
Molecular and Functional Characterization of Caveolae in Mixed Cultures of Human NT-2 Neurons and Astrocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
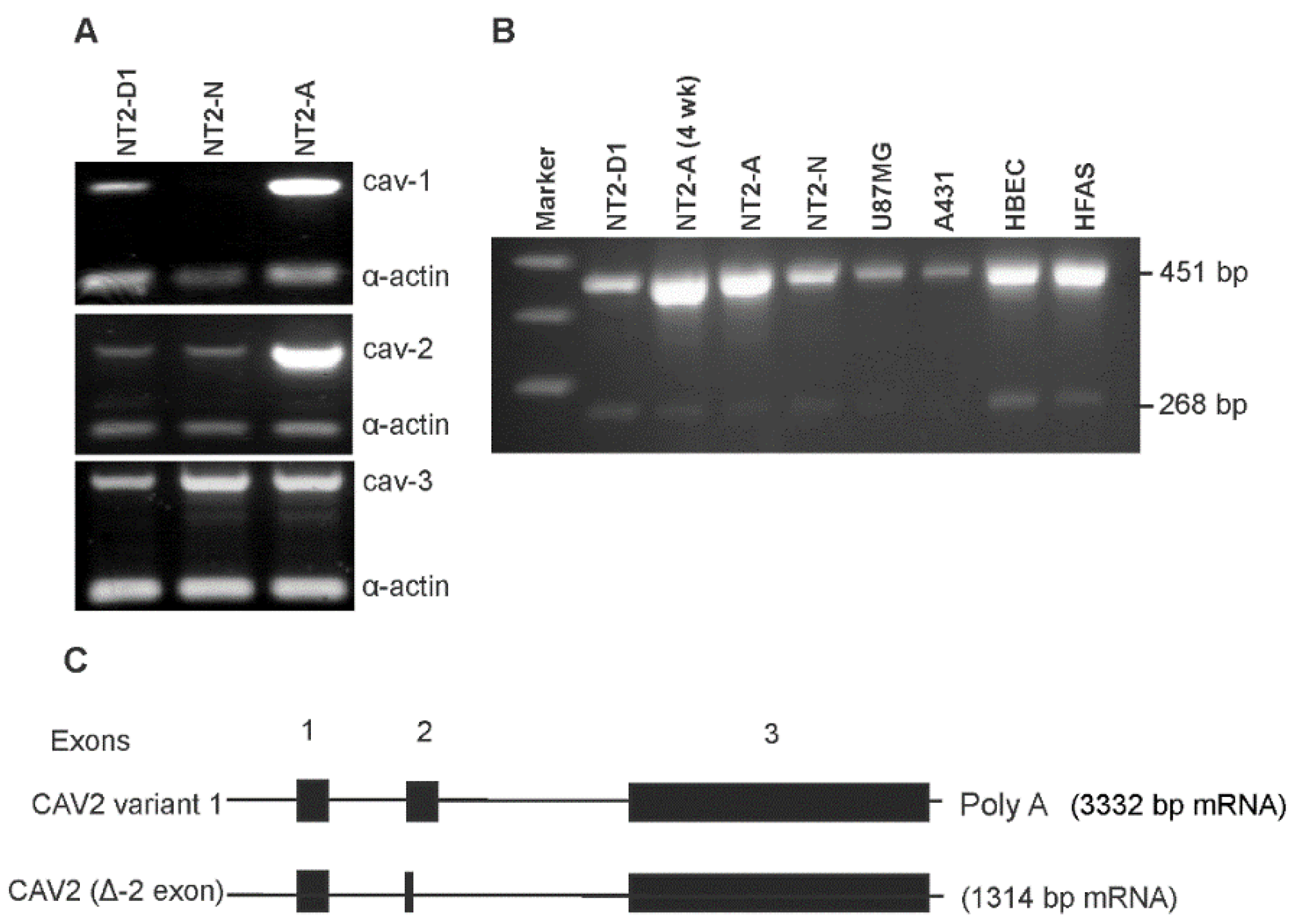
2.2. RNA Isolation and the Reverse Transcriptase-Polymerase Chain Reaction
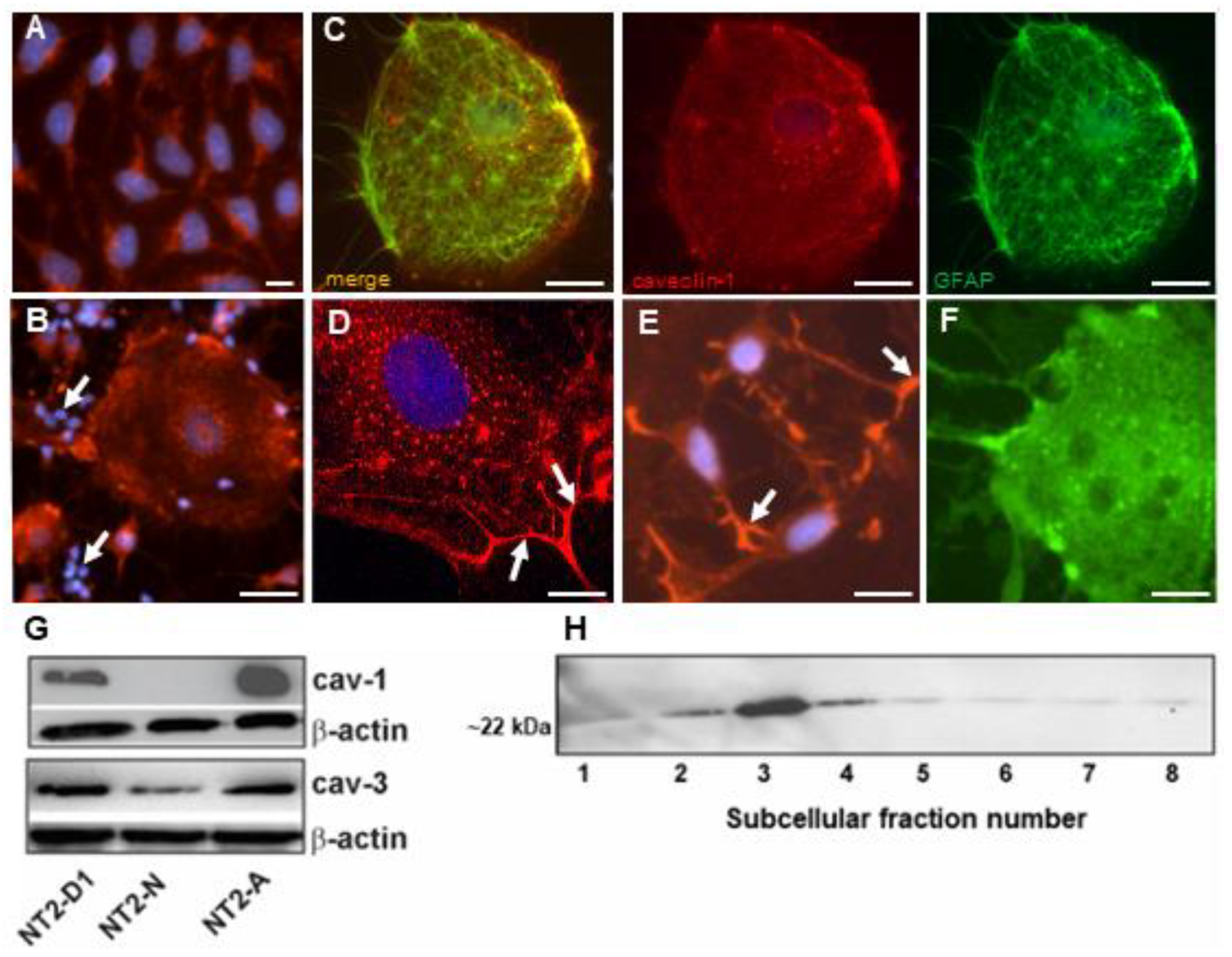
2.3. Subcellular Fractionation and Isolation of Low-Density Caveolae Membranes
2.4. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blot Analysis
2.5. Sequence Analysis
2.6. Immunofluorescence Microscopy
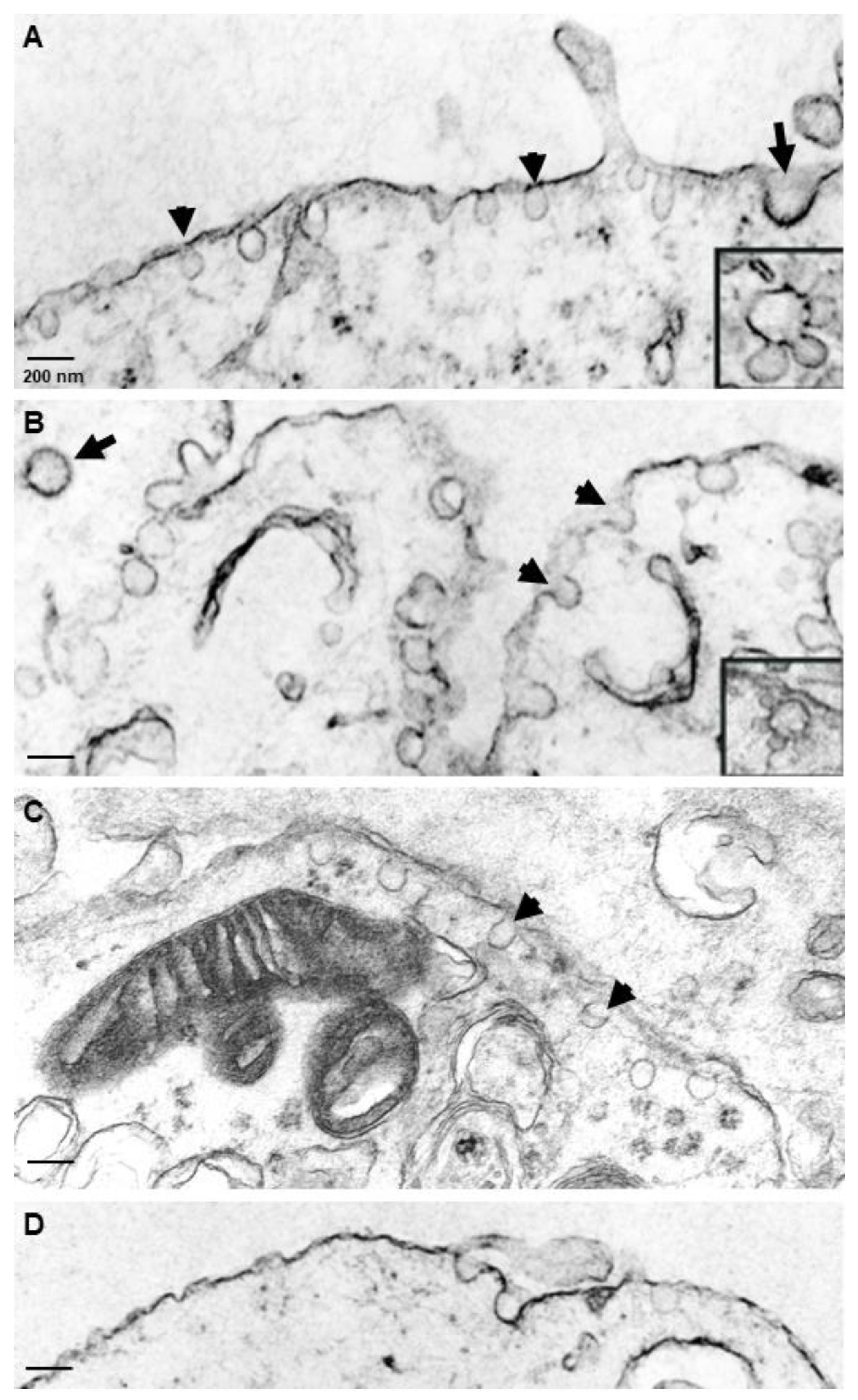
2.7. Transmission Electron Microscopy
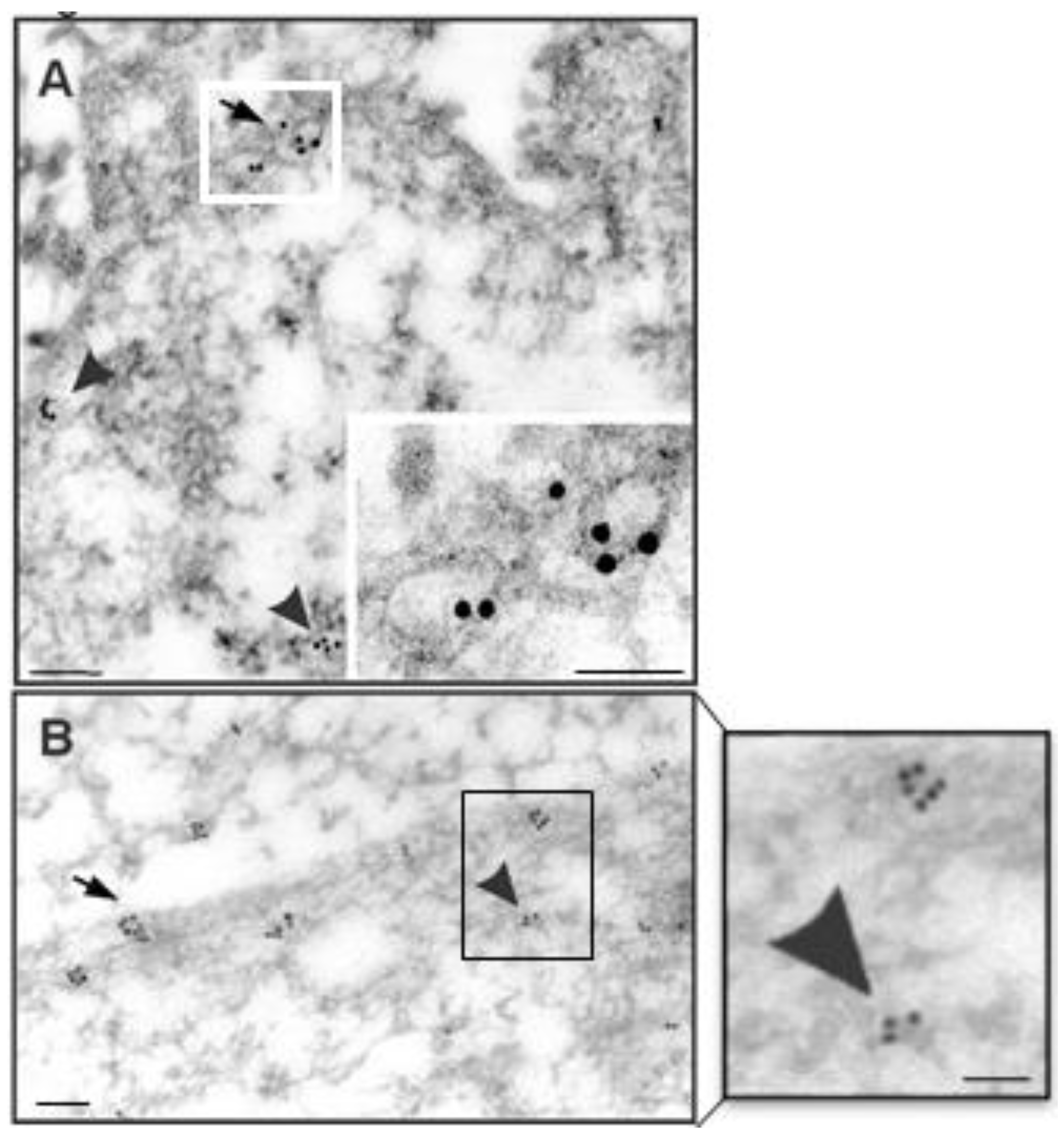
2.8. Immunoelectron Microscopy
2.9. Measurement of Endocytosis of Cholera Toxin (CTX-B)
2.10. Statistical Analysis
3. Results
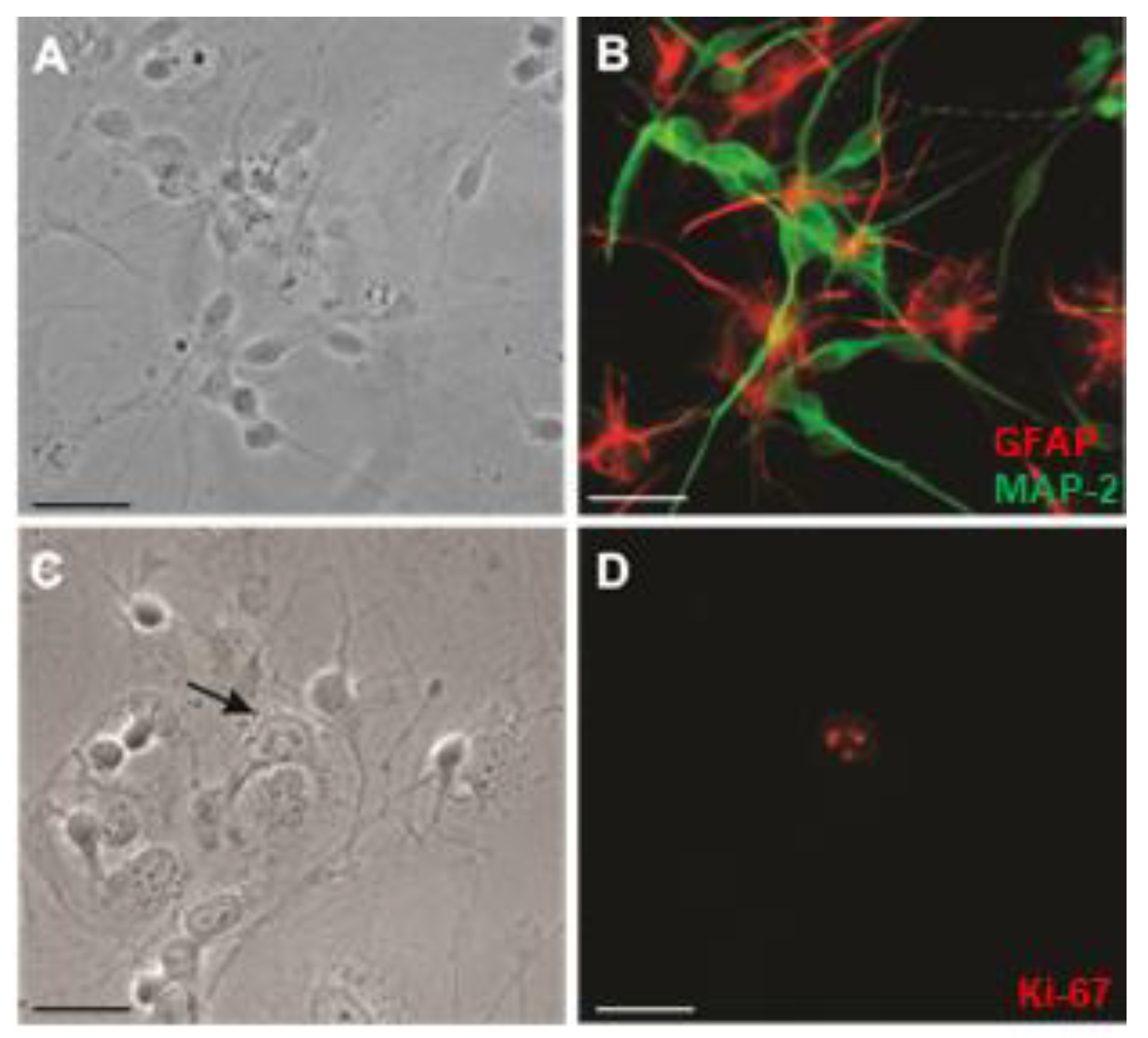
3.1. NT2 Precursor Cells Terminally Differentiate into Post-Mitotic NT2/N Neurons and NT2/A Astrocytes
3.2. Caveolin Proteins Are Differentially Expressed in NT2 Cells
3.3. Caveolae-like Structures Are Abundantly Expressed in Differentiated NT2/A and NT2/N Cells
3.4. Caveolin-1 Is Localized to Caveolae in NT2/A Astrocytes
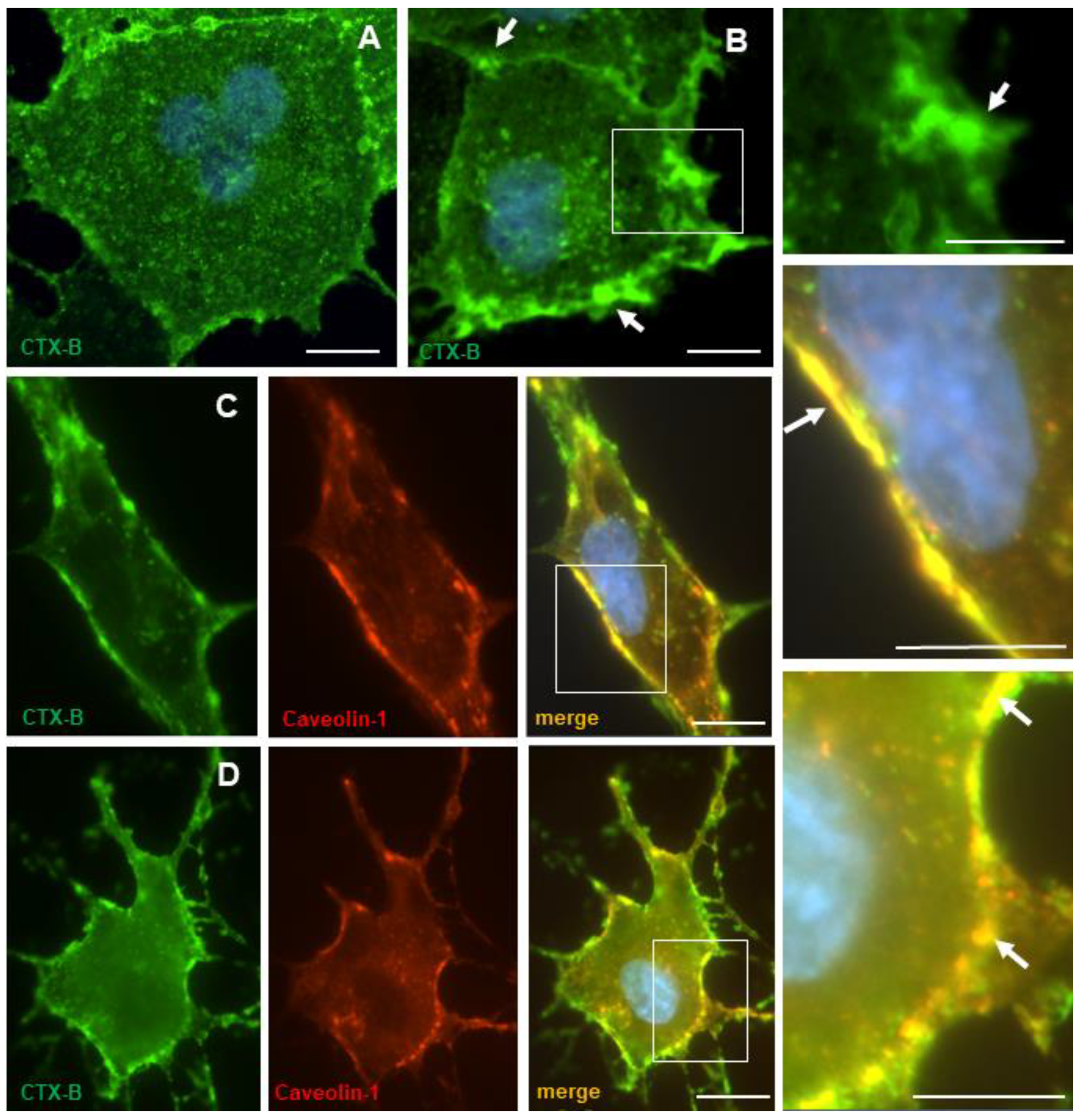
3.5. Cholera Toxin B-Subunit (CTX-B) Colocalizes with Caveolin-1 in Lipid Rafts
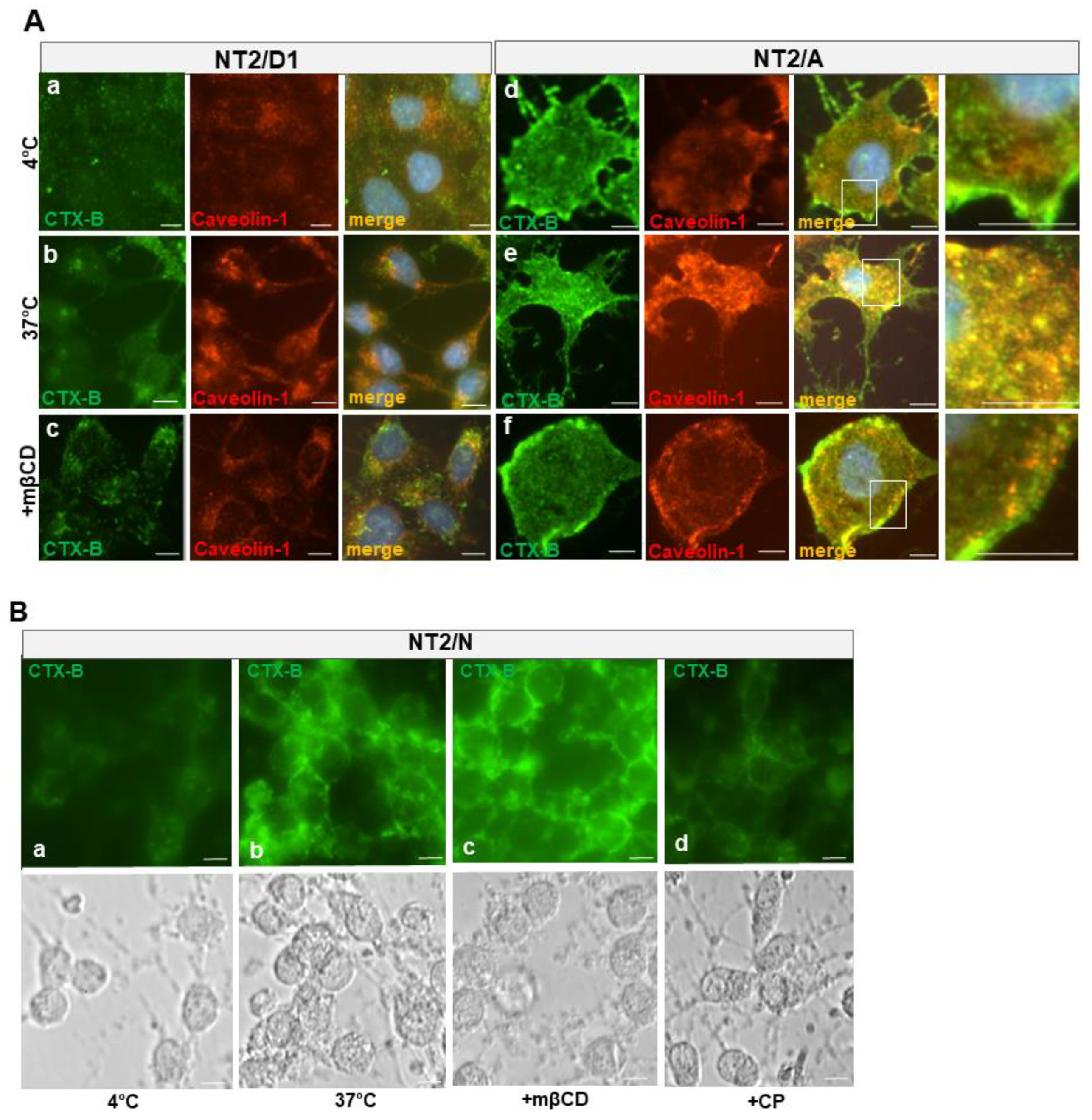
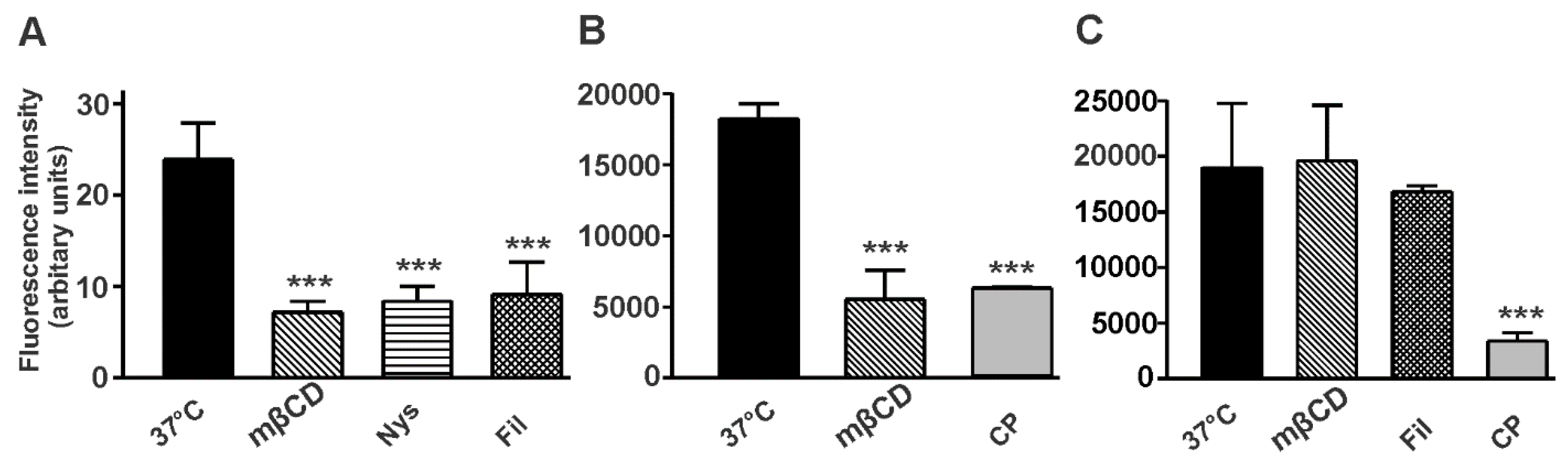
3.6. Cholera Toxin B-Subunit (CTX-B) Is Internalized by Different Mechanisms in NT2/A and NT2/N Cultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smart, E.J.; Graf, G.; McNiven, M.A.; Sessa, W.C.; Engelman, J.A.; Scherer, P.E.; Okamoto, T.; Lisanti, M.P. Caveolins, Liquid-Ordered Domains, and Signal Transduction. Mol. Cell. Biol. 1999, 19, 7289–7304. [Google Scholar] [CrossRef] [Green Version]
- Scherer, P.E.; Okamoto, T.; Chun, M.; Nishimoto, I.; Lodish, H.F.; Lisanti, M.P. Identification, sequence, and expression of caveolin-2 defines a caveolin gene family. Proc. Natl. Acad. Sci. USA 1996, 93, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Busija, A.R.; Patel, H.H.; Insel, P.A. Caveolins and cavins in the trafficking, maturation, and degradation of caveolae: Implications for cell physiology. Am. J. Physiol. Cell Physiol. 2017, 312, C459–C477. [Google Scholar] [CrossRef]
- Fra, A.M.; Williamson, E.; Simons, K.; Parton, R. De novo formation of caveolae in lymphocytes by expression of VIP21-caveolin. Proc. Natl. Acad. Sci. USA 1995, 92, 8655–8659. [Google Scholar] [CrossRef] [Green Version]
- Vogel, U.; Sandvig, K.; Van Deurs, B. Expression of caveolin-1 and polarized formation of invaginated caveolae in Caco-2 and MDCK II cells. J. Cell Sci. 1998, 111 Pt 6, 825–832. [Google Scholar] [CrossRef]
