
Successional Pathways of Avifauna in a Shifting Mosaic Landscape: Interplay between Land Abandonment and Wildfires

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Sampling Design
- (1)
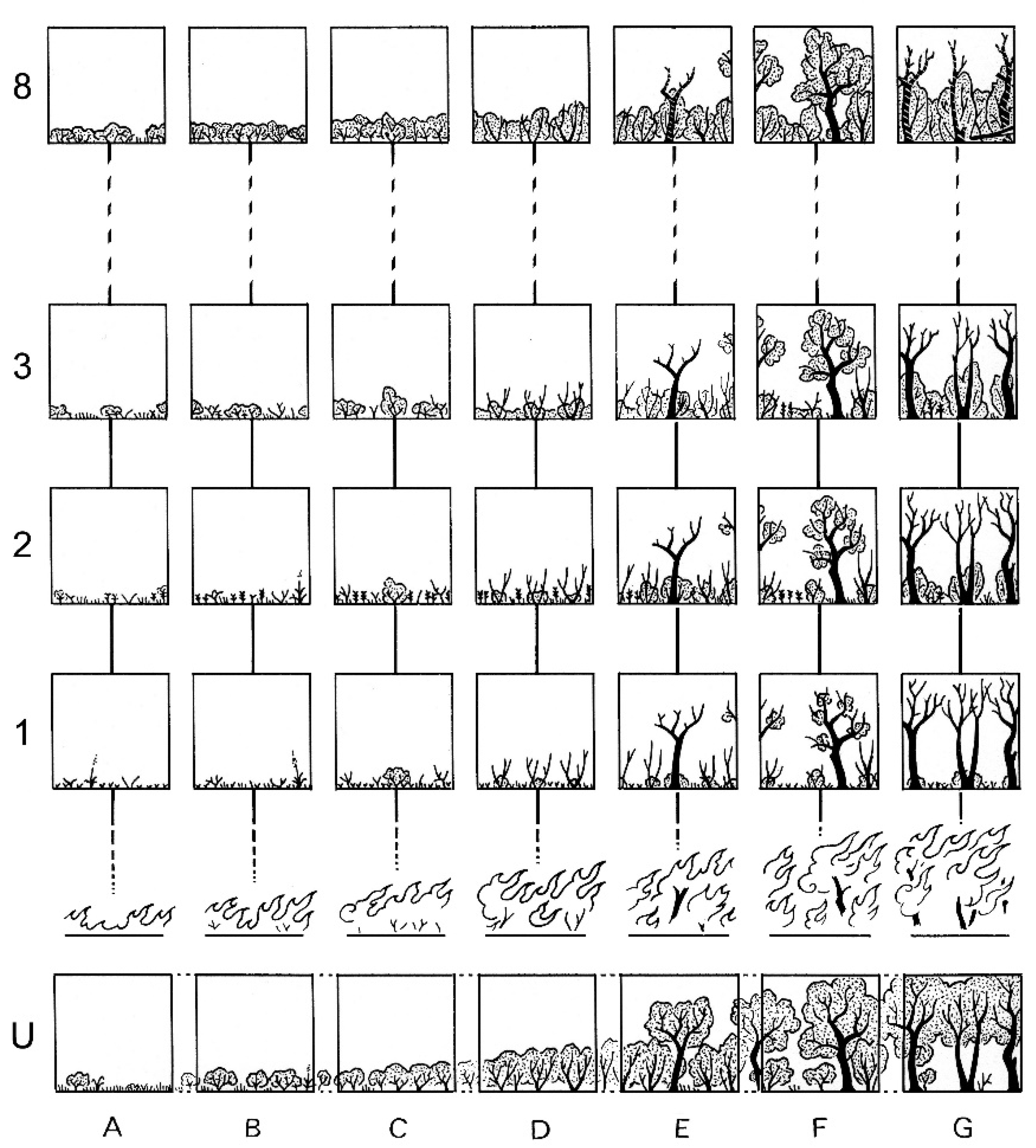
- In a first step, we conducted stratified sampling of a sequence of vegetation types. In the unburned landscape mosaic of the study area, we empirically defined seven a priori classes on a grassland-to-forest gradient of increasing vegetation height and cover (i.e., increasing maturity):
- Dry or rocky grasslands, whose dominant species is Brachypodium retusum, with a few scattered low shrubs (we could not find any grassland completely free of scrub) (n = 9 sampling stations);
- Low and patchy scrub (generally Ulex parviflorus and Cistus spp.) with grass (n = 19);
- Low maquis (Ulex parviflorus, Cistus spp. and Calicotome spinosa dominating), about 1–1.5 m high (n = 6);
- Closed heather maquis (Erica arborea and E. scoparia) with Phillyrea sp., about 2 m high (n = 13);
- Heather maquis with scattered oaks (Quercus ilex or Q. suber) (n = 7);
- Clear cork oak (Quercus suber) forest with a more or less dense understorey (n = 47);
- Closed holm oak (Quercus ilex) forest with a clear understorey (n = 15).
- (2)
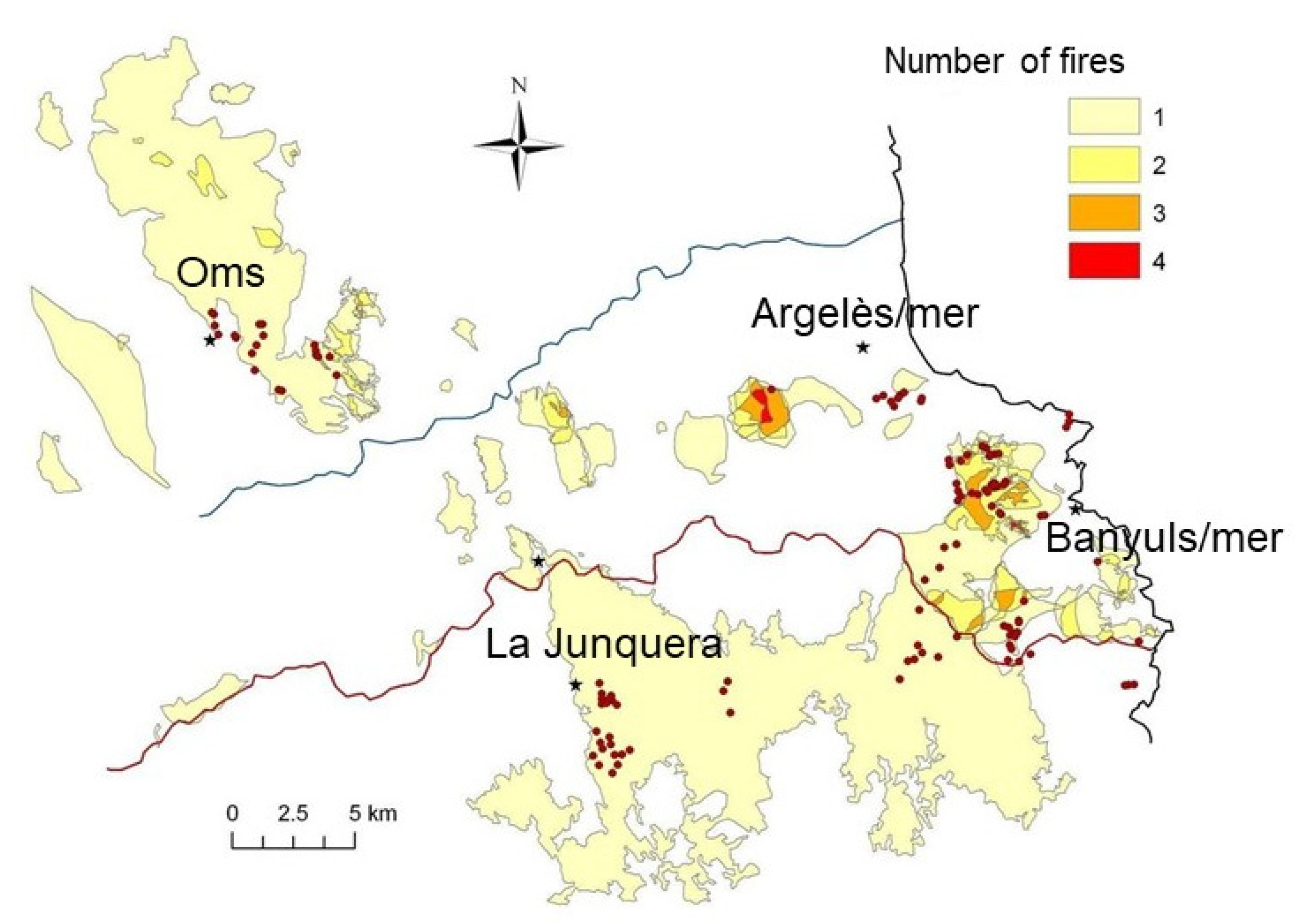
- In a second step, in a before–after design, we then conducted postfire diachronic sampling on 71 selected stations belonging to the seven structural classes of the above gradient (Figure 2). This phase of the field sampling was constrained by the unpredictable occurrence and location of wildfires, by the availability of a particular vegetation type of the above categories within their perimeter, and by the presence of formerly sampled stations. Station monitoring was carried out annually, at least for the first eight years after the fire whenever possible (repeated fires could interrupt the sequences). In class G (holm oak forests; Figure 1), where no prefire samples were available, control sampling stations were selected in proximity. Due to the staggered fire dates, monitoring was not synchronous in all these stations. The large difference in surface area of the respective plant formations in the landscape, the alteration of certain stations during the study that led to the interruption of their monitoring, and various other constraints did not allow for balanced class–year numbers. The staggered fire dates smoothed out year effects other than time-since-fire in the comparisons of postfire trajectories, which compensated for the lack of parallel controls that in most cases would have been impossible to find in this rugged and heterogeneous landscape.
