.; Paige Eveson, J.; Hartog, J.R. Seasonal forecasting for decision support in marine fisheries and aquaculture. Fish. Oceanogr. 2016, 25, 45–56. [Google Scholar] [CrossRef]
- NOAA (National Oceanic and Atmospheric Administration). A Strategic Vision for NOAA’s Ecological Forecasting Roadmap, 2015–2019; NOAA: Silver Spring, MD, USA, 2015.
- Hazen, E.L.; Palacios, D.M.; Forney, K.A.; Howell, E.A.; Becker, E.; Hoover, A.L.; Irvine, L.; DeAngelis, M.; Bograd, S.J.; Mate, B.R.; et al. WhaleWatch: A dynamic management tool for predicting blue whale density in the California Current. J. Appl. Ecol. 2017, 54, 1415–1428. [Google Scholar] [CrossRef]
- Malick, M.J.; Siedlecki, S.A.; Norton, E.L.; Kaplan, I.C.; Haltuch, M.A.; Hunsicker, M.E.; Parker-Stetter, S.L.; Marshall, K.N.; Berger, A.M.; Hermann, A.J.; et al. Environmentally Driven Seasonal Forecasts of Pacific Hake Distribution. Front. Mar. Sci. 2020, 7, 578490. [Google Scholar] [CrossRef]
- Payne, M.R.; Hobday, A.J.; MacKenzie, B.R.; Tommasi, D. Editorial: Seasonal-to-Decadal Prediction of Marine Ecosystems: Opportunities, Approaches, and Applications. Front. Mar. Sci. 2019, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.S.L.; Rollinson, C.R.; Allyn, A.J.; Ashander, J.; Brodie, S.; Brookson, C.B.; Collins, E.; Dietze, M.C.; Gallinat, A.S.; Juvigny-Khenafou, N.; et al. The power of forecasts to advance ecological theory. Methods Ecol. Evol. 2022. [Google Scholar] [CrossRef]
- Hilborn, R.; Walters, C.J. Quantitative Fisheries Stock Assessment; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Steven, A.D.L.; Aryal, S.; Bernal, P.; Bravo, F.; Bustamante, R.H.; Condie, S.; Dambacher, J.M.; Dowideit, S.; Fulton, E.A.; Gorton, R.; et al. SIMA Austral: An operational information system for managing the Chilean aquaculture industry with international application. J. Oper. Oceanogr. 2019, 12 (Suppl. 2), S29–S46. [Google Scholar] [CrossRef] [Green Version]
- Steven, A.D.L.; Baird, M.E.; Brinkman, R.; Car, N.J.; Cox, S.J.; Herzfeld, M.; Hodge, J.; Jones, E.; King, E.; Margvelashvili, N.; et al. eReefs: An operational information system for managing the Great Barrier Reef. J. Oper. Oceanogr. 2019, 12 (Suppl. 2), S12–S28. [Google Scholar] [CrossRef] [Green Version]
- Capotondi, A.; Jacox, M.; Bowler, C.; Kavanaugh, M.; Lehodey, P.; Barrie, D.; Brodie, S.; Chaffron, S.; Cheng, W.; Dias, D.F.; et al. Observational Needs Supporting Marine Ecosystems Modeling and Forecasting: From the Global Ocean to Regional and Coastal Systems. Front. Mar. Sci. 2019, 6, 623. [Google Scholar] [CrossRef] [Green Version]
- Stock, C.A.; Alexander, M.A.; Bond, N.A.; Brander, K.M.; Cheung, W.W.L.; Curchitser, E.N.; Delworth, T.L.; Dunne, J.P.; Griffies, S.M.; Haltuch, M.A.; et al. On the use of IPCC-class models to assess the impact of climate on Living Marine Resources. Prog. Oceanogr. 2011, 88, 1–27. [Google Scholar] [CrossRef]
