
The Middle Pleistocene Hippopotamus from Malagrotta (Latium, Italy): New Data and Future Perspectives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
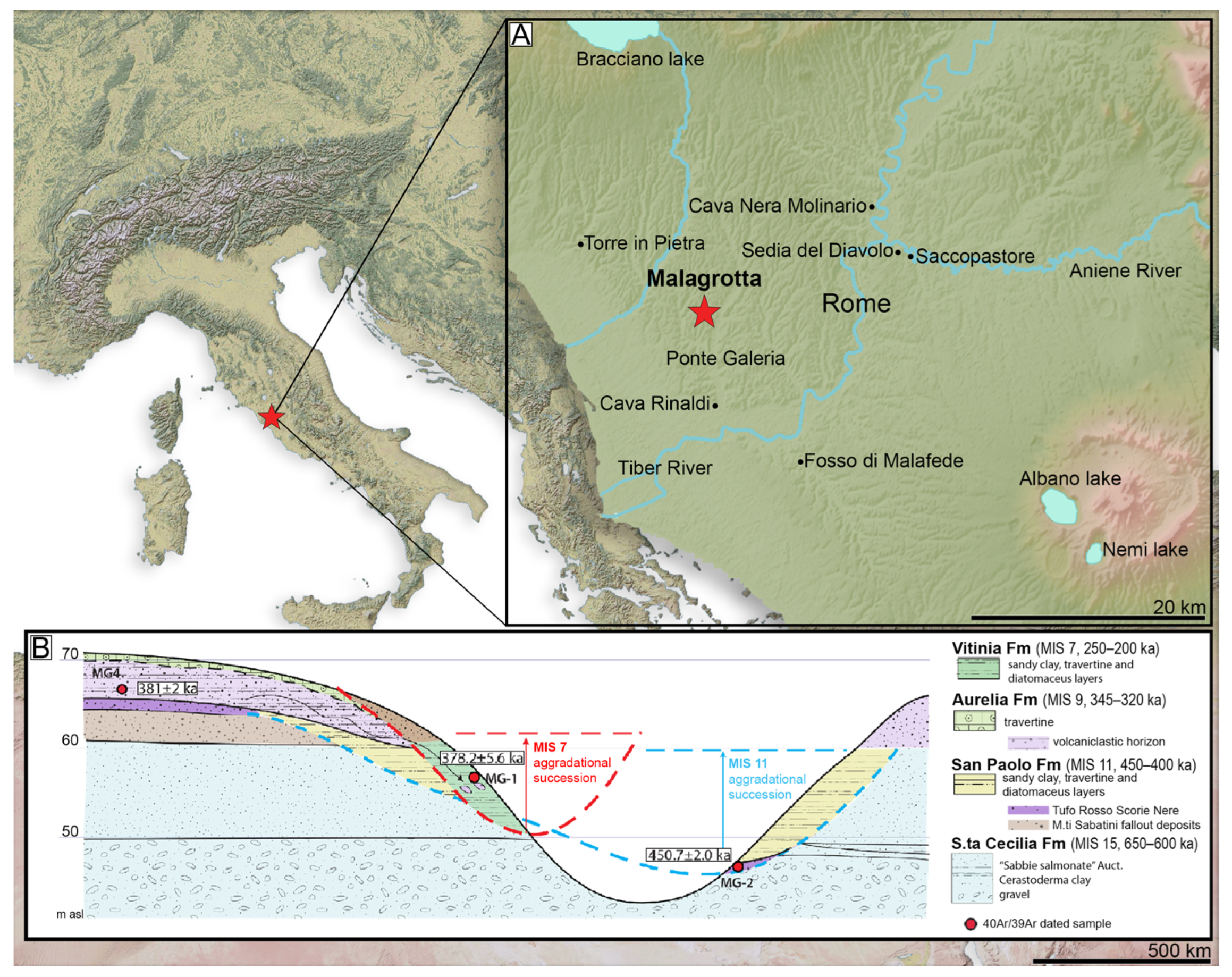
Geological Setting