- Hailstones, D.; Sleer, L.S.; Parton, R.G.; Stanley, K.K. Regulation of caveolin and caveolae by cholesterol in MDCK cells. J. Lipid Res. 1998, 39, 369–379. [Google Scholar] [CrossRef]
- Couet, J.; Belanger, M.M.; Roussel, E.; Drolet, M.-C. Cell biology of caveolae and caveolin. Adv. Drug Deliv. Rev. 2001, 49, 223–235. [Google Scholar] [CrossRef]
- Razani, B.; Woodman, S.E.; Lisanti, M.P. Caveolae: From Cell Biology to Animal Physiology. Pharmacol. Rev. 2002, 54, 431–467. [Google Scholar] [CrossRef] [PubMed]
- Shaul, P.W.; Anderson, R.G.W. Role of plasmalemmal caveolae in signal transduction. Am. J. Physiol. 1998, 275, L843–L851. [Google Scholar] [CrossRef]
- Oka, N.; Yamamoto, M.; Schwencke, C.; Kawabe, J.I.; Ebina, T.; Ohno, S.; Couet, J.; Lisanti, M.P.; Ishikawa, Y. Caveolin interaction with protein kinase C. Isoenzyme-dependent regulation of kinase activity by the caveolin scaffolding domain peptide. J. Biol. Chem. 1997, 272, 33416–33421. [Google Scholar] [CrossRef] [Green Version]
- Parkin, E.; Hussain, I.; Turner, A.J.; Hooper, N. The Amyloid Precursor Protein Is Not Enriched in Caveolae-Like, Detergent-Insoluble Membrane Microdomains. J. Neurochem. 2002, 69, 2179–2188. [Google Scholar] [CrossRef]
- Ikezu, T.; Ueda, H.; Trapp, B.D.; Nishiyama, K.; Sha, J.F.; Volonte, D.; Galbiati, F.; Byrd, A.L.; Bassell, G.; Serizawa, H.; et al. Affinity-purification and characterization of caveolins from the brain: Differential expression of caveolin-1, -2, and -3 in brain endothelial and astroglial cell types. Brain Res. 1998, 804, 177–192. [Google Scholar] [CrossRef]
- Silva, W.; Maldonado, H.; Lisanti, M.; DeVellis, J.; Chompré, G.; Mayol, N.; Ortiz, M.; Velázquez, G.; Maldonado, A.; Montalvo, J. Identification of caveolae and caveolin in C6 glioma cells. Int. J. Dev. Neurosci. 1999, 17, 705–714. [Google Scholar] [CrossRef]
- Cameron, P.L.; Liu, C.; Smart, D.K.; Hantus, S.T.; Fick, J.R.; Cameron, R.S. Caveolin-1 expression is maintained in rat and human astroglioma cell lines. Glia 2002, 37, 275–290. [Google Scholar] [CrossRef]
- Niesman, I.R.; Zemke, N.; Fridolfsson, H.N.; Haushalter, K.J.; Levy, K.; Grove, A.; Schnoor, R.; Finley, J.C.; Patel, P.M.; Roth, D.M.; et al. Caveolin isoform switching as a molecular, structural, and metabolic regulator of microglia. Mol. Cell. Neurosci. 2013, 56, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, B.W.; Nuñez, V.; Kaplan, L.; Granger, A.J.; Bistrong, K.; Zucker, H.L.; Kumar, P.; Sabatini, B.L.; Gu, C. Caveolae in CNS arterioles mediate neurovascular coupling. Nat. Cell Biol. 2020, 579, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, F.; Volonté, D.; Gil, O.; Zanazzi, G.; Salzer, J.L.; Sargiacomo, M.; Scherer, P.E.; Engelman, J.A.; Schlegel, A.; Parenti, M.; et al. Expression of caveolin-1 and -2 in differentiating PC12 cells and dorsal root ganglion neurons: Caveolin-2 is up-regulated in response to cell injury. Proc. Natl. Acad. Sci. USA 1998, 95, 10257–10262. [Google Scholar] [CrossRef] [Green Version]