- Hicks, C.C.; Cohen, P.J.; Graham, N.A.J.; Nash, K.L.; Allison, E.H.; D’Lima, C.; Mills, D.J.; Roscher, M.; Thilsted, S.H.; Thorne-Lyman, A.L.; et al. Harnessing global fisheries to tackle micronutrient deficiencies. Nature 2019, 574, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiault, L.; Mora, C.; Cinner, J.E.; Cheung, W.W.L.; Graham, N.A.J.; Januchowski-Hartley, F.A.; Mouillot, D.; Sumaila, U.R.; Claudet, J. Escaping the perfect storm of simultaneous climate change impacts on agriculture and marine fisheries. Sci. Adv. 2019, 5, eaaw9976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.A.; Kay, S.; Hossain, M.A.R.; Ahmed, M.; Cheung, W.W.L.; Lazar, A.N.; Barange, M. Projecting marine fish production and catch potential in Bangladesh in the 21st century under long-term environmental change and management scenarios. ICES J. Mar. Sci. 2015, 73, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Basson, M.; Hobday, A.J.; Eveson, J.P.; Patterson, T.A. Spatial Interactions among Juvenile Southern Bluefin Tuna at the Global Scale: A Large Scale Archival Tag Experiment; Final Report to the Australian Fisheries Research and Development Corporation, FRDC Project No. 2003/002; CSIRO: Canberra, Australia, 2012.
- Eveson, J.; Hobday, A.; Hartog, J.; Spillman, C.; Rough, K. Forecasting Spatial Distribution of Southern Bluefin Tuna Habitat in the Great Australian Bight; FRDC Final report, FRDC Project No. 2012-239; Fisheries Research and Development Corporation: Canberra, Australia, 2014. [Google Scholar]
- Eveson, J.P.; Hobday, A.J.; Hartog, J.R.; Spillman, C.M.; Rough, K.M. Seasonal forecasting of tuna habitat in the Great Australian Bight. Fish. Res. 2015, 170, 39–49. [Google Scholar] [CrossRef]
- Eveson, J.P.; Hartog, J.; Spillman, C.; Rough, K. Forecasting Spatial Distribution of Southern Bluefin Tuna Habitat in the Great Australian Bight—Updating and Improving Habitat and Forecast Models. Final Report, FRDC Project No. 2018-194. 2021. Available online: https://www.frdc.com.au/sites/default/files/products/2018-194-DLD.pdf (accessed on 1 December 2022).
- Hobday, A.J.; Hartog, J.R.; Timmiss, T.; Fielding, J. Dynamic spatial zoning to manage southern bluefin tuna (Thunnus maccoyii) capture in a multi-species longline fishery. Fish. Oceanogr. 2010, 19, 243–253. [Google Scholar] [CrossRef]
- Scales, K.L.; Moore, T.S.; Sloyan, B.; Spillman, C.M.; Eveson, J.P.; Patterson, T.A.; Williams, A.J.; Hobday, A.J.; Hartog, J.R. Forecast-ready models to support fisheries adaptation to global variability and change. Fish. Oceanogr. 2022, submitted.
- FAO. The State of World Fisheries and Aquaculture (SOFIA); FAO: Rome, Italy, 2020. [Google Scholar]
- Quiñones, R.A.; Fuentes, M.; Montes, R.M.; Soto, D.; León-Muñoz, J. Environmental issues in Chilean salmon farming: A review. Rev. Aquac. 2019, 11, 375–402. [Google Scholar] [CrossRef]
- Steven, A.D.L. Managing the societal uses of phytoplankton: Technology applications and needs. In Advances in Phytoplankton Ecology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 265–297. [Google Scholar]
- Cullen-Knox, C.; Fleming, A.; Lester, L.; Ogier, E. Publicised scrutiny and mediatised environmental conflict: The case of Tasmanian salmon aquaculture. Mar. Policy 2019, 100, 307–315. [Google Scholar] [CrossRef]
- Wild-Allen, K.; Andrewartha, J.; Baird, M.; Bodrossy, L.; Brewer, E.; Eriksen, R.; Skerratt, J.; Revill, A.; Sherrin, K.; Wild, D. Macquarie Harbour Oxygen Process Model; CSIRO Oceans & Atmosphere: Hobart, Australia, 2020.