2. Materials and Methods
3. Systematic Palaeontology
4. Results
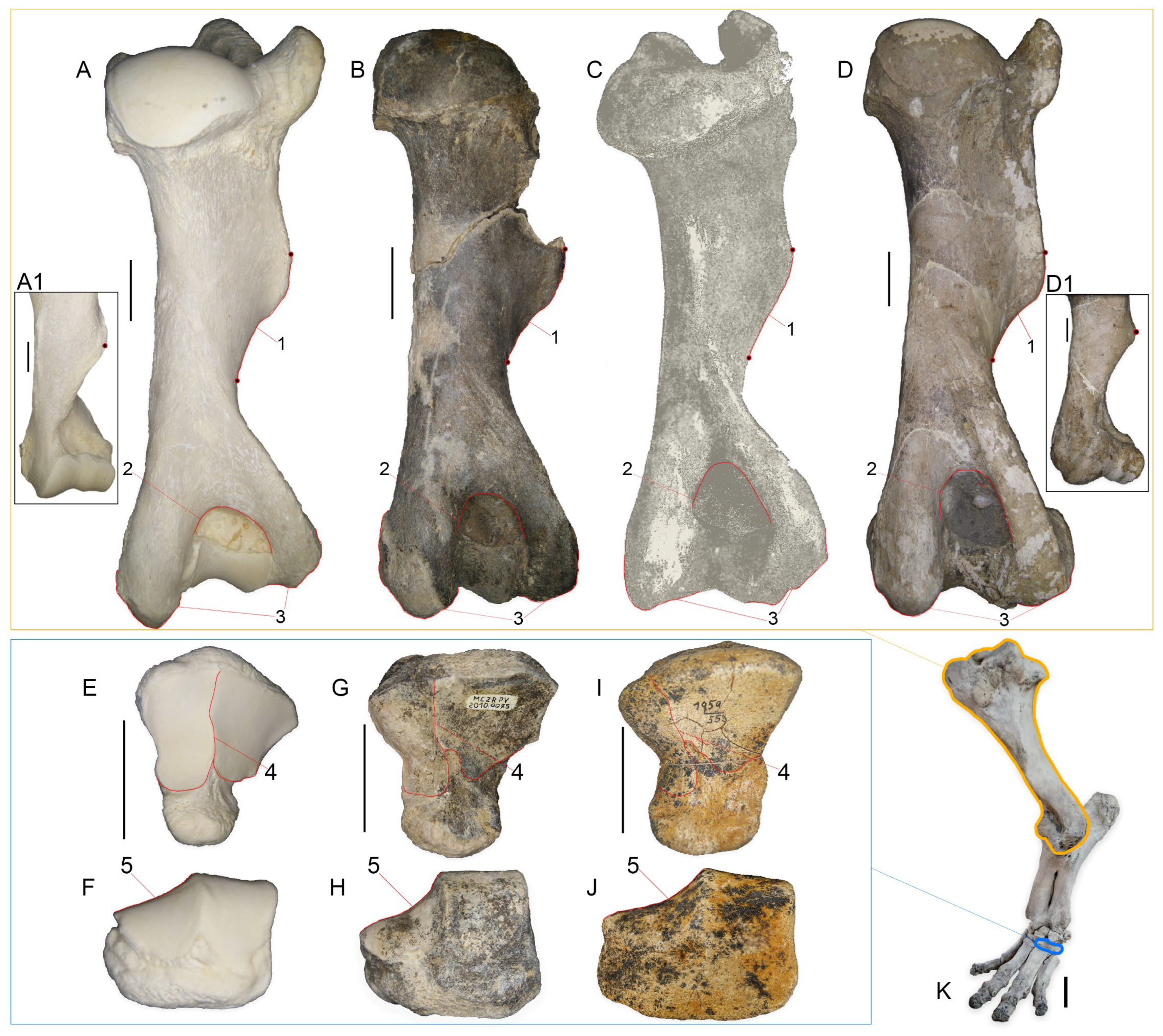
4.1. Morphological Description
4.1.1. Mandible
4.1.2. Teeth
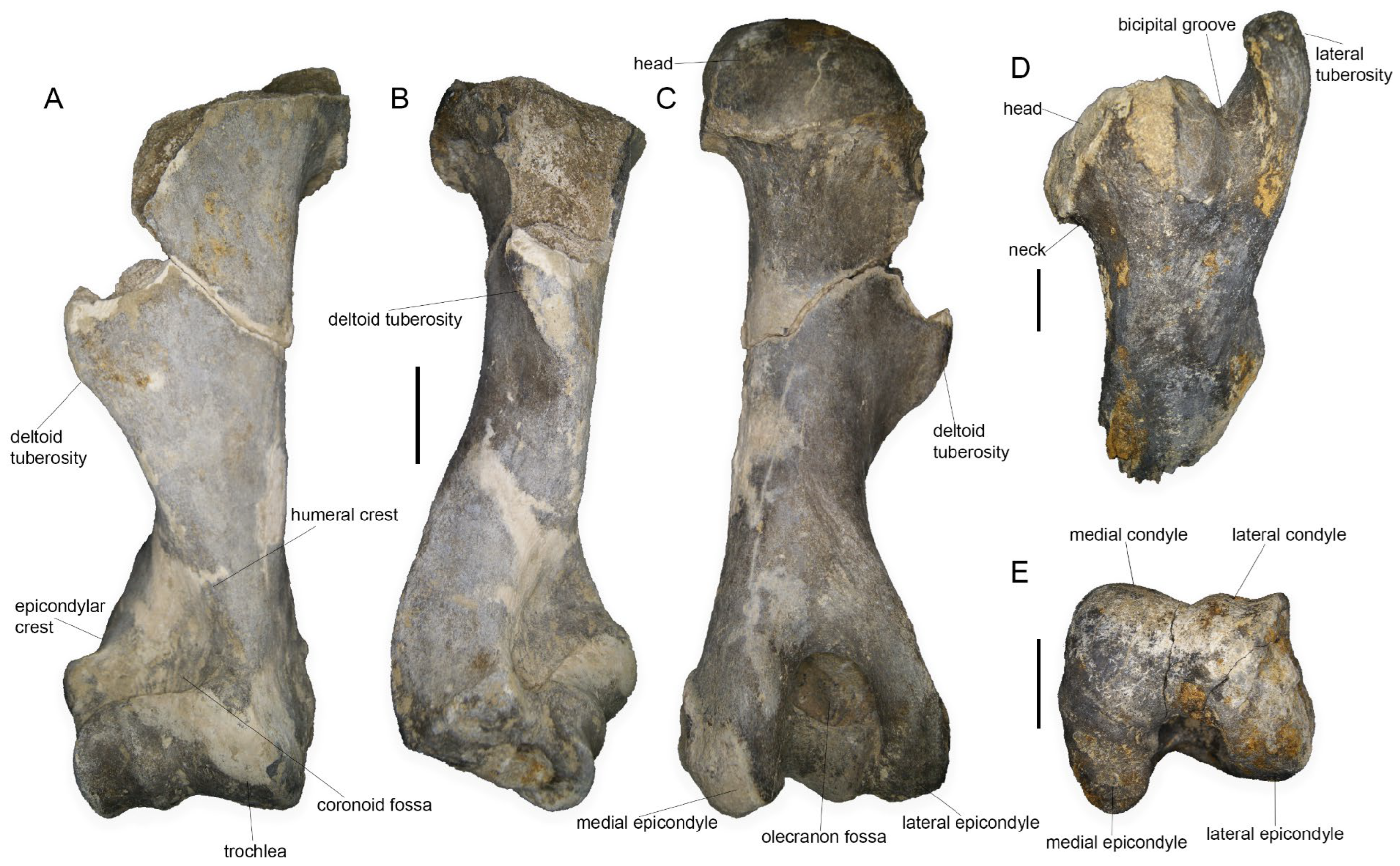
4.1.3. Humerus
4.1.4. Vertebra
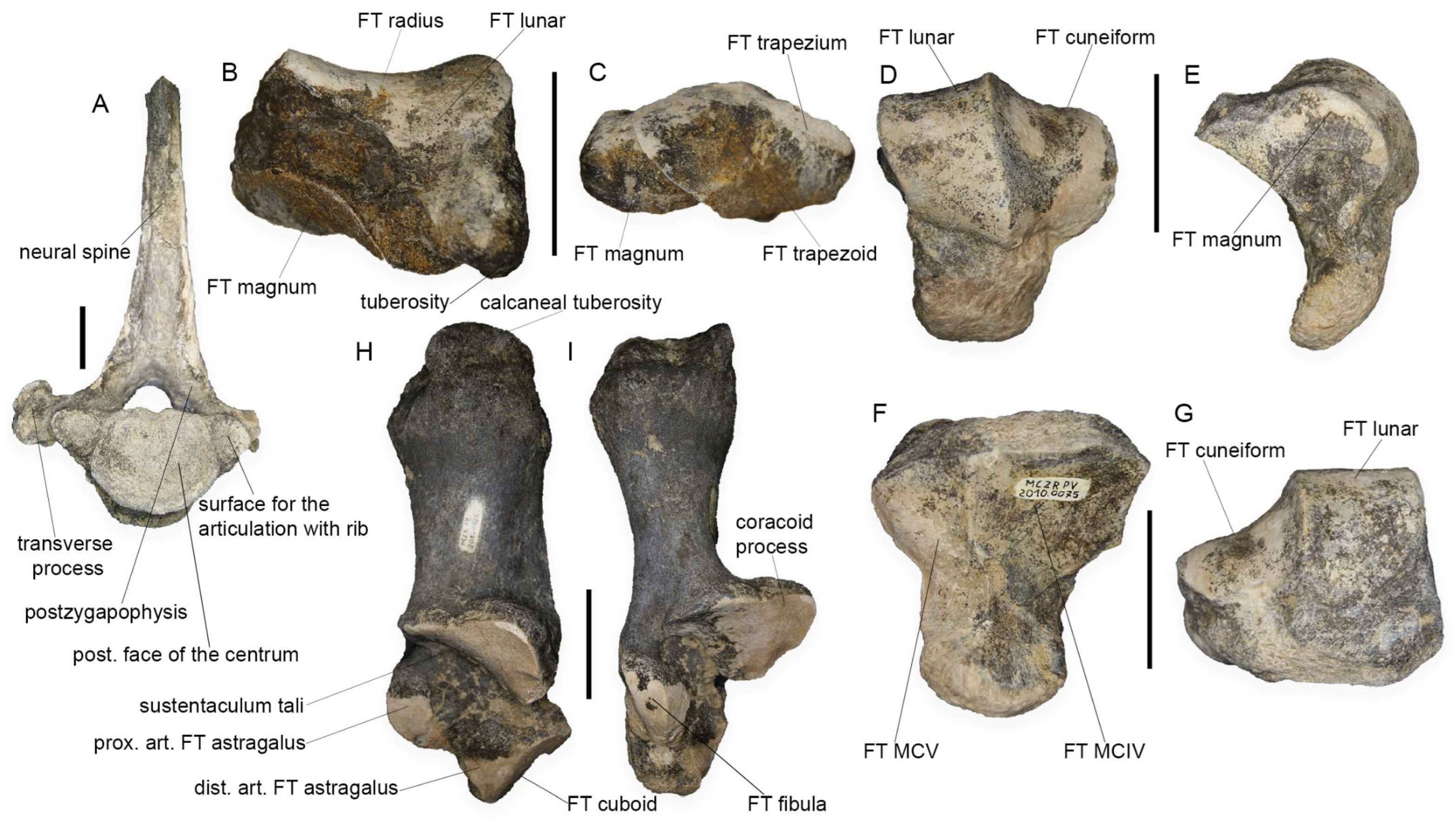
4.1.5. Scaphoid
4.1.6. Unciform
4.1.7. Calcaneum
4.2. Morphological and Morphometric Comparison
4.2.1. Mandible
4.2.2. Teeth
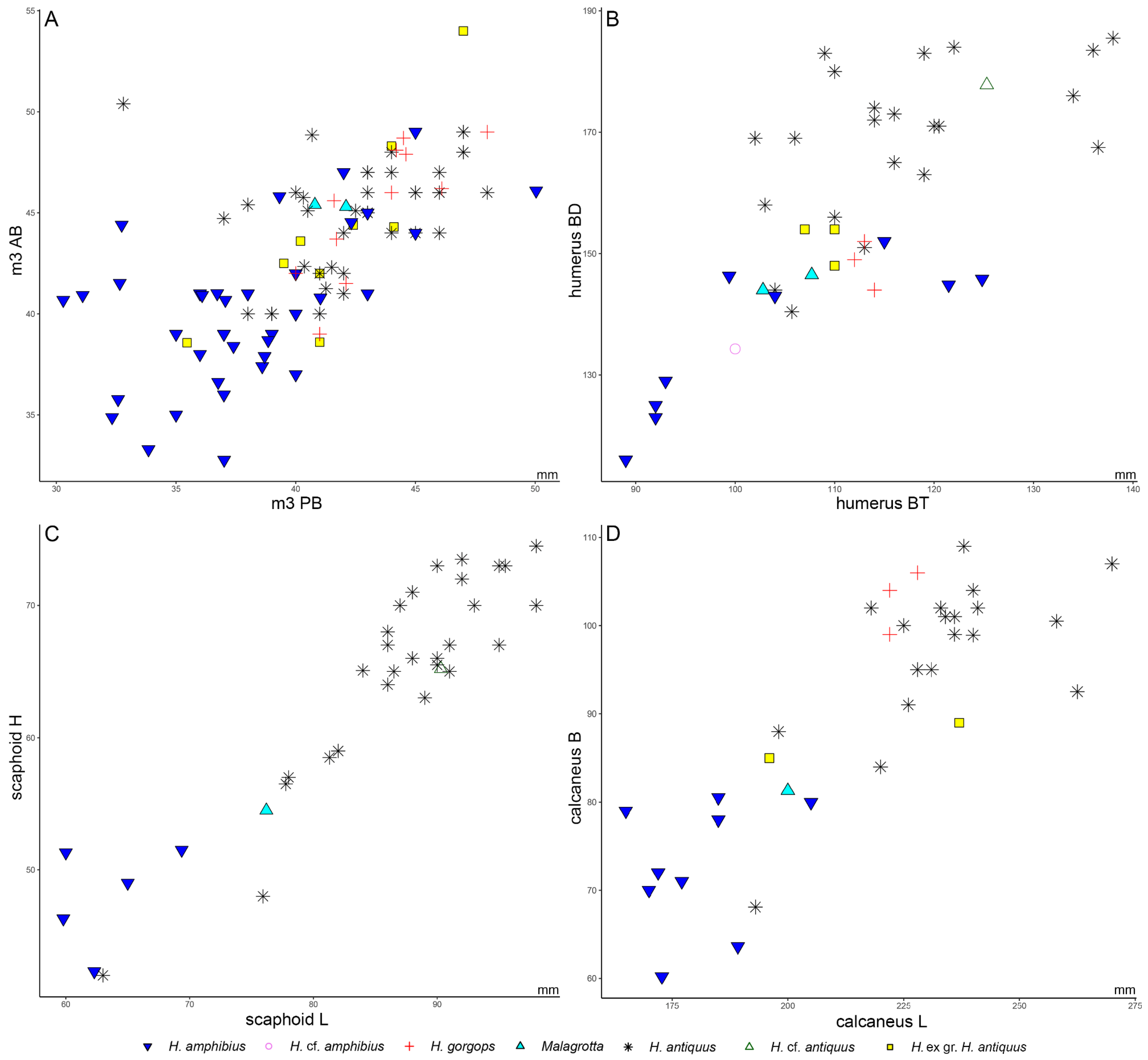
4.2.3. Humerus
4.2.4. Scaphoid
4.2.5. Unciform
4.2.6. Calcaneum
4.3. Taxonomic Attribution
5. Discussion
5.1. Size and Climatic Conditions
5.2. How Many Hippopotamids in Europe during the Pleistocene?
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis; Editio Decima Reformata; Laurentii Salvii: Stockholm, Sweden, 1758; 824p. [Google Scholar]
- Morton, S.G. Additional observations on a new species of hippopotamus of Western Africa (Hippopotamus liberiensis). J. Acad. Nat. Sci. Phila. 1849, 1, 24. [Google Scholar]
- Boisserie, J.-R. The phylogeny and taxonomy of Hippopotamidae (Mammalia: Artiodactyla): A review based on morphology and cladistic analysis. Zool. J. Linn. Soc. 2005, 143, 1–26. [Google Scholar] [CrossRef]
- Desmarest, A.G. Mammalogie, ou Description des Espèces des Mammifères; Seconde Partie; Veuve Agasse: Paris, France, 1822. [Google Scholar]
- Blandamura, F.; Azzaroli, A. L’ “Ippopotamo Maggiore” di Filippo Nesti; Accademia Nazionale dei Lincei-Memorie Sc. Fisiche, ecc: Roma, Italy, 1977; pp. 169–188. [Google Scholar]
- Coryndon, S.C. Fossil Hippopotamidae from the Baringo Basin and relationships within the Gregory Rift, Kenya. Geol. Soc. Lond. Spec. Publ. 1978, 6, 279–292. [Google Scholar] [CrossRef]
- Coryndon, S.C. The Taxonomy and Nomenclature of the Hippopotamidae (Mammalia, Artiodactyla) and a Description of Two New Fossil Species. I. the Nomenclature of the Hippopotamidae; CNRS: Paris, France, 1977. [Google Scholar]
- Faure, M. Les hippopotames quaternaires non-insulaires d’Europe occidentale. Publ. Musée Conflu. 1985, 23, 13–79. [Google Scholar] [CrossRef]
- Kahlke, R.-D. Die Hippopotamus-Reste aus dem Unterpleistozän von Untermassfeld. Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen). Teil 1997, 1, 277–374. [Google Scholar]
- Auguste, P. Évolution des peuplements mammaliens en Europe du Nord-Ouest durant le Pléistocène moyen et supérieur. Le cas de la France septentrionale. Quaternaire. Rev. L’assoc. Fr. Pour L’étude Quat. 2009, 20, 527–550. [Google Scholar] [CrossRef]