- (3)
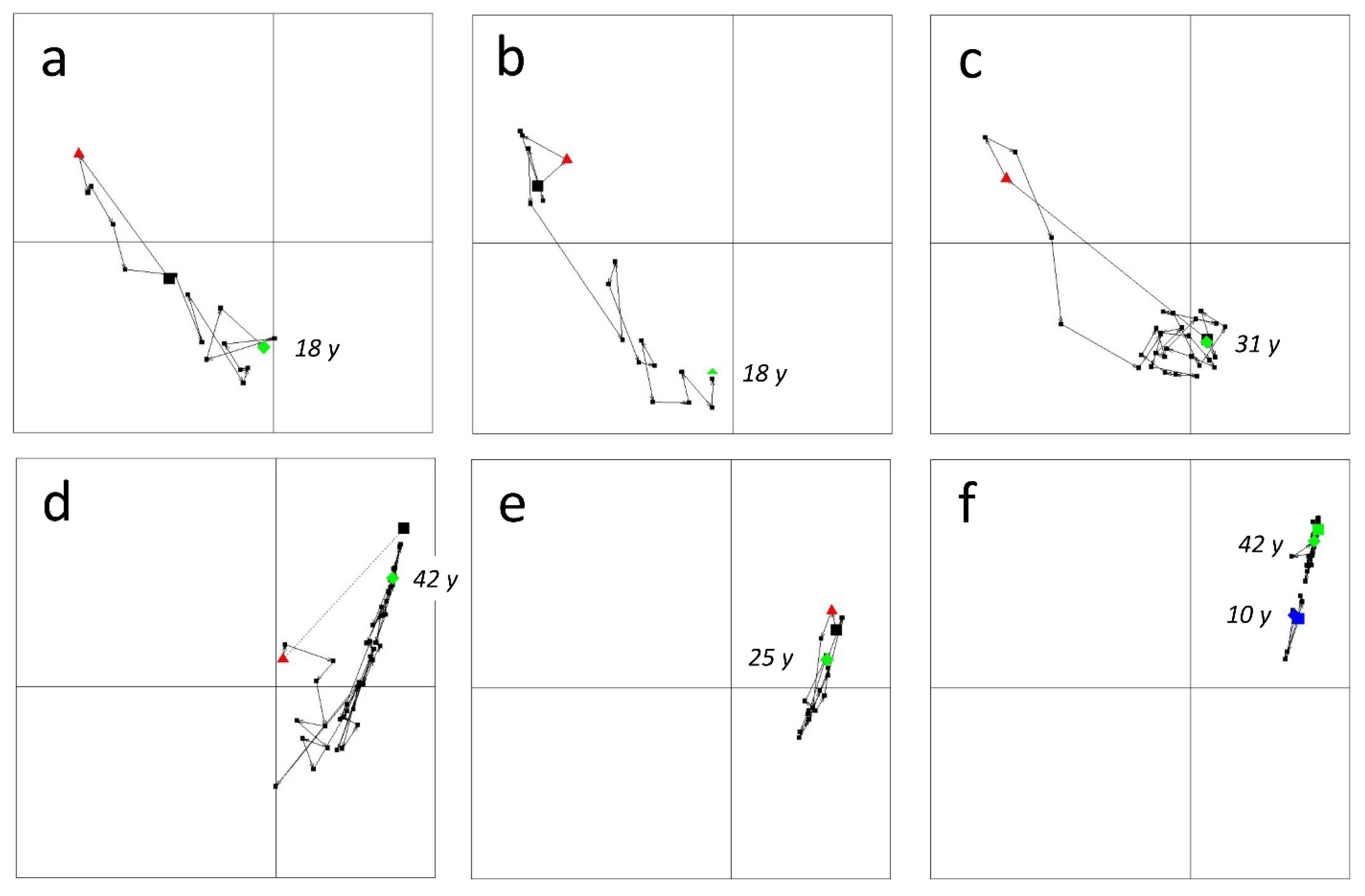
- Depending on the opportunities offered by the occurrence of large wildfires during the study period, some stations—not sampled before the fire—were monitored diachronically, up to 42 years in some cases (Figure A3). These were compared as far as possible with control stations selected in the vicinity.
2.3. Sampling Method
2.4. Statistical Analysis
3. Results
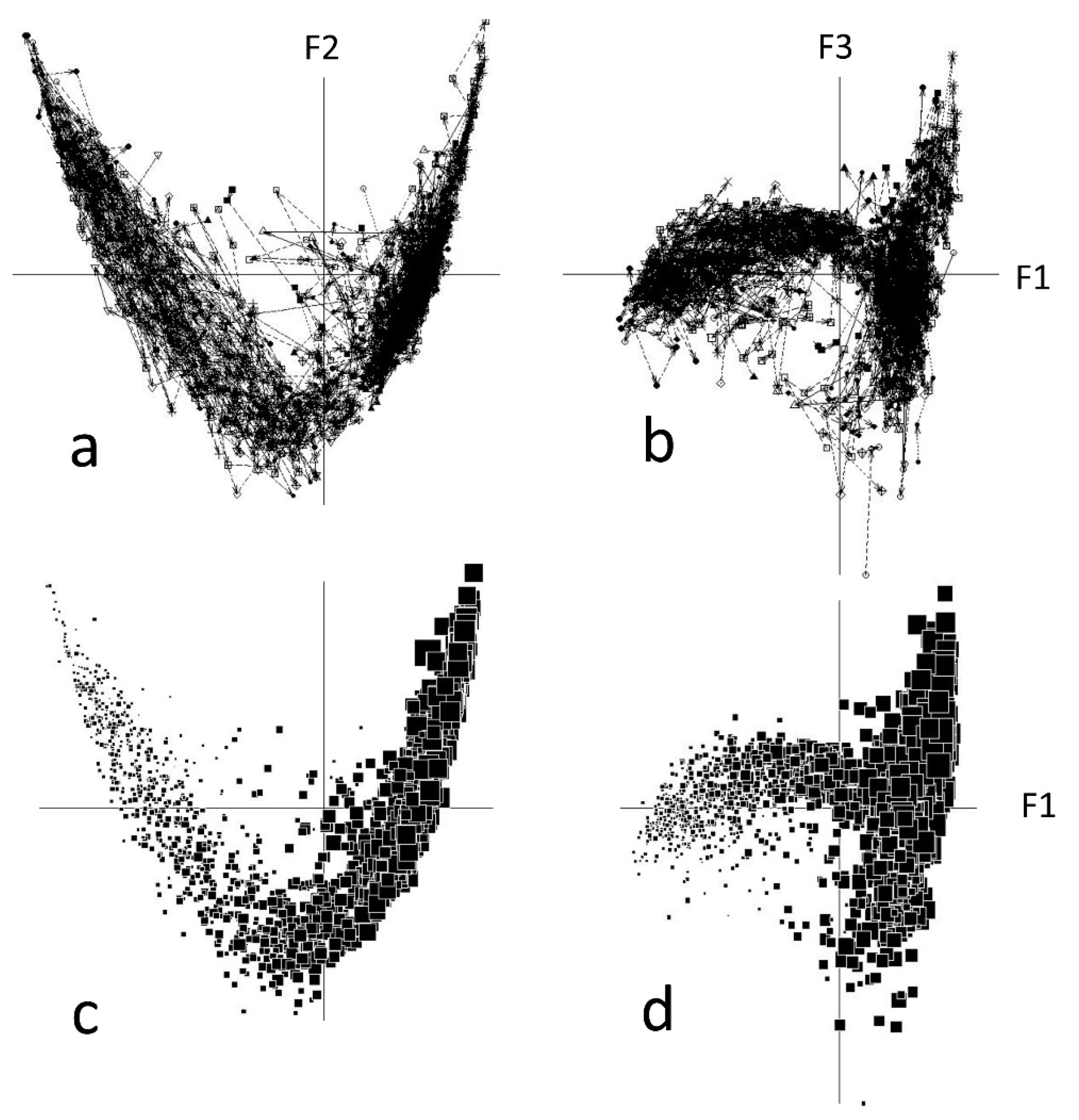
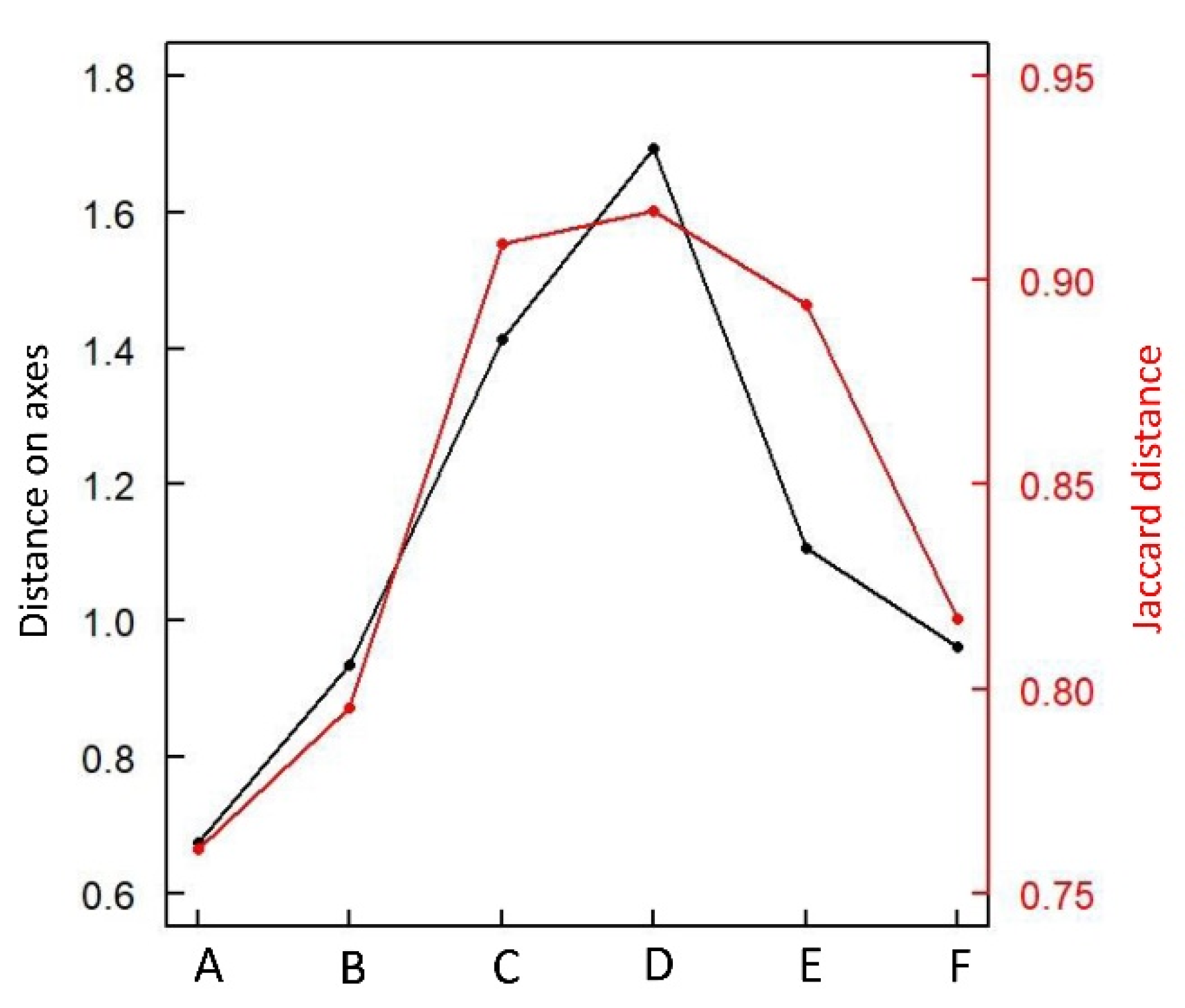
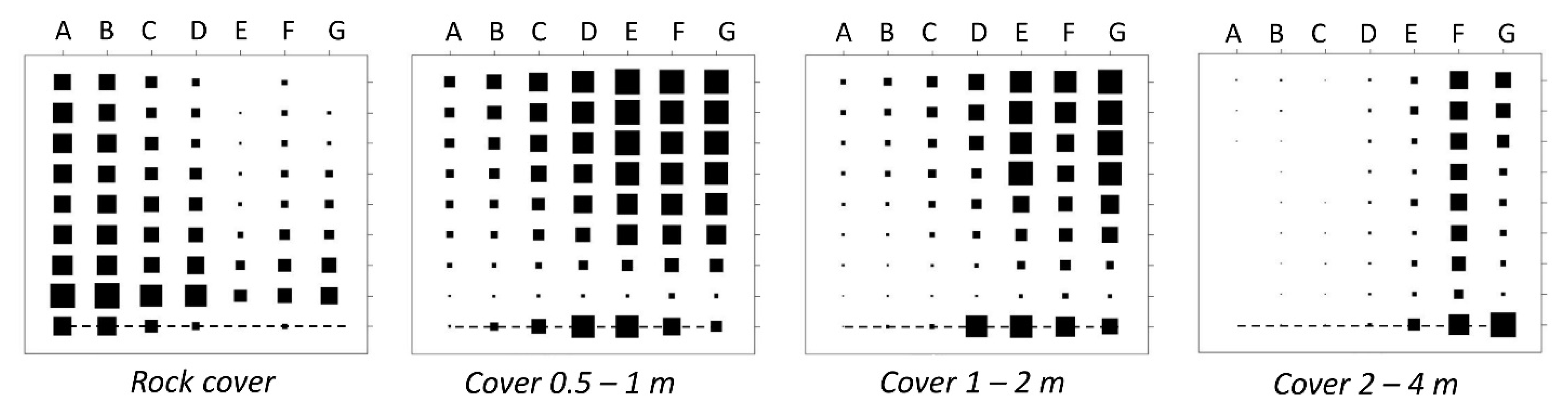
3.1. Species Distribution in Two Dimensions: Vegetation Gradient vs. Postfire Succession
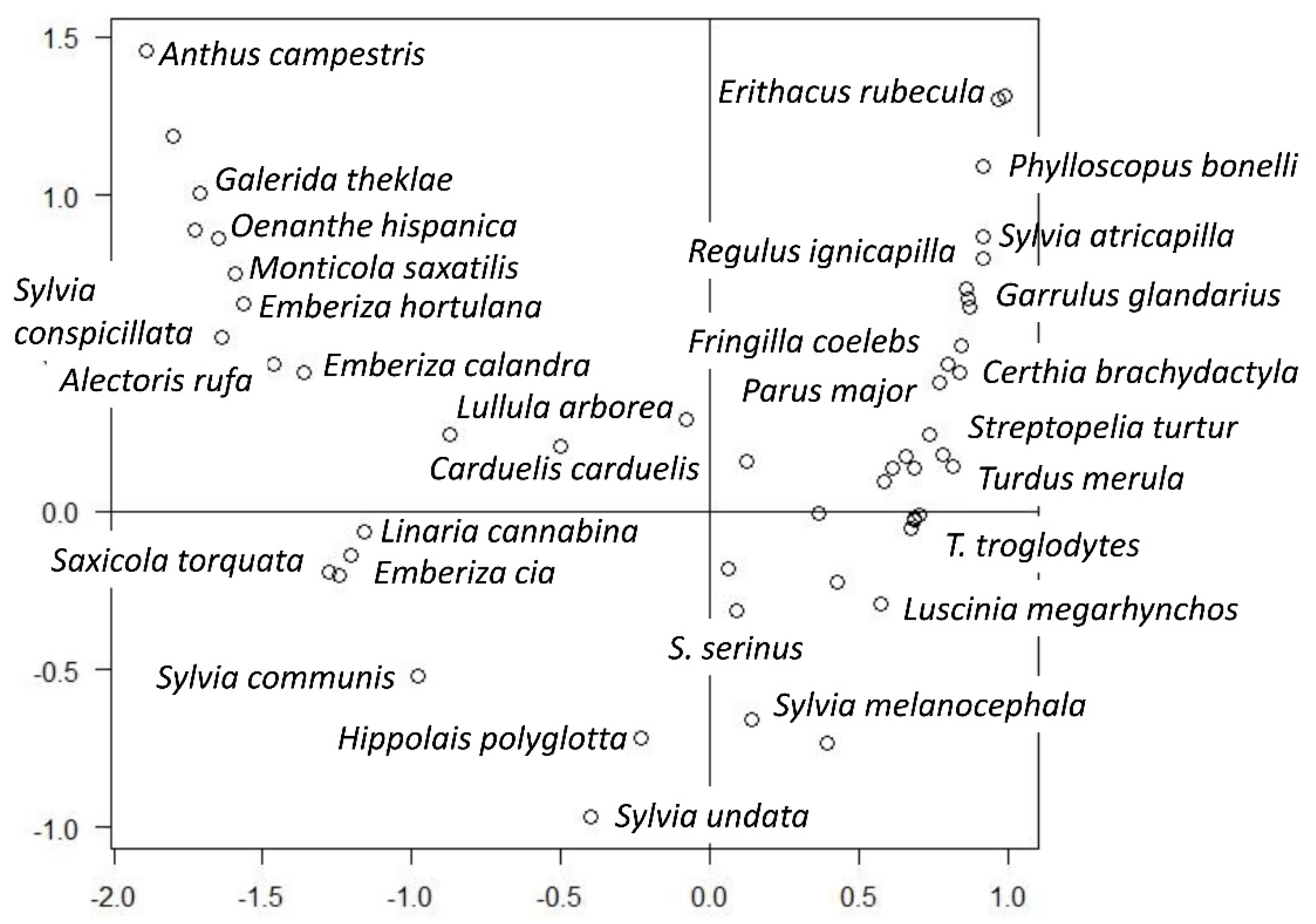
3.2. A Common Successional Gradient
Species Ordination
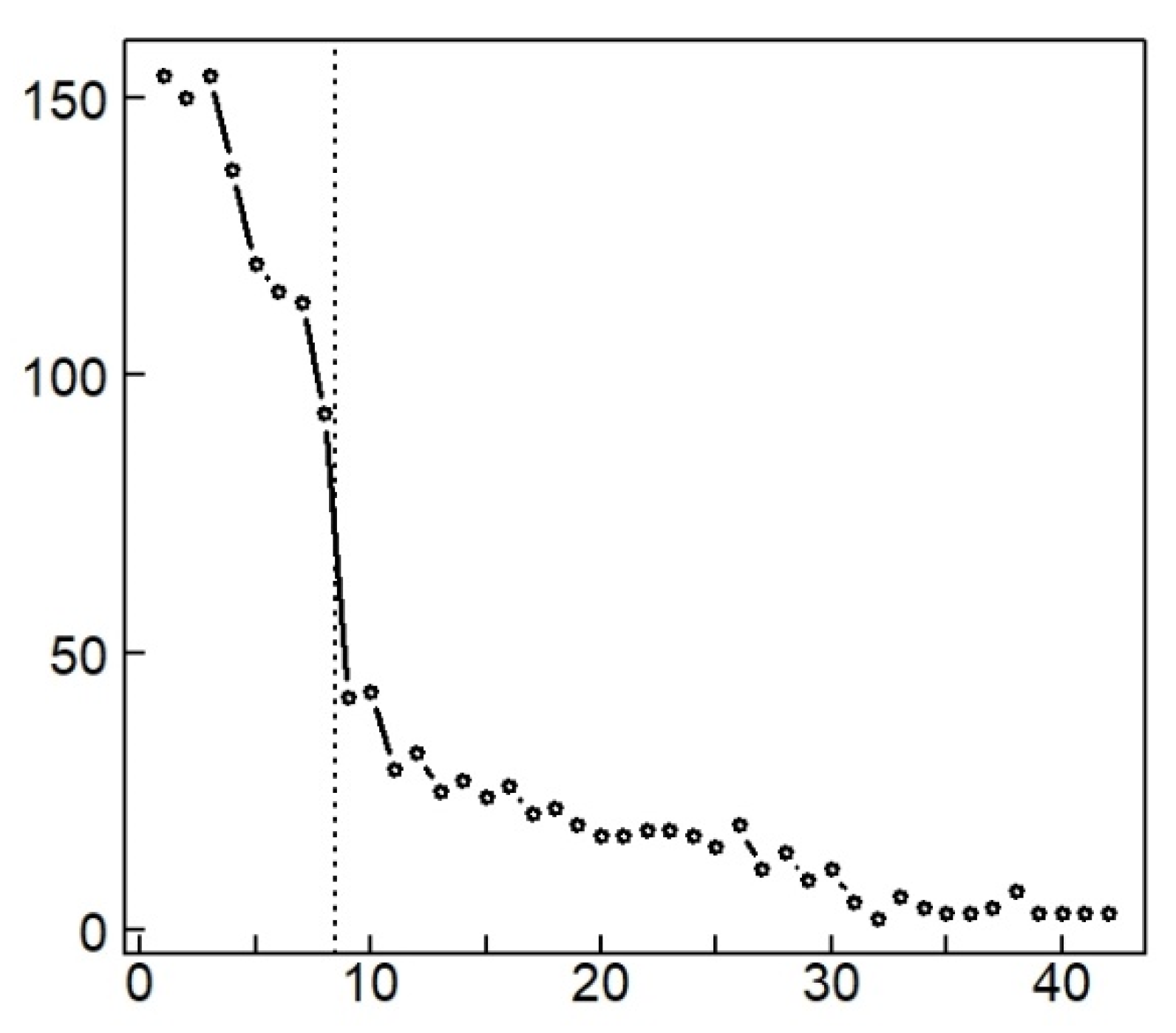
3.3. Postfire Trajectories of the Samples
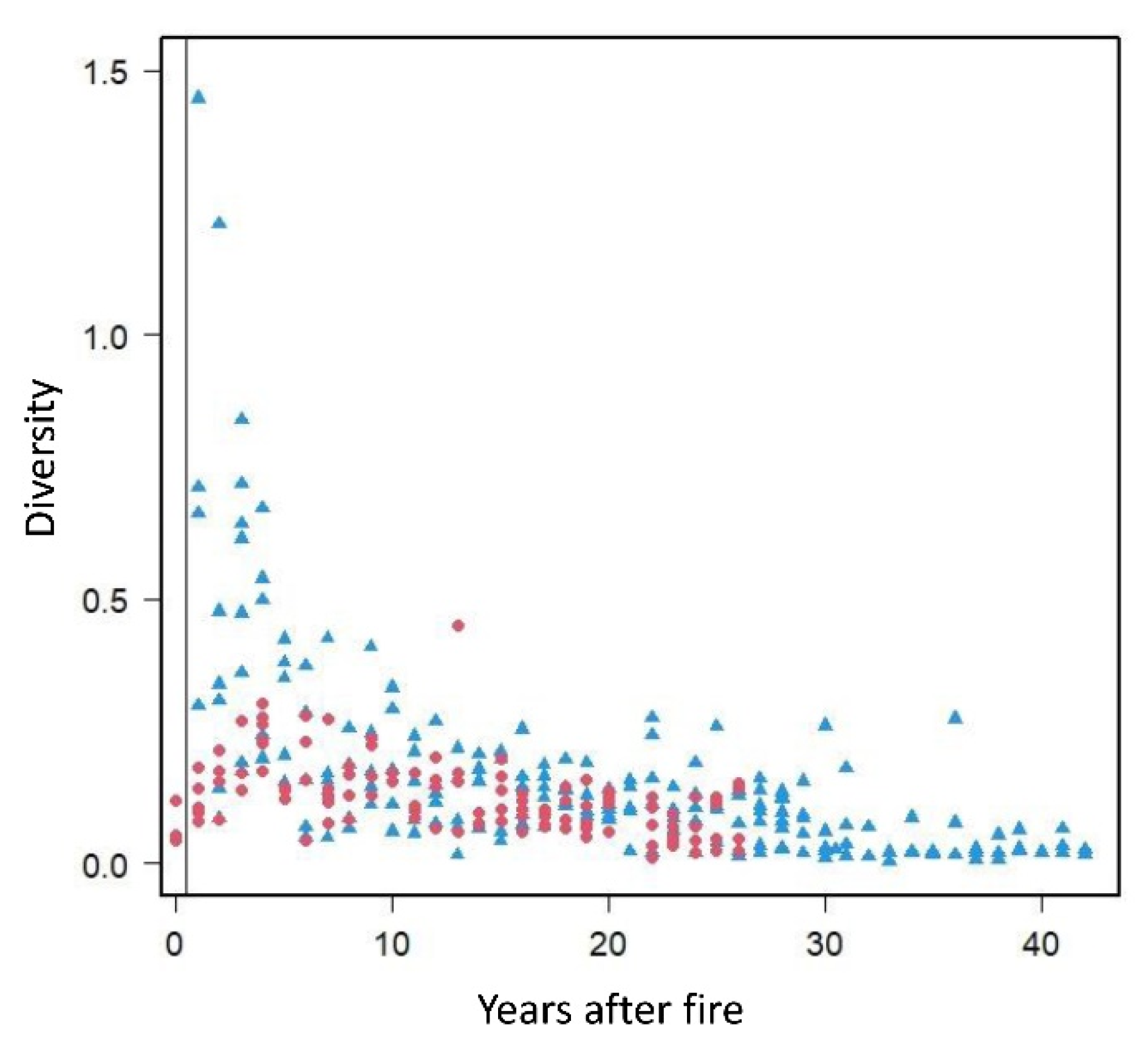
3.4. High Diversity in Recently Burned Forest Stations
3.5. Before–After Study
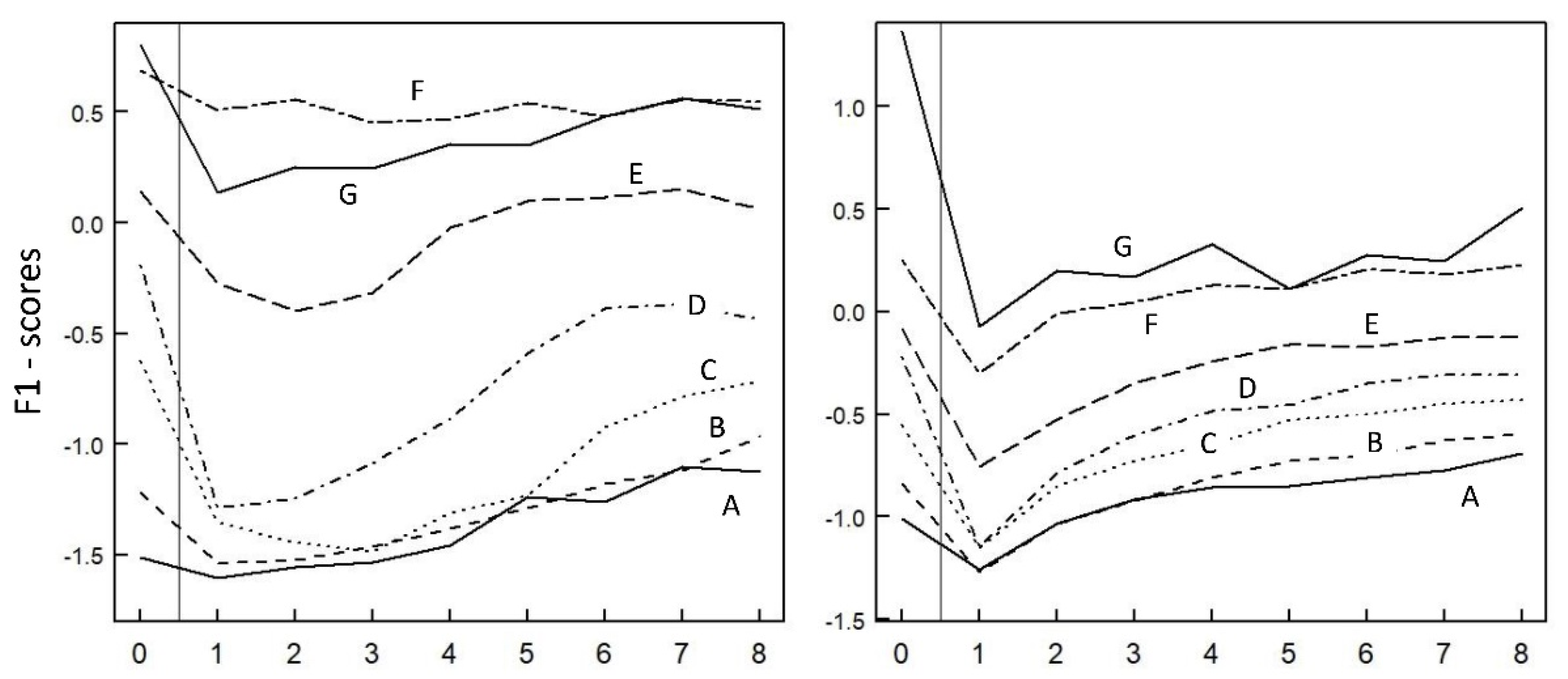
3.5.1. Postfire Successional Pathways
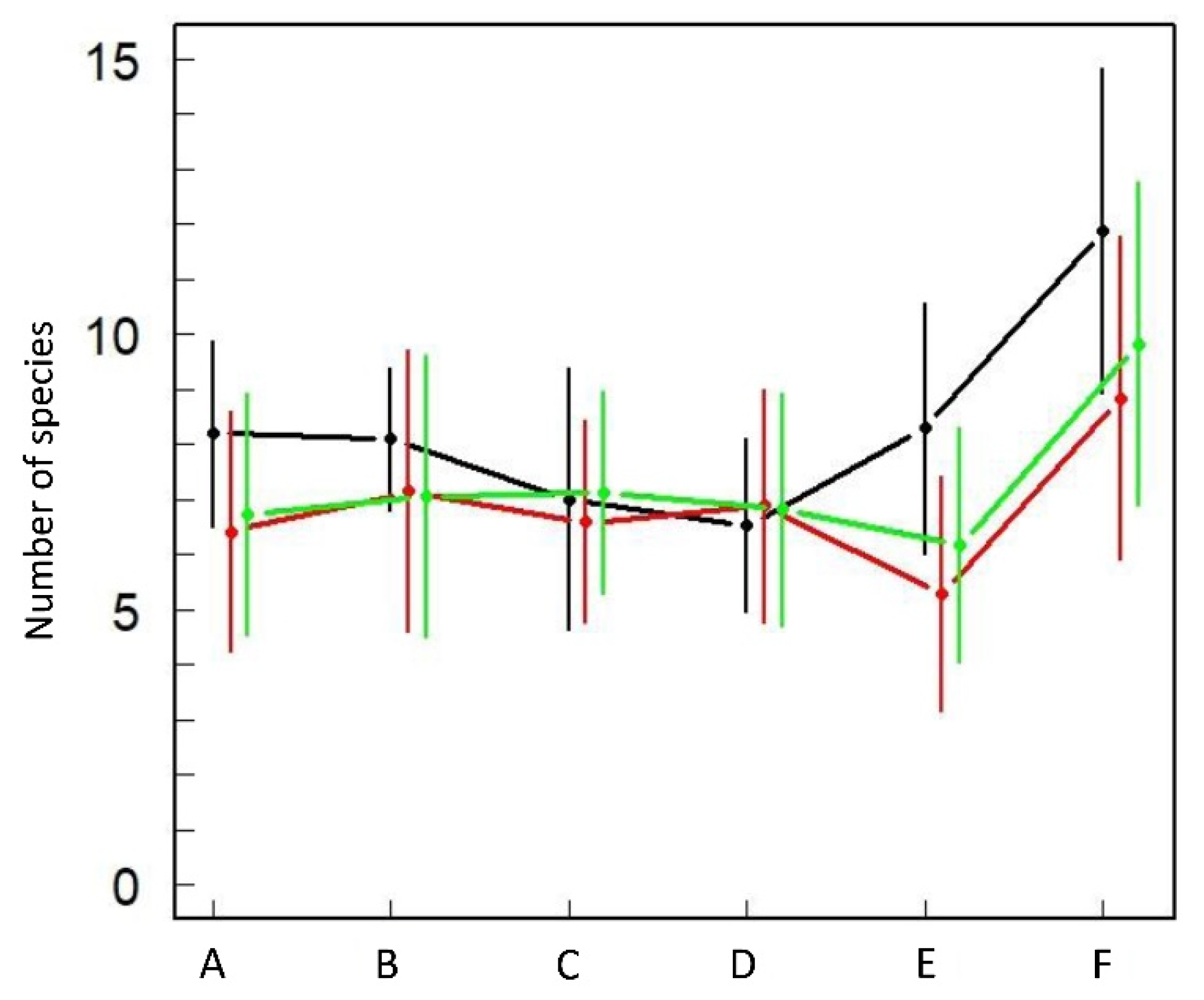
3.5.2. Avifaunal Shifts in the Seven Vegetation Types
4. Discussion
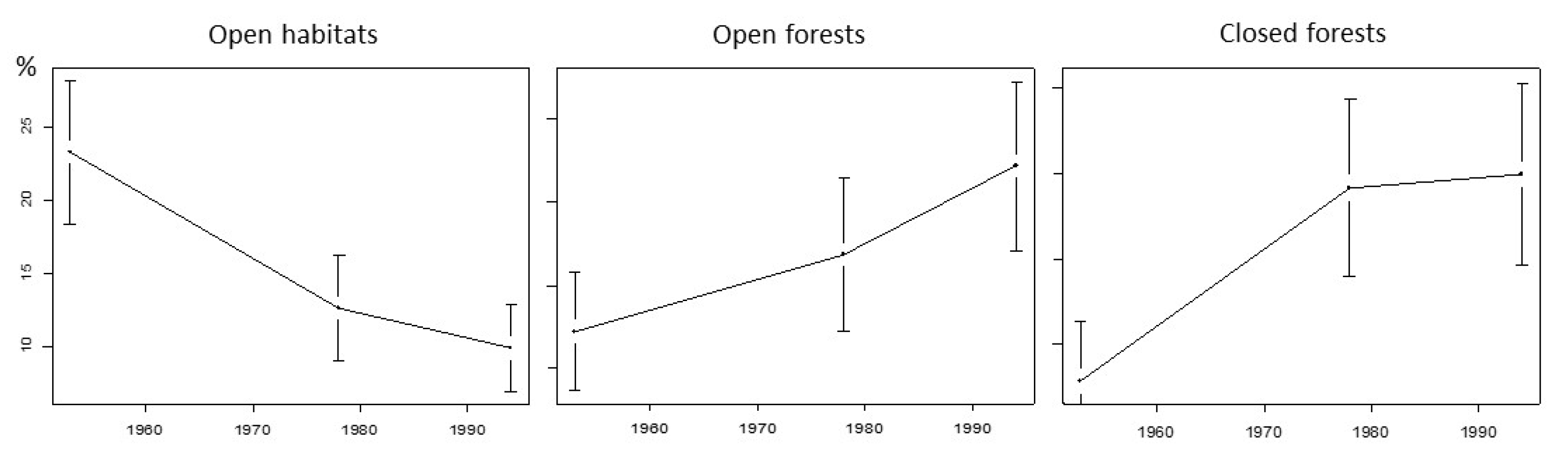
4.1. Return Time and Ratchet Effect
4.2. Three Modalities in the Postfire Successional Gradient
4.3. Variability and Convergence of Successional Pathways
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anthus campestris | −1.8922 | Sylvia hortensis | 0.4283 |
| Coturnix coturnix | −1.7981 | Luscinia megarhynchos | 0.5714 |
| Oenanthe leucura | −1.7267 | Emberiza cirlus | 0.5825 |
| Galerida theklae | −1.7122 | Turdus merula | 0.6151 |
| Oenanthe hispanica | −1.6463 | Jynx torquilla | 0.6571 |
| Sylvia conspicillata | −1.6371 | Cuculus canorus | 0.6773 |
| Monticola saxatilis | −1.5935 | Picus viridis | 0.6852 |
| Emberiza hortulana | −1.5665 | Sylvia cantillans | 0.6853 |
| Alectoris rufa | −1.4619 | Oriolus oriolus | 0.6884 |