- Chávez, C.; Dresdner, J.; Figueroa, Y.; Quiroga, M. Main issues and challenges for sustainable development of salmon farming in Chile: A socio-economic perspective. Rev. Aquac. 2019, 11, 403–421. [Google Scholar] [CrossRef] [Green Version]
- Fulton, E.A. Appendix C Integrated system model content. In SIMA Austral. FIE V008-Integrated Ecosystem-Based Sanitary and Environmental Management System for Aquaculture. Final Report; Steven, A.D.L., Andrewartha, J.R., Bernal, P.A., Bravo, F., Bustamante, R.H., Condie, S.A., Crane, M., Dambacher, J.M., Dowideit, S., Eds.; CSIRO: Brisbane, Australia, 2018; p. 178. [Google Scholar]
- Bravo, F.; Bustamante, R.H.E.F. Appendix J: Semi-Qualitative Risk Assessment at Marine Salmon Farm Sites; CSIRO: Brisbane, Australia, 2018.
- Kudela, R.M.; Berdalet, E.; Enevoldsen, H.; Pitcher, G.; Raine, R.; Urban, E. GEOHAB—The Global Ecology and Oceanography of Harmful Algal Blooms Program: Motivation, Goals, and Legacy. Oceanography 2017, 30, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Bottein, M.-Y.D.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H.; et al. Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts. Commun. Earth Environ. 2021, 2, 117. [Google Scholar] [CrossRef]
- Stauffer, B.A.; Bowers, H.A.; Buckley, E.; Davis, T.W.; Johengen, T.H.; Kudela, R.; McManus, M.A.; Purcell, H.; Smith, G.J.; Vander Woude, A.; et al. Considerations in Harmful Algal Bloom Research and Monitoring: Perspectives From a Consensus-Building Workshop and Technology Testing. Front. Mar. Sci. 2019, 6, 399. [Google Scholar] [CrossRef] [Green Version]
- Davidson, K.; Whyte, C.; Aleynik, D.; Dale, A.; Gontarek, S.; Kurekin, A.A.; McNeill, S.; Miller, P.I.; Porter, M.; Saxon, R.; et al. HABreports: Online Early Warning of Harmful Algal and Biotoxin Risk for the Scottish Shellfish and Finfish Aquaculture Industries. Front. Mar. Sci. 2021, 8, 631732. [Google Scholar] [CrossRef]
- He, Z.; Yang, D.; Wang, Y.; Yin, B. Impact of 4D-Var data assimilation on modelling of the East China Sea dynamics. Ocean. Model. 2022, 176, 102044. [Google Scholar] [CrossRef]
- Yang, D.; Yin, B.; Chai, F.; Feng, X.; Xue, H.; Gao, G.; Yu, F. The onshore intrusion of Kuroshio subsurface water from February to July and a mechanism for the intrusion variation. Prog. Oceanogr. 2018, 167, 97–115. [Google Scholar] [CrossRef]
- Gao, H. Big Earth Data in Support of the Sustainable Development Goals; Chinese Academy of Sciences: Beijing, China, 2021. Available online: https://www.fmprc.gov.cn/mfa_eng/topics_665678/2030kcxfzyc/202109/P020211019152777729038.pdf (accessed on 1 December 2022).