- Bu, J.; Bruckner, S.R.; Sengoku, T.; Geddes, J.W.; Estus, S. Glutamate regulates caveolin expression in rat hippocampal neurons. J. Neurosci. Res. 2003, 72, 185–190. [Google Scholar] [CrossRef]
- Cameron, P.L.; Ruffin, J.W.; Bollag, R.; Rasmussen, H.; Cameron, R.S. Identification of Caveolin and Caveolin-Related Proteins in the Brain. J. Neurosci. 1997, 17, 9520–9535. [Google Scholar] [CrossRef]
- Virgintino, D.; Robertson, D.; Errede, M.; Benagiano, V.; Tauer, U.; Roncali, L.; Bertossi, M. Expression of caveolin-1 in human brain microvessels. Neuroscience 2002, 115, 145–152. [Google Scholar] [CrossRef]
- Shikanai, M.; Nishimura, Y.V.; Sakurai, M.; Nabeshima, Y.-I.; Yuzaki, M.; Kawauchi, T. Caveolin-1 Promotes Early Neuronal Maturation via Caveolae-Independent Trafficking of N-Cadherin and L1. iScience 2018, 7, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.; Tuan, R.S. The less-often-traveled surface of stem cells: Caveolin-1 and caveolae in stem cells, tissue repair and regeneration. Stem Cell Res. Ther. 2013, 4, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Numakawa, T.; Shimazu, K.; Koshimizu, H.; Hara, T.; Hatanaka, H.; Mei, L.; Lu, B.; Kojima, M. BDNF-induced recruitment of TrkB receptor into neuronal lipid rafts: Roles in synaptic modulation. J. Cell Biol. 2004, 167, 1205–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.-J.; Chung, Y.H.; Hwang, C.-I.; Murata, M.; Fujimoto, T.; Mook-Jung, I.-H.; Cha, C.I.; Park, W.-Y. Caveolin-1 upregulation in senescent neurons alters amyloid precursor protein processing. Exp. Mol. Med. 2006, 38, 126–133. [Google Scholar] [CrossRef]
- Head, B.P.; Peart, J.N.; Panneerselvam, M.; Yokoyama, T.; Pearn, M.L.; Niesman, I.R.; Bonds, J.A.; Schilling, J.M.; Miyanohara, A.; Headrick, J.; et al. Loss of Caveolin-1 Accelerates Neurodegeneration and Aging. PLoS ONE 2010, 5, e15697. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, C.; Huang, Q.; Wu, D.; Cao, J.; Xu, X.; Yang, C.; Li, X. Caveolin-1 Plays an Important Role in the Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells into Cardiomyocytes. Cardiology 2016, 136, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, C.; Wang, Y.-J.; Guan, X.; Wang, L.; Shen, L.-M.; Zou, W. Caveolin-1 downregulation promotes the dopaminergic neuron-like differentiation of human adipose-derived mesenchymal stem cells. Neural Regen. Res. 2021, 16, 714–720. [Google Scholar] [CrossRef]