| Emberiza calandra | −1.3623 | Troglodytes troglodytes | 0.7019 |
| Saxicola torquata | −1.2745 | Streptopelia turtur | 0.7381 |
| Monticola solitarius | −1.2431 | Parus major | 0.7711 |
| Emberiza cia | −1.2066 | Clumba palumbus | 0.7828 |
| Linaria cannabina | −1.1591 | Fringilla coelebs | 0.7998 |
| Sylvia communis | −0.9809 | Aegithalos caudatus | 0.8138 |
| Pica pica | −0.8698 | Certhia brachydactyla | 0.8371 |
| Carduelis carduelis | −0.4995 | Lophophanes cristatus | 0.8420 |
| Sylvia undata | −0.3991 | Cyanistes caeruleus | 0.8622 |
| Hippolais polyglotta | −0.2333 | Garrulus glandarius | 0.8671 |
| Lullula arborea | −0.0780 | Sylvia borin | 0.8695 |
| Lanius senator | 0.0609 | Sylvia atricapilla | 0.9142 |
| Serinus serinus | 0.0902 | Phylloscopus bonelli | 0.9143 |
| Upupa epops | 0.1227 | Regulus ignicapilla | 0.9174 |
| Sylvia melanocephala | 0.1381 | Sitta europea | 0.9694 |
| Chloris chloris | 0.3629 | Erithacus rubecula | 0.9879 |
| Prunella modularis | 0.3957 |
| Axis 1 | Axis 2 | Axis 3 | Axis 4 | |
|---|---|---|---|---|
| Eigenvalue | 0.72 | 0.33 | 0.4 | 0.19 |
| Inertia % | 13.9 | 6.4 | 4.6 | 3.6 |
| cvR | −0.68 | 0.17 | −0.16 | 0.30 |
| cv025 | −0.46 | −0.56 | −0.25 | −0.08 |
| cv05 | 0.26 | −0.72 | - | −0.12 |
| cv1 | 0.51 | −0.58 | 0.12 | −0.086 |
| cv2 | 0.63 | −0.15 | 0.32 | - |
| cv4 | 0.73 | 0.38 | 0.27 | - |
| cv8 | 0.62 | 0.41 | 0.09 | - |
| cv16 | 0.32 | 0.26 | - | - |






References
- Pickett, S.T.A.; White, P.S. (Eds.) The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: Orlando, FL, USA, 1985; 472p. [Google Scholar]
- Moreira, F.; Viedma, O.; Arianoutsou, M.; Curt, T.; Koutsias, N.; Rigolot, E.; Bilgili, E. Landscape–wildfire interactions in southern Europe: Implications for landscape management. J. Environ. Manag. 2011, 92, 2389–2402. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, P.M.; Monteiro-Henriques, T.; Guiomar, N.; Loureiro, C.; Barros, A.M. Bottom-up variables govern large-fire size in Portugal. Ecosystems 2016, 19, 1362–1375. [Google Scholar] [CrossRef]