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in Understanding Harmful Algal Blooms: Paradigm Shifts and New Technologies for Research, Monitoring, and Management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condie, S.A.; Oliver, E.C.J.; Hallegraeff, G.M. Environmental drivers of unprecedented Alexandrium catenella dinoflagellate blooms off eastern Tasmania, 2012–2018. Harmful Algae 2019, 87, 101628. [Google Scholar] [CrossRef]
- Turnbull, A.; Malhi, N.; Seger, A.; Harwood, T.; Jolley, J.; Fitzgibbon, Q.; Hallegraeff, G. Paralytic shellfish toxin uptake, tissue distribution, and depuration in the Southern Rock Lobster Jasus edwardsii Hutton. Harmful Algae 2020, 95, 101818. [Google Scholar] [CrossRef]
- Xie, H.; Fischer, A.M.; Strutton, P.G. Generalized linear models to assess environmental drivers of paralytic shellfish toxin blooms (Southeast Tasmania, Australia). Cont. Shelf Res. 2021, 223, 104439. [Google Scholar] [CrossRef]
- Condie, S.A. Settling regimes for non-motile particles in stratified waters. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 681–699. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.E.; Mongin, M.; Rizwi, F.; Bay, L.K.; Cantin, N.E.; Soja-Woźniak, M.; Skerratt, J. A mechanistic model of coral bleaching due to temperature-mediated light-driven reactive oxygen build-up in zooxanthellae. Ecol. Model. 2018, 386, 20–37. [Google Scholar] [CrossRef]
- Baird, M.E.; Wild-Allen, K.A.; Parslow, J.; Mongin, M.; Robson, B.; Skerratt, J.; Rizwi, F.; Soja-Woźniak, M.; Jones, E.; Herzfeld, M.; et al. CSIRO Environmental Modelling Suite (EMS): Scientific description of the optical and biogeochemical models (vB3p0). Geosci. Model Dev. 2020, 13, 4503–4553. [Google Scholar] [CrossRef]
- Baird, M.E.; Mongin, M.; Rizwi, F.; Bay, L.K.; Cantin, N.E.; Morris, L.A.; Skerratt, J. The effect of natural and anthropogenic nutrient and sediment loads on coral oxidative stress on runoff-exposed reefs. Mar. Pollut. Bull. 2021, 168, 112409. [Google Scholar] [CrossRef] [PubMed]
- Condie, S.A.; Anthony, K.R.N.; Babcock, R.C.; Baird, M.E.; Beeden, R.; Fletcher, C.S.; Gorton, R.; Harrison, D.; Hobday, A.J.; Plagányi, É.E.; et al. Large-scale interventions may delay decline of the Great Barrier Reef. R. Soc. Open Sci. 2021, 8, 201296. [Google Scholar] [CrossRef] [PubMed]
- Heron, S.F.; Eakin, C.M.; Douvere, F.; Anderson, K.; Day, J.C.; Geiger, E.; Hoegh-Guldberg, O.; van Hooidonk, R.; Hughes, T.; Marshall, P.; et al. Impacts of Climate Change on World Heritage Coral Reefs: A First Global Scientific Assessment; UNESCO World Heritage Centre: Paris, France, 2017. [Google Scholar]
- Pratchett, M.S.; Caballes, C.F.; Rivera-Posada, J.A.; Sweatman, H.P.A. Limits to understanding and managing outbreaks of crown-of-thorns starfish (Acanthaster spp.). Oceanogr. Mar. Biol. Annu. Rev. 2014, 52, 133–200. [Google Scholar]
- Mellin, C.; Matthews, S.; Anthony, K.R.N.; Brown, S.C.; Caley, M.J.; Johns, K.A.; Osborne, K.; Puotinen, M.; Thompson, A.; Wolff, N.H.; et al. Spatial resilience of the Great Barrier Reef under cumulative disturbance impacts. Glob. Chang. Biol. 2019, 25, 2431–2445. [Google Scholar] [CrossRef]
- Osborne, K.; Dolman, A.M.; Burgess, S.C.; Johns, K.A. Disturbance and the Dynamics of Coral Cover on the Great Barrier Reef (1995–2009). PLoS ONE 2011, 6, e17516. [Google Scholar] [CrossRef] [Green Version]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27–year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [Green Version]