- Wang, S.; Kan, Q.; Sun, Y.; Han, R.; Zhang, G.; Peng, T.; Jia, Y. Caveolin-1 regulates neural differentiation of rat bone mesenchymal stem cells into neurons by modulating Notch signaling. Int. J. Dev. Neurosci. 2012, 31, 30–35. [Google Scholar] [CrossRef]
- Li, Y.; Luo, J.; Lau, W.M.; Zheng, G.-Q.; Fu, S.; Wang, T.-T.; Zeng, H.; So, K.-F.; Chung, S.K.; Tong, Y.; et al. Caveolin-1 Plays a Crucial Role in Inhibiting Neuronal Differentiation of Neural Stem/Progenitor Cells via VEGF Signaling-Dependent Pathway. PLoS ONE 2011, 6, e22901. [Google Scholar] [CrossRef] [Green Version]
- Pleasure, S.J.; Lee, V.M.-Y. NTera 2 Cells: A human cell line which displays characteristics expected of a human committed neuronal progenitor cell. J. Neurosci. Res. 1993, 35, 585–602. [Google Scholar] [CrossRef]
- Sandhu, J.K.; Sikorska, M.; Walker, P.R. Characterization of Astrocytes Derived From Human NTera-2/D1 Embryonal Carcinoma Cells. J. Neurosci. Res. 2002, 68, 604–614. [Google Scholar] [CrossRef]
- Tarczyluk, M.A.; Nagel, D.A.; O’Neil, J.D.; Parri, R.; Tse, E.; Coleman, M.D.; Hill, E. Functional Astrocyte-Neuron Lactate Shuttle in a Human Stem Cell-Derived Neuronal Network. J. Cereb. Blood Flow Metab. 2013, 33, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.S.; Sikorska, M.; Walker, P.R.; Sandhu, J.K. Effects of glutathione depletion on the viability of human NT2-derived neuronal and astroglial cultures. Neuron Glia Biol. 2004, 1, 317–326. [Google Scholar] [CrossRef]
- Smart, E.J.; Ying, Y.S.; Mineo, C.; Anderson, R.G. A detergent-free method for purifying caveolae membrane from tissue culture cells. Proc. Natl. Acad. Sci. USA 1995, 92, 10104–10108. [Google Scholar] [CrossRef] [Green Version]
- Janes, P.; Ley, S.C.; Magee, A.I. Aggregation of Lipid Rafts Accompanies Signaling via the T Cell Antigen Receptor. J. Cell Biol. 1999, 147, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, J.K.; Pandey, S.; Ribecco-Lutkiewicz, M.; Monette, R.; Borowy-Borowski, H.; Walker, P.R.; Sikorska, M. Molecular mechanisms of glutamate neurotoxicity in mixed cultures of NT2-derived neurons and astrocytes: Protective effects of coenzyme Q10. J. Neurosci. Res. 2003, 72, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Williams, T.M.; Lisanti, M.P. The caveolin proteins. Genome Biol. 2004, 5, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parton, R.G. Ultrastructural localization of gangliosides; GM1 is concentrated in caveolae. J. Histochem. Cytochem. 1994, 42, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Nichols, B. Caveosomes and endocytosis of lipid rafts. J. Cell Sci. 2003, 116, 4707–4714. [Google Scholar] [CrossRef] [Green Version]