- Ascoli, D.; Moris, J.V.; Marchetti, M.; Sallustio, L. Land use change towards forests and wooded land correlates with large and frequent wildfires in Italy. Ann. Silvic. Res. 2021, 46, 177–188. [Google Scholar]
- Van der Maarel, E. Vegetation dynamics: Pattern in time and space. Vegetatio 1988, 77, 7–19. [Google Scholar] [CrossRef]
- Tatoni, T.; Roche, P. Comparison of old-field and forest revegetation dynamics in Provence. J. Veg. Sci. 1994, 5, 295–302. [Google Scholar] [CrossRef]
- Perevolotsky, A.; Seligman, N.G. Role of grazing in Mediterranean rangeland ecosystems: Inversion of a paradigm. BioScience 1998, 48, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.J.; Walker, L.R. Old field succession: Development of concepts. In Old Fields: Dynamics and Restoration of Abandoned Farmland; Cramer, V.A., Hobbs, R.J., Eds.; Island Press: Washington, DC, USA, 2007; pp. 17–30. [Google Scholar]
- Pickett, S.T.A. Space-for-time substitutions as an alternative to long-term studies. In Long-Term Studies in Ecolog: Approaches and Alternatives; Likens, G.E., Ed.; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Pickett, S.T.A.; Collins, S.L.; Armesto, J.J. Models, mechanisms and pathways of succession. Bot. Rev. 1987, 53, 335–371. [Google Scholar] [CrossRef]
- Johson, E.A.; Miyanishi, K. Testing the assumption of chronosequences in succession. Ecol. Lett. 2008, 11, 419–431. [Google Scholar] [CrossRef]
- Walker, L.R.; Wardle, D.A.; Bardgett, R.D.; Clarkson, B.D. The use of chronosequences in studies of ecological succession and soil development. J. Ecol. 2010, 98, 725–736. [Google Scholar] [CrossRef]