- Pratchett, M.S.; Caballes, C.F.; Wilmes, J.C.; Matthews, S.; Mellin, C.; Sweatman, H.P.A.; Nadler, L.E.; Brodie, J.; Thompson, C.A.; Hoey, J.; et al. Thirty Years of Research on Crown-of-Thorns Starfish (1986–2016): Scientific Advances and Emerging Opportunities. Diversity 2017, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Westcott, D.A.; Fletcher, C.S.; Kroon, F.J.; Babcock, R.C.; Plagányi, E.E.; Pratchett, M.S.; Bonin, M.C. Relative efficacy of three approaches to mitigate Crown-of-Thorns Starfish outbreaks on Australia’s Great Barrier Reef. Sci. Rep. 2020, 10, 12594. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, G. A management approach to the COTS question. In The Possible Cause and Consequences of Outbreaks of the Crown-of-Thorns Starfish; Great Barrier Reef Marine Park Authority: Townsville, Australia, 1993; pp. 157–160. [Google Scholar]
- Plagányi, É.E.; Babcock, R.C.; Rogers, J.; Bonin, M.; Morello, E.B. Ecological analyses to inform management targets for the culling of crown-of-thorns starfish to prevent coral decline. Coral Reefs 2020, 39, 1483–1499. [Google Scholar] [CrossRef]
- Hock, K.; Wolff, N.H.; Ortiz, J.C.; Condie, S.A.; Anthony, K.R.N.; Blackwell, P.G.; Mumby, P.J. Connectivity and systemic resilience of the Great Barrier Reef. PLoS Biol. 2017, 15, e2003355. [Google Scholar] [CrossRef] [PubMed]
- Condie, S.A.; Plagányi, É.E.; Morello, E.B.; Hock, K.; Beeden, R. Great Barrier Reef recovery through multiple interventions. Conserv. Biol. 2018, 32, 1356–1367. [Google Scholar] [CrossRef]
- Hock, K.; Wolff, N.H.; Beeden, R.; Hoey, J.; Condie, S.A.; Anthony, K.R.N.; Possingham, H.P.; Mumby, P.J. Controlling range expansion in habitat networks by adaptively targeting source populations. Conserv. Biol. 2016, 30, 856–866. [Google Scholar] [CrossRef]
- Fenner, P.; Carney, I. The Irukandji syndrome. A devastating syndrome caused by a north Australian jellyfish. Aust. Fam. Physician 1999, 28, 1131–1137. [Google Scholar]
- Gershwin, L.-A.; Condie, S.A.; Mansbridge, J.V.; Richardson, A.J. Dangerous jellyfish blooms are predictable. J. R. Soc. Interface 2014, 11, 20131168. [Google Scholar] [CrossRef]
- Gershwin, L.-A.; De Nardi, M.; Winkel, K.D.; Fenner, P.J. Marine Stingers: Review of an Under-Recognized Global Coastal Management Issue. Coast. Manag. 2010, 38, 22–41. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J. Generalized Linear Models, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 1989. [Google Scholar]
- Condie, S.; Condie, R. Retention of plankton within ocean eddies. Glob. Ecol. Biogeogr. 2016, 25, 1264–1277. [Google Scholar] [CrossRef]
- Hock, K.; Wolff, N.H.; Condie, S.A.; Anthony, K.R.N.; Mumby, P.J. Connectivity networks reveal the risks of crown-of-thorns starfish outbreaks on the Great Barrier Reef. J. Appl. Ecol. 2014, 51, 1188–1196. [Google Scholar] [CrossRef]
- Condie, S.A.; Herzfeld, M.; Hock, K.; Andrewartha, J.R.; Gorton, R.; Brinkman, R.; Schultz, M. System level indicators of changing marine connectivity. Ecol. Indic. 2018, 91, 531–541. [Google Scholar] [CrossRef]
- Hock, K.; Doropoulos, C.; Gorton, R.; Condie, S.A.; Mumby, P.J. Split spawning increases robustness of coral larval supply and inter-reef connectivity. Nat. Commun. 2019, 10, 3463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershwin, L.A.; Richardson, A.J.; Winkel, K.D.; Fenner, P.J.; Lippmann, J.; Hore, R.; Avila-Soria, G.; Brewer, D.; Kloser, R.J.; Steven, A.; et al. Biology and ecology of Irukandji jellyfish (Cnidaria: Cubozoa). Adv. Mar. Biol. 2013, 66, 1–85. [Google Scholar] [CrossRef] [PubMed]