- Athanassiou, A. The Fossil Record of Continental Hippopotamids (Mammalia: Artiodactyla: Hippopotamidae) in Greece. In Fossil Vertebrates of Greece; Vlachos, E., Ed.; Springer International Publishing: Cham, Switzerland, 2022; Volume 2, pp. 281–299. [Google Scholar] [CrossRef]
- Adams, N.F.; Candy, I.; Schreve, D.C. An Early Pleistocene hippopotamus from Westbury Cave, Somerset, England: Support for a previously unrecognized temperate interval in the British Quaternary record. J. Quat. Sci. 2022, 37, 28–41. [Google Scholar] [CrossRef]
- Martino, R.; Pandolfi, L. The Quaternary Hippopotamus records from Italy. Hist. Biol. 2022, 34, 1146–1156. [Google Scholar] [CrossRef]
- Fidalgo, D.; Rosas, A.; Bartolini-Lucenti, S.; Boisserie, J.-R.; Pandolfi, L.; Martínez-Navarro, B.; Palmqvist, P.; Rook, L.; Madurell-Malapeira, J. Increase on environmental seasonality through the European Early Pleistocene inferred from dental enamel hypoplasia. Sci. Rep. 2023, 13, 16941. [Google Scholar] [CrossRef]
- Fidalgo, D.; Rosas, A.; Madurell-Malapeira, J.; Pineda, A.; Huguet, R.; García-Tabernero, A.; Cáceres, I.; Ollé, A.; Vallverdú, J.; Saladie, P. A review on the Pleistocene occurrences and palaeobiology of Hippopotamus antiquus based on the record from the Barranc de la Boella Section (Francolí Basin, NE Iberia). Quat. Sci. Rev. 2023, 307, 108034. [Google Scholar] [CrossRef]
- Fidalgo, D.; Malapeira, J.M.; Martino, R.; Pandolfi, L.; Rosas, A. An updated review of the Quaternary hippopotamus fossil record from Iberian Peninsula. Quaternary 2024, 7, 4. [Google Scholar] [CrossRef]
- Fidolini, F.; Ghinassi, M.; Magi, M.; Papini, M.; Sagri, M. The Plio-Pleistocene fluvio-lacustrine Upper Valdarno Basin (central Italy): Stratigraphy and basin fill evolution. Ital. J. Geosci. 2013, 132, 13–32. [Google Scholar]
- Konidaris, G.; Tourloukis, V.; Boni, G.; Athanassiou, A.; Giusti, D.; Thompson, N.; Syrides, G.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Marathousa 2: A New Middle Pleistocene Locality in the Megalopolis Basin (Greece) with Evidence of Hominin Exploitation of Megafauna (Hippopotamus). Paleoanthropology 2023, 2023, 34–55. [Google Scholar]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Tentori, D.; Cavasinni, C.; Conti, J.; Messina, M.Y.; Sarra, A.; Sardella, R. Reinforcing the idea of an early dispersal of Hippopotamus amphibius in Europe: Restoration and multidisciplinary study of the skull from the Middle Pleistocene of Cava Montanari (Rome, central Italy). PLoS ONE 2023, 18, e0293405. [Google Scholar] [CrossRef]
- Mazza, P.P.A. Interrelations between Pleistocene hippopotami of Europe and Africa. Boll. Soc. Paleont. Ital. 1991, 30, 153–186. [Google Scholar]