- Prodon, R.; Lebreton, J.D. Breeding avifauna of a Mediterranean succession—The holm oak and cork oak series in the eastern Pyrenees, 1. Analysis and Modeling of the Structure Gradient. Oikos 1981, 37, 21–38. [Google Scholar] [CrossRef]
- Lanyon, W.E. Breeding birds and old-field succession on fallow Long Island farmland. Bull. Am. Mus. Nat. Hist. 1981, 168, 1–60. [Google Scholar]
- Bollinger, E.K. Successional changes and habitat selection in hayfield bird communities. Auk 1995, 112, 720–730. [Google Scholar]
- Preiss, E.; Martin, J.-L.; Debussche, M. Rural depopulation and recent landscape changes in a Mediterranean region: Consequences to the breeding avifauna. Landsc. Ecol. 1997, 12, 51–61. [Google Scholar] [CrossRef]
- Sirami, C.; Brotons, L.; Martin, J.-M. Vegetation and songbird response to land abandonment: From landscape to census plot. Div. Distrib. 2007, 13, 42–52. [Google Scholar] [CrossRef]
- Prodon, R. Consequences for avifauna of landscape encroachment by woody vegetation in northern Catalonia. Rev. Catalana d’Ornitol. 2020, 36, 1–9. [Google Scholar]
- Prodon, R.; Fons, R.; Athias-Binche, F. The impact of fire on animal communities in Mediterranean area. In The Role of Fire in Ecological Systems; Trabaud, L., Ed.; SPB Academic Publishing: The Hague, The Netherlands, 1987; pp. 121–157. [Google Scholar]
- Pons, P.; Prodon, R. Short-term temporal pattern in a Mediterranean shrubland bird community after wildfire. Acta Oecol. 1996, 17, 29–41. [Google Scholar]
- Herrando, S.; Brotons, L.; del Amo, R.; Llacuna, S. Bird community succession after fire in a dry Mediterranean shrubland. Ardea 2002, 90, 303–310. [Google Scholar]
- Herrando, S.; Brotons, L.; Llacuna, S. Does fire increase the spatial heterogeneity of bird communities in Mediterranean landscapes? Ibis 2003, 145, 307–317. [Google Scholar] [CrossRef]