- Mouquet, N.; Lagadeuc, Y.; Devictor, V.; Doyen, L.; Duputié, A.; Eveillard, D.; Faure, D.; Garnier, E.; Gimenez, O.; Huneman, P.; et al. REVIEW: Predictive ecology in a changing world. J. Appl. Ecol. 2015, 52, 1293–1310. [Google Scholar] [CrossRef]
- Maina, J.; Kithiia, J.; Cinner, J.; Neale, E.; Noble, S.; Charles, D.; Watson, J.E.M. Integrating social–ecological vulnerability assessments with climate forecasts to improve local climate adaptation planning for coral reef fisheries in Papua New Guinea. Reg. Environ. Chang. 2016, 16, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Cooper, G.S.; Dearing, J.A. Modelling future safe and just operating spaces in regional social-ecological systems. Sci. Total Environ. 2019, 651, 2105–2117. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration. Process Paper, Satellite Products and Services Review Board, 2018, SPSRB Improvement Working Group, ver. 17, Department of Commerce, NOAA/NESDIS 23 July 2018. Available online: https://ecoforecast.org/wp-content/uploads/2019/11/SPSRB-Process-Paper_August-21-2018-_Final.pdf (accessed on 1 December 2022).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case study and Location | Method | Nowcast | Forecast Lead Time | Delivery Mode | Part of an Information System | Uptake |
|---|---|---|---|---|---|---|
| Fisheries | ||||||
| Tuna—eastern Australia | Habitat preferences | Yes | 0–3 months | No | Yes | |
| Tuna—southern Australia | Habitat preferences | Yes | 0–3 months | Website | No | Yes |
| Aquaculture | ||||||
| Hypoxia—Tasmania | Coastal model | Yes | <10 days | Dashboard | Yes, with interactive data explorer | Yes, but not sustained |
| SIMA-Austral Chile | Integrated models | Yes | Days to months | Website | Yes | Yes |
| Algal blooms | ||||||
| Yellow Sea, China | Ocean model | Yes | 6.5 days | Website | No | Yes, but not sustained |
| Tasmania | Environmental correlation | Yes | 0–3 months | Not yet | No | No |
| Iconic habitats and their users | ||||||
| Coral bleaching—GBR | eReefs models (hydrodynamic and biogeochemistry) | Yes | 3 days | Email and reports | Yes (nowcast) | Yes (through reports, papers) |
| Crown of Thorns—GBR | Combined hydrodynamic and ecosystem models | No | A few years | Reports and publications | No | No |
| Jellyfish—Queensland | GLM | Yes | <3 days | Publications | No | Only as guidelines |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Hobday, A.J.; Condie, S.A.; Baird, M.E.; Eveson, J.P.; Hartog, J.R.; Richardson, A.J.; Steven, A.D.L.; Wild-Allen, K.; Babcock, R.C.; et al. Ecological Forecasting and Operational Information Systems Support Sustainable Ocean Management. Forecasting 2022, 4, 1051-1079. https://doi.org/10.3390/forecast4040057
Sun C, Hobday AJ, Condie SA, Baird ME, Eveson JP, Hartog JR, Richardson AJ, Steven ADL, Wild-Allen K, Babcock RC, et al. Ecological Forecasting and Operational Information Systems Support Sustainable Ocean Management. Forecasting. 2022; 4(4):1051-1079. https://doi.org/10.3390/forecast4040057
Chicago/Turabian StyleSun, Chaojiao, Alistair J. Hobday, Scott A. Condie, Mark E. Baird, J. Paige Eveson, Jason R. Hartog, Anthony J. Richardson, Andrew D. L. Steven, Karen Wild-Allen, Russell C. Babcock, and et al. 2022. "Ecological Forecasting and Operational Information Systems Support Sustainable Ocean Management" Forecasting 4, no. 4: 1051-1079. https://doi.org/10.3390/forecast4040057