- Mazza, P.P.A. New evidence on the Pleistocene hippopotami of western Europe. Geol. Romana 1995, 31, 61–241. [Google Scholar]
- Petronio, C. Note on the taxonomy of Pleistocene hippopotamuses. Ibex 1995, 3, e55. [Google Scholar]
- Mazza, P.P.A.; Bertini, A. Were Pleistocene hippopotamuses exposed to climate-driven body size changes?: Pleistocene hippopotamuses. Boreas 2013, 42, 194–209. [Google Scholar] [CrossRef]
- van der Made, J.; Sahnouni, M.; Boulaghraief, K. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In Proceedings of the II Meeting of African Prehistory, Burgos, Spain, 15–16 April 2015; pp. 15–16. [Google Scholar]
- Palombo, M.R.; Raia, P.; Giovinazzo, C. Early-Middle Pleistocene structural changes in mammalian communities from the Italian peninsula. Geol. Soc. Lond. Spec. Publ. 2005, 247, 251–262. [Google Scholar] [CrossRef]
- Masini, F.; Sala, B. Large- and small-mammal distribution patterns and chronostratigraphic boundaries from the Late Pliocene to the Middle Pleistocene of the Italian peninsula. Quat. Int. 2007, 160, 43–56. [Google Scholar] [CrossRef]
- Marra, F.; Nomade, S.; Pereira, A.; Petronio, C.; Salari, L.; Sottili, G.; Bahain, J.-J.; Boschian, G.; Di Stefano, G.; Falguères, C.; et al. A review of the geologic sections and the faunal assemblages of Aurelian Mammal Age of Latium (Italy) in the light of a new chronostratigraphic framework. Quat. Sci. Rev. 2018, 181, 173–199. [Google Scholar] [CrossRef]
- Ceruleo, P.; Rolfo, M.F.; Petronio, C.; Salari, L. Review of Middle Pleistocene archaeological and biochronological data in Malagrotta-Ponte Galeria area (Rome, Central Italy). Atti Soc. Toscana Sci. Nat. Mem. Ser. A 2021, 128, 85–103. [Google Scholar]
- Caloi, L.; Palombo, M.R.; Petronio, C. Resti cranici di Hippopotamus antiquus (=H. major) e Hippopotamus amphibius conservati nel Museo di Paleontologia dell’Università di Roma. Geol. Romana 1980, 19, 91–119. [Google Scholar]
- Caloi, L.; Palombo, M.R. Megaceros savini e Megaceros cf. verticornis (Cervidae) del Pleistocene medio-inferiore di Ponte Galeria (Roma). Geol. Romana 1980, 19, 121–130. [Google Scholar]
- Capasso Barbato, L.; Minieri, M.R. Nuovi resti di carnivori del Pleistocene medio dei dintorni di Roma. Geol. Romana 1987, 26, 1–15. [Google Scholar]
- Anzidei, A.P.; Caloi, L.; Giacopini, L.; Mantero, D.; Palombo, M.R.; Sebastiani, R.; Segre, A.G. Saggi di scavo nei depositi pleistocenici del Km 18,900 della Via Aurelia e di Collina Barbattini (Castel di Guido-Roma). Quad. Archeol. Etrusco-Ital. 1993, 21, 81–90. [Google Scholar]