- Pons, P.; Clavero, M.; Bas, J.M.; Prodon, R. Time-window of occurrence and vegetation cover preferences of Dartford and Sardinian Warblers after fire. J. Ornitol. 2012, 153, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Prodon, R. Birds and the fire cycle in a resilient Mediterranean forest: Is there any baseline? Forests 2021, 12, 1644. [Google Scholar] [CrossRef]
- Moreira, F.; Ferreira, P.G.; Rego, F.C.; Bunting, S. Landscape changes and breeding bird assemblages in northwestern Portugal: The role of fire. Landsc. Ecol. 2001, 16, 175–187. [Google Scholar] [CrossRef]
- Moreira, F.; Russo, D. Modelling the impact of agricultural abandonment and wildfires on vertebrate diversity in Mediterranean Europe. Landsc. Ecol. 2007, 22, 1461–1476. [Google Scholar] [CrossRef]
- Zozaya, E.L.; Brotons, L.; Saura, S. Recent fire history and connectivity patterns determine bird species distribution dynamics in landscapes dominated by land abandonment. Landsc. Ecol. 2012, 27, 171–184. [Google Scholar] [CrossRef]
- Horn, H.S. Causes of variety in patterns of secondary successions. In Forest Succession: Concepts and Applications; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 1981; pp. 24–35. [Google Scholar]
- Prach, K. Succession of vegetation in abandoned fields in Finland. Ann. Bot. Fenn. 1985, 22, 307–314. [Google Scholar]
- Blondel, J.; Farré, H. The convergent trajectories of bird communities along ecological successions in European forests. Oecologia 1988, 75, 83–93. [Google Scholar] [CrossRef]
- Hobson, K.A.; Schieck, J. Changes in bird communities in boreal mixed-wood forest: Harvest and wildfire effects over 30 years. Ecol. Appl. 1999, 9, 849–863. [Google Scholar] [CrossRef]
- Suárez-Seoane, S.; Osborne, P.E.; Baudry, J. Responses of birds of different biogeographic origins and habitat requirements to agricultural land abandonment in northern Spain. Biol. Conserv. 2002, 105, 333–344. [Google Scholar] [CrossRef]