- Parton, R.G.; Richards, A.A. Lipid Rafts and Caveolae as Portals for Endocytosis: New Insights and Common Mechanisms. Traffic 2003, 4, 724–738. [Google Scholar] [CrossRef] [Green Version]
- Torgersen, M.L.; Skretting, G.; Van Deurs, B.; Sandvig, K. Internalization of cholera toxin by different endocytic mechanisms. J. Cell Sci. 2001, 114, 3737–3747. [Google Scholar] [CrossRef]
- Okamoto, T.; Schlegel, A.; Scherer, P.E.; Lisanti, M.P. Caveolins, a Family of Scaffolding Proteins for Organizing “Preassembled Signaling Complexes” at the Plasma Membrane. J. Biol. Chem. 1998, 273, 5419–5422. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, M.G.; Werner, S. Caveolin-1 and -2 Expression Is Differentially Regulated in Cultured Keratinocytes and within the Regenerating Epidermis of Cutaneous Wounds. Exp. Cell Res. 2000, 258, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.Y.; Carpentier, J.L.; Van Obberghen, E.; Grunfeld, C.; Gorden, P.; Orci, L. Morphological changes of the 3T3-L1 fibroblast plasma membrane upon differentiation to the adipocyte form. J. Cell Sci. 1983, 61, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, F.; Volonte, D.; Liu, J.; Capozza, F.; Frank, P.G.; Zhu, L.; Pestell, R.G.; Lisanti, M.P. Caveolin-1 expression negatively regulates cell cycle progression by inducing G(0)/G(1) arrest via a p53/p21(WAF1/Cip1)-dependent mechanism. Mol. Biol. Cell 2001, 12, 2229–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abulrob, A.; Giuseppin, S.; Andrade, M.F.; McDermid, A.; Moreno, M.; Stanimirovic, D. Interactions of EGFR and caveolin-1 in human glioblastoma cells: Evidence that tyrosine phosphorylation regulates EGFR association with caveolae. Oncogene 2004, 23, 6967–6979. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lau, W.M.; So, K.F.; Tong, Y.; Shen, J. Caveolin-1 promote astroglial differentiation of neural stem/progenitor cells through modulating Notch1/NICD and Hes1 expressions. Biochem. Biophys. Res. Commun. 2011, 407, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Kogo, H.; Ishiguro, K.; Kuwaki, S.; Fujimoto, T. Identification of a splice variant of mouse caveolin-2 mRNA encoding an isoform lacking the C-terminal domain. Arch. Biochem. Biophys. 2002, 401, 108–114. [Google Scholar] [CrossRef]
- Gorodinsky, A.; Harris, D. Glycolipid-anchored proteins in neuroblastoma cells form detergent-resistant complexes without caveolin. J. Cell Biol. 1995, 129, 619–627. [Google Scholar] [CrossRef]