- Petronio, C. Nuovi resti di ippopotamo del Pleistocene medio-inferiore dei dintorni di Roma e problemi di tassonomia e filogenesi del gruppo. Geol. Romana 1986, 25, 63–73. [Google Scholar]
- Caloi, L.; Palombo, M.R. Resti di mammiferi del Pleistocene medio di Malagrotta (Roma). Boll. Serv. Geol. D’italia 1980, 100, 141–188. [Google Scholar]
- Cassoli, P.F.; De Giuli, C.; Radmilli, A.M.; Segre, A.G. Giacimento del paleolitico inferiore a Malagrotta (Roma). In Atti Della XXIII Riunione Scientifica Dell’Istituto Italiano Di Preistoria e Protostoria; Istituto Italiano di Preistoria e Protostoria: Firenze, Italy, 1982; pp. 531–549. [Google Scholar]
- Villa, P.; Boschian, G.; Pollarolo, L.; Saccà, D.; Marra, F.; Nomade, S.; Pereira, A. Elephant bones for the Middle Pleistocene toolmaker. PLoS ONE 2021, 16, e0256090. [Google Scholar] [CrossRef]
- Marra, F.; Pereira, A.; Boschian, G.; Nomade, S. MIS 13 and MIS 11 aggradational successions of the Paleo-Tiber delta: Geochronological constraints to sea-level fluctuations and to the Acheulean sites of Castel di Guido and Malagrotta (Rome, Italy). Quat. Int. 2022, 616, 1–11. [Google Scholar] [CrossRef]
- Karner, D.; Marra, F. Correlation of fluviodeltaic aggradational sections with glacial climate history: A revision of the Pleistocene stratigraphy of Rome. Geol. Soc. Am. Bull. 1998, 110, 748–758. [Google Scholar] [CrossRef]
- Boisserie, J.-R.; Lihoreau, F.; Orliac, M.; Fisher, R.E.; Weston, E.M.; Ducrocq, S. Morphology and phylogenetic relationships of the earliest known hippopotamids (Cetartiodactyla, Hippopotamidae, Kenyapotaminae): Kenyapotamus and hippopotamid origins. Zool. J. Linn. Soc. 2010, 158, 325–366. [Google Scholar] [CrossRef]
- Reynolds, S.H. A Monograph on the British Pleistocene Mammalia VOL. III, Part I. Hippopotamus. Pages 1–38; Plates I–VI. Monogr. Palaeontogr. Soc. 1922, 74, 1–38. [Google Scholar] [CrossRef]
- Hooijer, D.A. The fossil Hippopotamidae of Asia, with notes on the recent species. Zool. Verh. 1950, 8, 1–124. [Google Scholar]
- Hooijer, D.A.; Singer, R. The fossil hippopotamus from Hopefield, South Africa. Zool. Meded. 1961, 37, 157–165. [Google Scholar]
- Melentis, J.K. Über Hippopotamus antiquus DESMAREST aus dem Mittelpleistozän des Beckens von Megalopolis in Peloponnes (Griechenland). Ann. Geol. Pays Hell. 1965, 16, 403–435. [Google Scholar]
- Harris, J.M. Family Hippopotamidae. In Koobi Fora Research Project; Oxford Academic: Oxford, UK, 1991; Volume 3, pp. 31–85. [Google Scholar]