- Del Moral, R.; Thomason, L.A.; Wenke, A.C.; Lozanoff, N.; Abata, M.D. Primary succession trajectories on pumice at Mount St. Helens, Washington. J. Veg. Sci. 2012, 23, 73–85. [Google Scholar] [CrossRef]
- McIntosh, R.P. The relationship between succession and the recovery process in ecosystems. In The Recovery Process in Damaged Ecosystems; Cairns, J., Jr., Ed.; Ann Arbor Sciences: Ann Arbor, MI, USA, 1980; pp. 11–62. [Google Scholar]
- Young, T.P.; Chase, J.M.; Huddleston, R.T. Community succession and assembly. Comparing, contrasting and combining paradigms in the context of ecological restoration. Ecol. Restor. 2001, 19, 5–18. [Google Scholar] [CrossRef]
- Damgaard, C. A Critique of the space-for-time substitution practice in community ecology. Trends Ecol. Evol. 2019, 34, 416–421. [Google Scholar] [CrossRef]
- Jacquet, K.; Prodon, R. Measuring the postfire resilience of a bird-vegetation system: A 28-year study in a Mediterranean oak woodland. Oecologia 2009, 161, 801–811. [Google Scholar] [CrossRef]
- Di Castri, F. Mediterranean-type shrublands of the world. In Ecosystems of the World 11: Mediterranean-Type Shrublands; Di Castri, F., Goodal, D.W., Specht, R.L., Eds.; Elsevier Publishing: Amsterdam, The Netherlands, 1981; pp. 1–52. [Google Scholar]
- Perelman, R. Perception of Mediterranean landscapes, particularly of maquis landscapes. In Ecosystems of the World 11: Mediterranean-Type Shrublands; Di Castri, F., Goodal, D.W., Specht, R.L., Eds.; Elsevier Publishing: Amsterdam, The Netherlands, 1981; pp. 538–554. [Google Scholar]
- French Environment and Energy Management Agency (ADEME); Météo-France. Changement Climatique au 20ème Siècle en Languedoc-Roussillon. Données et Références. 56p. Available online: https://occitanie.ademe.fr/mediatheque/autres-publications/energie-et-changement-climatique (accessed on 31 October 2022). (In French).