- Li, W.P.; Liu, P.; Pilcher, B.K.; Anderson, R.G. Cell-specific targeting of caveolin-1 to caveolae, secretory vesicles, cytoplasm or mitochondria. J. Cell Sci. 2001, 114, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Gudas, L.J.; Wagner, J.A. Retinoids regulate stem cell differentiation. J. Cell. Physiol. 2010, 226, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Song, K.S.; Koh, S.S.; Kikuchi, A.; Lisanti, M.P. Baculovirus-based expression of mammalian caveolin in Sf21 insect cells. A model system for the biochemical and morphological study of caveolae biogenesis. J. Biol. Chem. 1996, 271, 28647–28654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.S.; Scherer, P.E.; Tang, Z.; Okamoto, T.; Li, S.; Chafel, M.; Chu, C.; Kohtz, D.S.; Lisanti, M.P. Expression of caveolin-3 in skeletal, cardiac, and smooth muscle cells. Caveolin-3 is a component of the sarcolemma and co-fractionates with dystrophin and dystrophin-associated glycoproteins. J. Biol. Chem. 1996, 271, 15160–15165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, Z.; Almenar-Queralt, A.; Leem, J.; DerMardirossian, C.; Roth, D.M.; Patel, P.M.; Patel, H.H.; Head, B.P. Caveolin-1 Phosphorylation Is Essential for Axonal Growth of Human Neurons Derived From iPSCs. Front. Cell. Neurosci. 2019, 13, 324. [Google Scholar] [CrossRef] [Green Version]
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Kavaliauskiene, S.; Skotland, T. Clathrin-independent endocytosis: An increasing degree of complexity. Histochem. Cell Biol. 2018, 150, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shogomori, H.; Futerman, A. Cholera Toxin Is Found in Detergent-insoluble Rafts/Domains at the Cell Surface of Hippocampal Neurons but Is Internalized via a Raft-independent Mechanism. J. Biol. Chem. 2001, 276, 9182–9188. [Google Scholar] [CrossRef] [Green Version]
- Nichols, B.J. A distinct class of endosome mediates clathrin-independent endocytosis to the Golgi complex. Nat. Cell Biol. 2002, 4, 374–378. [Google Scholar] [CrossRef]
- Pang, H.; Le, P.U.; Nabi, I.R. Ganglioside GM1 levels are a determinant of the extent of caveolae/raft-dependent endocytosis of cholera toxin to the Golgi apparatus. J. Cell Sci. 2004, 117, 1421–1430. [Google Scholar] [CrossRef] [Green Version]
- Minshall, R.D.; Tiruppathi, C.; Vogel, S.M.; Niles, W.D.; Gilchrist, A.; Hamm, H.E.; Malik, A.B. Endothelial cell-surface gp60 activates vesicle formation and trafficking via G(i)-coupled Src kinase signaling pathway. J. Cell Biol. 2000, 150, 1057–1070. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.K.; Brown, J.C.; Choudhury, A.; Peterson, T.E.; Holicky, E.; Marks, D.L.; Simari, R.; Parton, R.; Pagano, R.E. Selective Stimulation of Caveolar Endocytosis by Glycosphingolipids and Cholesterol. Mol. Biol. Cell 2004, 15, 3114–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blochet, C.; Buscemi, L.; Clément, T.; Gehri, S.; Badaut, J.; Hirt, L. Involvement of caveolin-1 in neurovascular unit remodeling after stroke: Effects on neovascularization and astrogliosis. Br. J. Pharmacol. 