- Galobart, À.; Blaya, E.; Madurell, J.; Ros, X.; Maroto, J. Resultats de la segonda fase d’actuacions (1999–2005) als jaciments paleontòlogics del Plistocèn inferior d’Incarcal. Paleontol. I Evol. 2003, 34, 153–173. [Google Scholar]
- Tsoukala, E.; Chatzopoulou, K. A new Early Pleistocene (latest Villafranchian) site with mammals in Kalamotó (Mygdonia Basin, Macedonia, Greece)–preliminary report. Mitt. Komm. Quartärforsch. Österr. Akad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Martinez-Navarro, B.; Rook, L.; Papini, M.; Libsekal, Y. A new species of bull from the Early Pleistocene paleoanthropological site of Buia (Eritrea): Parallelism on the dispersal of the genus Bos and the Acheulian culture. Quat. Int. 2010, 212, 169–175. [Google Scholar] [CrossRef]
- Athanassiou, A.; Michailidis, D.; Vlachos, E.; Tourloukis, V.; Thompson, N.; Harvati, K. Pleistocene vertebrates from the Kyparíssia lignite mine, Megalopolis Basin, S. Greece: Testudines, Aves, Suiformes. Quat. Int. 2018, 497, 178–197. [Google Scholar] [CrossRef]
- Mecozzi, B. The Hippopotamus remains from the latest Early Pleistocene site of Cava Redicicoli (Rome, central Italy). Boll. Soc. Paleontol. Ital. 2023, 62, 264. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’, Version 2. Create Elegant Data Visualisations Using the Grammar of Graphics. R Foundation: Vienna, Austria, 2016; pp. 1–189.
- Wickham, H.; Hester, J.; Chang, W.; Bryan, J. R Package, Version 1. Devtools: Tools to Make Developing R Packages Easier. R Foundation: Vienna, Austria, 2016. Available online: https://cran.r-project.org/web/packages/devtools/devtools.pdf(accessed on 8 February 2024).
- Mazo, A.V. Los hipopótamos del Pleistoceno medio de Huescar-1 (Granada). Trab. Neog. Cuantern 1989, 1, 317–325. [Google Scholar]
- Georgitsis, M.K.; Liakopoulou, D.Ε.; Theodorou, G.E. Morphofunctional examination of the carpal bones of pygmy hippopotamus from Ayia Napa, Cyprus. Anat. Rec. 2022, 305, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Georgitsis, M.K.; Liakopoulou, D.Ε.; Theodorou, G.E.; Tsiolakis, E. Functional morphology of the hindlimb of fossilized pygmy hippopotamus from Ayia Napa (Cyprus). J. Morphol. 2022, 283, 1048–1079. [Google Scholar] [CrossRef] [PubMed]
- Kahlke, R.D. Die Unterpleistozäne Komplexfundstelle Untermassfeld-Zusammenfassung des Kenntnisstandes sowie synthetische Betrachtungen zu Genesemodell, Paläoökologie und Stratigraphie. Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen). Teil 2001, 3, 931–1030. [Google Scholar]