- Becat, J. Les Albères: Mutations économiques d’une montagne méditerranéenne: Montagne-providence du passé, montagne-paysage d’aujourd’hui. Conflent 1973, 61, 1–47. (In French) [Google Scholar]
- Debussche, M.; Lepart, J.; Dervieux, A. Mediterranean landscape changes: Evidence from old postcards. Glob. Ecol. Biogeogr. 1999, 8, 3–15. (In French) [Google Scholar] [CrossRef]
- Marty, P.; Aronson, J.; Lepart, J. Dynamics and restoration of abandoned farmland and other old fields in southern France. In Old Fields: Dynamics and Restoration of Abandoned Farmland; Cramer, V.A., Hobbs, R.J., Eds.; Island Press: Washington, DC, USA, 2007; pp. 202–224. [Google Scholar]
- Diamond, J. Laboratory experiments, field experiments, and natural experiments. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper and Row: New York, NY, USA, 1986; pp. 3–22. [Google Scholar]
- Fukami, T.; Wardle, D.A. Long-term ecological dynamics: Reciprocal insights from natural and anthropogenic gradients. Proc. R. Soc. B 2005, 272, 2105–2115. [Google Scholar] [CrossRef] [Green Version]
- Prodon, R.; Diaz-Delgado, R. Assessing the postfire resilience of a Mediterranean forest from satellite and ground data (NDVI, vegetation structure, avifauna). Écoscience 2021, 28, 81–91. [Google Scholar] [CrossRef]
- Austin, M.P. Use of ordination and other multivariate descriptive methods to study succession. Plant Ecol. 1977, 35, 165–175. [Google Scholar] [CrossRef]
- Malanson, G.P.; Trabaud, L. Ordination analysis of components of resilience of Quercus coccifera garrigue. Ecology 1987, 68, 463–472. [Google Scholar] [CrossRef]
- Halpern, C.B. Early successional pathways and the resistance and resilience of forest communities. Ecology 1988, 69, 1703–1715. [Google Scholar] [CrossRef]
- Prodon, R. Animal communities and vegetation dynamics: Measuring and modelling animal community dynamics along forest successions. In Responses of Forest Ecosystems to Environmental Changes; Teller, A., Mathy, P., Jeffers, J.N.R., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 126–141. [Google Scholar]
- Debussche, M.; Escarré, J.; Lepart, J.; Houssard, C.; Lavorel, S. Changes in Mediterranean plant succession: Old-fields revisited. J. Veg. Sci. 1996, 7, 519–526. [Google Scholar] [CrossRef]
- Podani, J.; Csontos, P.; Tamás, J.; Miklós, I. A new multivariate approach to studying temporal changes of vegetation. Plant Ecol. 2005, 181, 85–100. [Google Scholar] [CrossRef]
- Saïd, S. Floristic and life-form diversity in post-pasture successions on a Mediterranean island (Corsica). Plant Ecol. 2001, 162, 67–76. [Google Scholar] [CrossRef]
- Chessel, D.; Lebreton, J.-D.; Prodon, R. Mesures symétriques d’amplitude d’habitat et de diversité intra-échantillon dans un tableau espèces-relevés: Cas d’un gradient simple. C. R. Acad. Sci. Paris D 1982, 295, 83–88. (In French) [Google Scholar]
- Thioulouse, J.; Chessel, D. A Method for Reciprocal Scaling of Species Tolerance and Sample Diversity. Ecology 1992, 73, 670–680. [Google Scholar] [CrossRef]
- Thioulouse, J.; Dray, S.; Dufour, A.-B.; Siberchicot, A.; Jombart, T.; Pavoine, S. Multivariate Analysis of Ecological Data with ade4; Springer: New York, USA, 2018; 329p. [Google Scholar]
- Jacquet, K. Biodiversité et Perturbations: Dynamique de L’avifaune Après Incendie et ses Relations Avec la Dynamique Végétale. Ph.D. Thesis, Université Montpellier-2, Montpellier, France, 10 November 2006. (In French). [Google Scholar]
- Barton, P.S.; Ikin, K.; Smith, A.L.; MacGregor, C.; Lindenmayer, D.B. Vegetation structure moderates the effect of fire on bird assemblages in a heterogeneous landscape. Landsc. Ecol. 2014, 29, 703–714. [Google Scholar] [CrossRef]
- Wiens, J.A.; Rotenberry, J.T. Response of breeding passerine birds to rangeland alteration in a North American shrubsteppe locality. J. Appl. Ecol. 1985, 22, 655–668. [Google Scholar] [CrossRef]
- Llimona, E.; Matheu, F.; Prodon, R. Role of standing snags and of tree regeneration in postfire bird succession: Comparison of pine and oak forests of Montserrat (Catalonia). In Fire in Mediterranean Ecosystems; Trabaud, L., Prodon, R., Eds.; Ecosystem Research Reports; ECSC-EEC-EAEC: Luxembourg, 1993; Volume 5, pp. 315–331. [Google Scholar]
- Hutto, R.L. Composition of bird communities following stand-replacement fires in northern Rocky Mountain (U.S.A.) conifer forests. Conserv. Biol. 1995, 9, 1041–1058. [Google Scholar] [CrossRef] [Green Version]
- Pons, P.; Henry, P.-Y.; Gargallo, G.; Prodon, R.; Lebreton, J.-D. Local survival after fire in Mediterranean shrublands: Combining capture-recapture data over several bird species. Popul. Ecol. 2003, 45, 187–196. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Blanchard, W.; McBurney, L.; Blair, D.; Banks, S.C.; Driscoll, D.A.; Smith, A.L.; Gill, A.M. Complex responses of birds to landscape-level fire extent, fire severity and environmental drivers. Div. Distrib. 2014, 20, 467–477. [Google Scholar] [CrossRef]
- Santana, V.M.; Baeza, M.J.; Marrs, R.H.; Vallejo, V.R. Old-field secondary succession in SE Spain: Can fire divert it? Plant Ecol. 2010, 211, 337–349. [Google Scholar] [CrossRef]
- Petroselli, A.; Vessella, F.; Cavagnuolo, L.; Piovesan, G.; Schirone, B. Ecological behavior of Quercus suber and Quercus ilex inferred by topographic wetness index (TWI). Trees 2013, 27, 1201–1215. [Google Scholar] [CrossRef]
- Quézel, P.; Médail, F. Ecologie et Biogéographie des Forêts du Bassin Méditerranéen; Elsevier: Paris, France, 2003. (In French) [Google Scholar]
- MacMahon, J.A. Successional processes: Comparisons among biomes with special reference to probable roles of and influences on animals. In Forest Succession: Concepts and Applications; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 1981; pp. 277–304. [Google Scholar]
- Raphael, M.G.; Morrison, M.L.; Yoder-Williams, M.P. Breeding bird populations during twenty-five years of postfire succession in the Sierra Nevada. Condor 1987, 89, 614–626. [Google Scholar] [CrossRef]
- Brotons, L.; Pons, P.; Herrando, S. Colonization of dynamic Mediterranean landscapes: Where do birds come from after fire. J. Biogeogr. 2005, 32, 789–798. [Google Scholar] [CrossRef]
- Brotons, L.; de Cáceres, M.; Fall, A.; Fortin, M.-J. Modeling bird species distribution change in fire prone Mediterranean landscapes: Incorporating species dispersal and landscape dynamics. Ecography 2012, 35, 458–467. [Google Scholar] [CrossRef]
- Prodon, R. Landscape dynamics and bird diversity in the north Mediterranean basin: Conservation issues. In Life and Environment in the Mediterranean; Trabaud, L., Ed.; WIT Press: Southampton, UK; Boston, MA, USA, 2000; pp. 261–299. [Google Scholar]
- Brawn, J.D.; Robinson, S.K.; Thompson, F.R., III. The role of disturbance in the ecology and conservation of birds. Annu. Rev. Ecol. Syst. 2001, 32, 251–276. [Google Scholar] [CrossRef] [Green Version]
- Clavero, M.; Brotons, L.; Herrando, S. Bird community specialization, bird conservation and disturbance: The role of wildfires. J. Anim. Ecol. 2011, 80, 128–136. [Google Scholar] [CrossRef]
- De Caceres, M.; Brotons, L.; Aquilué, N.; Fortin, M.-J. The combined effects of land-use legacies and novel fire regimes on bird distributions in the Mediterranean. J. Biogeogr. 2013, 40, 1535–1547. [Google Scholar] [CrossRef]
- Pons, P.; Lambert, B.; Rigolot, E.; Prodon, R. The effects of grassland management using fire on habitat occupancy and conservation of birds in a mosaic landscape. Biodiv. Conserv. 2003, 12, 1843–1860. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Serrano-Davies, E.; Noguerales, V. Returning home after fire: How fire can help us manage the persistence of scrub-steppe specialist bird population. Biodiv. Conserv. 2018, 27, 3087–3102. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prodon, R. Successional Pathways of Avifauna in a Shifting Mosaic Landscape: Interplay between Land Abandonment and Wildfires. Fire 2022, 5, 183. https://doi.org/10.3390/fire5060183
Prodon R. Successional Pathways of Avifauna in a Shifting Mosaic Landscape: Interplay between Land Abandonment and Wildfires. Fire. 2022; 5(6):183. https://doi.org/10.3390/fire5060183
Chicago/Turabian StyleProdon, Roger. 2022. "Successional Pathways of Avifauna in a Shifting Mosaic Landscape: Interplay between Land Abandonment and Wildfires" Fire 5, no. 6: 183. https://doi.org/10.3390/fire5060183