2018, 40, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, L.; Wen, Z.; Wu, L.; Jiang, Y.; Yang, L.; Xiao, L.; Xie, Y.; Ma, M.; Zhu, W.; et al. Caveolin-1 is a checkpoint regulator in hypoxia-induced astrocyte apoptosis via Ras/Raf/ERK pathway. Am. J. Physiol. Physiol. 2016, 310, C903–C910. [Google Scholar] [CrossRef] [Green Version]
- Bi, C.; Tham, D.K.L.; Perronnet, C.; Joshi, B.; Nabi, I.R.; Moukhles, H. The Oxidative Stress-Induced Increase in the Membrane Expression of the Water-Permeable Channel Aquaporin-4 in Astrocytes Is Regulated by Caveolin-1 Phosphorylation. Front. Cell. Neurosci. 2017, 11, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.H.; Park, S.J.; Jo, A.; Kang, J.L.; Jou, I.; Park, J.S.; Choi, Y.-H. Caveolin-1 is involved in reactive oxygen species-induced SHP-2 activation in astrocytes. Exp. Mol. Med. 2011, 43, 660–668. [Google Scholar] [CrossRef]
- Filchenko, I.; Blochet, C.; Buscemi, L.; Price, M.; Badaut, J.; Hirt, L. Caveolin-1 Regulates Perivascular Aquaporin-4 Expression after Cerebral Ischemia. Front. Cell Dev. Biol. 2020, 8, 371. [Google Scholar] [CrossRef]
- Bacci, A.; Verderio, C.; Pravettoni, E.; Matteoli, M. The role of glial cells in synaptic function. Philos. Trans. R. Soc. Lond B Biol. Sci. 1999, 354, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Svendsen, C.N. The amazing astrocyte. Nat. Cell Biol. 2002, 417, 29–32. [Google Scholar] [CrossRef]
- Yang, W.; Geng, C.; Yang, Z.; Xu, B.; Shi, W.; Yang, Y.; Tian, Y. Deciphering the roles of caveolin in neurodegenerative diseases: The good, the bad and the importance of context. Ageing Res. Rev. 2020, 62, 101116. [Google Scholar] [CrossRef] [PubMed]
- Bouillot, C.; Prochiantz, A.; Rougon, G.; Allinquant, B. Axonal Amyloid Precursor Protein Expressed by Neurons in Vitro Is Present in a Membrane Fraction with Caveolae-like Properties. J. Biol. Chem. 1996, 271, 7640–7644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikezu, T.; Trapp, B.D.; Song, K.S.; Schlegel, A.; Lisanti, M.P.; Okamoto, T. Caveolae, Plasma Membrane Microdomains for α-Secretase-mediated Processing of the Amyloid Precursor Protein. J. Biol. Chem. 1998, 273, 10485–10495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudreault, S.B.; Dea, D.; Poirier, J. Increased caveolin-1 expression in Alzheimer’s disease brain. Neurobiol. Aging 2004, 25, 753–759. [Google Scholar] [CrossRef]
- Nishiyama, K.; Trapp, B.D.; Ikezu, T.; Ransohoff, R.M.; Tomita, T.; Iwatsubo, T.; Kanazawa, I.; Hsiao, K.K.; Lisanti, M.P.; Okamoto, T. Caveolin-3 upregulation activates beta-secretase-mediated cleavage of the amyloid precursor protein in Alzheimer’s disease. J. Neurosci. 1999, 19, 6538–6548. [Google Scholar] [CrossRef]