- Kahlke, R.-D.; Gaudzinski, S. The blessing of a great flood: Differentiation of mortality patterns in the large mammal record of the Lower Pleistocene fluvial site of Untermassfeld (Germany) and its relevance for the interpretation of faunal assemblages from archaeological sites. J. Archaeol. Sci. 2005, 32, 1202–1222. [Google Scholar] [CrossRef]
- Mazza, P.P.A.; Ventra, D. Pleistocene debris-flow deposition of the hippopotamus-bearing Collecurti bonebed (Macerata, Central Italy): Taphonomic and paleoenvironmental analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 296–314. [Google Scholar] [CrossRef]
- Martínez Navarro, B.; Palmqvist, P.; Madurell, J.; Ros Montoya, S.; Espigares, M.P.; Torregrosa, V.; Pérez Claros, J.A.; Abbazzi, L.; Alberdi, M.T.; Lacombat, F.Y. La fauna de grandes mamíferos de Fuente Nueva-3 y Barranco León-5: Estado de la cuestión. In Proceedings of the Convegno XXIII Jornadas de la Sociedad Española de Paleontología Tenutosi a Caravaca, Cruz, Spain, 3–6 October 2007. [Google Scholar]
- Agostini, S.; Bertini, A.; Caramiello, S.; De Flavis, A.G.; Mazza, P.P.A.; Rossi, M.A.; Satolli, S.; Coccioni, R.; Marsili, A. A new mammalian bone bed from the lower Middle Pleistocene of Ortona (Chieti, Abruzzo, central Italy). Proc. Giornate Paleontol. 2005, 12, 1–5. [Google Scholar]
- Kierdorf, U.; Kahlke, R.D. Pathological Findings on Remains of Hippopotamids from the Early Pleistocene Site of Untermassfeld; Part 4; The Pleistocene of Untermassfeld near Meiningen: Thüringen, Germany, 2020. [Google Scholar]
- Di Stefano, G.; Petronio, C.; Sardella, R. Biochronology of the Pleistocene mammal faunas from Rome urban area. Il Quat. 1998, 11, 191–199. [Google Scholar]
- Luberti, G.M.; Marra, F.; Florindo, F. A review of the stratigraphy of Rome (Italy) according to geochronologically and paleomagnetically constrained aggradational successions, glacio-eustatic forcing and volcano-tectonic processes. Quat. Int. 2017, 438, 40–67. [Google Scholar] [CrossRef]
- Funiciello, R.; Giordano, G. La nuova carta geologica di Roma: Litostratigrafia e organizzazione stratigrafica. La geologia di Roma dal centro storico alla periferia. Mem. Descr. Carta Geol. D’italia 2008, 80, 39–85. [Google Scholar]
- Marra, F.; Rosa, C. Stratigrafia e assetto geologico dell’area romana. Mem. Descr. Carta Geol. D’italia 1995, 50, 49–118. [Google Scholar]
- van der Made, J. Ungulados de Gran Dolina, Atapuerca: Nuevos datos e interpretaciones. Atapuerca Evol. Humana 1998, 97–110. Available online: https://scholar.google.com/scholar_lookup?title=Ungulados%20de%20Gran%20Dolina%2C%20Atapuerca%3A%20nuevos%20datos%20e%20interpretaciones&author=J.%20van%20der%20Made&publication_year=1998&pages=97-109 (accessed on 8 February 2024).