- Trushina, E.; Canaria, C.A.; Lee, D.Y.; McMurray, C.T. Loss of caveolin-1 expression in knock-in mouse model of Huntington’s disease suppresses pathophysiology in vivo. Hum. Mol. Genet. 2014, 23, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Sawada, A.; Wang, S.; Jian, M.; Leem, J.; Wackerbarth, J.; Egawa, J.; Schilling, J.M.; Platoshyn, O.; Zemljic-Harpf, A.; Roth, D.M.; et al. Neuron-targeted caveolin-1 improves neuromuscular function and extends survival in SOD1 G93A mice. FASEB J. 2019, 33, 7545–7554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.H.; Choi, Y.R.; Heo, C.H.; Kang, S.J.; Joe, E.H.; Jou, I.; Kim, H.M.; Park, S.M. Loss of parkin promotes lipid rafts-dependent endocytosis through accumulating caveolin-1: Implications for Parkinson’s disease. Mol. Neurodegener. 2015, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-M.; Cha, S.-H.; Choi, Y.R.; Jou, I.; Joe, E.-H.; Park, S.M. DJ-1 deficiency impairs glutamate uptake into astrocytes via the regulation of flotillin-1 and caveolin-1 expression. Sci. Rep. 2016, 6, 28823. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size (bp) | GenBank Accession Number |
|---|---|---|---|
| Caveolin-1 | F: 5′ TCA ACC GCG ACC CTA AAC ACC 3′ R: 5′ TGA AAT AGC TCA GAA GAG ACA T 3′ | 561 | Z18951 |
| Caveolin-2 | F: 5′ AAGCTTTTCATGGACGACGACTCCTAC 3′ R: 5′ ACAATCCTGGCTCAGTTGCA 3′ | 451 | U32114 |
| Caveolin-3 | F: 5′ GAA GGA GGT CTA AAG CCA GG 3′ R: 5′ ATC TTA CAG GCA CGA ACA AA 3′ | 458 | NM_033337 |
| β-actin | F: 5′ GGA GCA ATG ATC TTG ATC TT 3′ R: 5′ CCT TCC TGG GCA TGG AGT CCT 3′ | 250 | NM_001613 |
| Cell Type | Size (nm) | Number of Caveolae-like Structures Measured |
|---|---|---|
| NT2/A (4 wk) | 97.6 ± 16.7 | 40 |
| NT2/A | 94.8 ± 11.5 | 34 |
| NT2/N | 102 ± 7.7 | 61 |
| NT2/D1 | 86 ± 20.8 | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandhu, J.K.; Ribecco-Lutkiewicz, M.; Abulrob, A. Molecular and Functional Characterization of Caveolae in Mixed Cultures of Human NT-2 Neurons and Astrocytes. Neuroglia 2021, 2, 68-88. https://doi.org/10.3390/neuroglia2010008
Sandhu JK, Ribecco-Lutkiewicz M, Abulrob A. Molecular and Functional Characterization of Caveolae in Mixed Cultures of Human NT-2 Neurons and Astrocytes. Neuroglia. 2021; 2(1):68-88. https://doi.org/10.3390/neuroglia2010008
Chicago/Turabian StyleSandhu, Jagdeep K., Maria Ribecco-Lutkiewicz, and Abedelnasser Abulrob. 2021. "Molecular and Functional Characterization of Caveolae in Mixed Cultures of Human NT-2 Neurons and Astrocytes" Neuroglia 2, no. 1: 68-88. https://doi.org/10.3390/neuroglia2010008