- Falguères, C.; Bahain, J.-J.; Yokoyama, Y.; Arsuaga, J.L.; de Castro, J.M.B.; Carbonell, E.; Bischoff, J.L.; Dolo, J.-M. Earliest humans in Europe: The age of TD6 gran Dolina, Atapuerca, Spain. J. Hum. Evol. 1999, 37, 343–352. [Google Scholar] [CrossRef]
- Álvarez-Posada, C.; Parés, J.M.; Sala, R.; Viseras, C.; Pla-Pueyo, S. New magnetostratigraphic evidence for the age of Acheulean tools at the archaeo-palaeontological site “Solana del Zamborino” (Guadix–Baza Basin, S Spain). Sci. Rep. 2017, 7, 13495. [Google Scholar] [CrossRef]
- Baena Escudero, R.; Proença Cunha, P.; Guerrero Amador, I.; Fernández Caro, J.J.; Cortés Sánchez, M.; Silva Barroso, P.G.; Simón Vallejo, M.D.; Sean, M.A.; Buylaert, J.P. El Pleistoceno Medio y Superior en la Secuencia Ceneral de las Terrazas del Guadalquivir (sgtg): Nuevas Dataciones por Luminiscencia de la t12 y t13 en la Rinconada (SW de España); idUS: Sevilla, Spain, 2019. [Google Scholar]
- Blain, H.-A.; Santonja, M.; Pérez-González, A.; Panera, J.; Rubio-Jara, S. Climate and environments during Marine Isotope Stage 11 in the central Iberian Peninsula: The herpetofaunal assemblage from the Acheulean site of Áridos-1, Madrid. Quat. Sci. Rev. 2014, 94, 7–21. [Google Scholar] [CrossRef]
- Rodríguez, E.S. Artiodáctilos y Proboscídeos de los yacimientos de Áridos de Arganda (Madrid). In Ocupaciones Achelenses en el Valle del Jarama (Arganda-Madrid); Fundacion Dialnet: Logroño, Spain, 1980; pp. 207–230. [Google Scholar]
- Palombo, M.R.; Valli, A.M.F. Remarks on the biochronology of mammalian faunal complexes from the Pliocene to the Middle Pleistocene in France. Geol. Romana 2003, 37, 145–163. [Google Scholar]
- Limondin-Lozouet, N.; Nicoud, E.; Antoine, P.; Auguste, P.; Bahain, J.-J.; Dabkowski, J.; Duperon, J.; Duperon, M.; Falgueres, C.; Ghaleb, B. Oldest evidence of Acheulean occupation in the Upper Seine valley (France) from an MIS 11 tufa at La Celle. Quat. Int. 2010, 223, 299–311. [Google Scholar] [CrossRef]
- Pandolfi, L.; Martino, R.; Belvedere, M.; Martínez-Navarro, B.; Medin, T.; Libsekal, Y.; Rook, L. The latest Early Pleistocene hippopotami from the human-bearing locality of Buia (Eritrea). Quat. Sci. Rev. 2023, 308, 108039. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martino, R.; Marra, F.; Ríos, M.; Pandolfi, L. The Middle Pleistocene Hippopotamus from Malagrotta (Latium, Italy): New Data and Future Perspectives. Quaternary 2024, 7, 13. https://doi.org/10.3390/quat7010013
Martino R, Marra F, Ríos M, Pandolfi L. The Middle Pleistocene Hippopotamus from Malagrotta (Latium, Italy): New Data and Future Perspectives. Quaternary. 2024; 7(1):13. https://doi.org/10.3390/quat7010013
Chicago/Turabian StyleMartino, Roberta, Fabrizio Marra, María Ríos, and Luca Pandolfi. 2024. "The Middle Pleistocene Hippopotamus from Malagrotta (Latium, Italy): New Data and Future Perspectives" Quaternary 7, no. 1: 13. https://doi.org/10.3390/quat7010013