
Neanderthal Hunting Weapons Re-Assessed: A Tip Cross-Sectional Area Analysis of Middle Palaeolithic Point Assemblages from South Eastern France

1
Palaeo-Research Institute, University of Johannesburg, Auckland Park, P.O. Box 524, Johannesburg 2006, South Africa
2
UMR 7194 CNRS-Museum National d’Histoire Naturelle, Institut de Paleontologie Humaine, 1 rue René Panhard, 75013 Paris, France
*
Author to whom correspondence should be addressed.
Quaternary 2023, 6(1), 17; https://doi.org/10.3390/quat6010017
Submission received: 29 January 2023
/
Revised: 10 February 2023
/
Accepted: 20 February 2023
/
Published: 1 March 2023
(This article belongs to the Special Issue Interdisciplinary Research into Cultural and Biological Transformations in the Paleolithic Period)
Abstract
:There are many opinions and arguments about the types of weapons that Neanderthals may have used. We list five assumptions about Neanderthal weapon-assisted hunting and suggest that the tip cross-sectional area (TCSA) approach may be used to assess these, and to provide a hypothetical overview of stone-tipped weaponry used in south eastern France between MIS 7 and MIS 3. We analysed stone points from Abri du Maras, Saint-Marcel, Grotte du Figuier, and Payre, and discuss possible hunting strategies in tandem with faunal evidence. Our results suggest that early Neanderthals may have hunted with bimanual thrusting spears in combination with one-handed stabbing spears, but that later groups possibly introduced javelin hunting to the Neanderthal arsenal. Stone-tipped assegais or one-handed stabbing spears, however, stand out as the Neanderthal weapons of choice throughout the Middle Palaeolithic, and we discuss the adaptive advantages of hunting with these weapons. Comparative results that include assemblages from elsewhere in the Old World suggest similarities in hunting with stone-tipped weapons between Neanderthal and H. sapiens before MIS 5, marked differences during MIS 5–4, but similarities again during MIS 3. We suggest that caution is needed when attributing MIS 3 point assemblages to Neanderthals based on age and/or technology only.
1. Introduction
Similar to Homo sapiens foraging groups, Neanderthals gathered plant foods and probably also collected other morsels such as eggs, snails, crabs and lizards—to name but a few [1,2,3,4]. Remains of such veld-foods seldom preserve in the archaeological record. Meat consumption, on the other hand, is well-documented in the many faunal assemblages, showing that Neanderthals hunted prey, scavenged from predators and probably harvested weakened, trapped or recently dead animals. They pursued and killed large prey such as Elephas with simple wooden spears [5], and herding angulates such as Equus with stone-tipped weapons [6,7]. The multi-functionality of Middle Palaeolithic convergent stone tools—and their possible disconnect with hunting technologies—remains a topic of debate [8,9]. This discourse may be a remnant of thinking about Neanderthals and other early human hunters as incapable of effective hunting or making and using composite (multi-part) weapons [10,11,12,13,14]. In much of the discussion there is also a skewed understanding about which methods are appropriate for generating micro-scale evidence of hunting, as opposed to those designed for setting up wide-ranging, testable hypotheses about ancient hunting strategies (see discussion in [15]).
The method we apply aims to demonstrate the ballistic suitability of the tools studied to form part of different types of hunting weapons. It cannot directly assess whether each of the artefacts were used as weapon tips, and some of the elements could have been used for other activities. Use trace evidence for Neanderthal hunting with stone-tipped weapons is, however, mounting [16,17], and together with archaeozoological and isotopic studies [18,19,20,21,22], we may accept that at least some convergent lithics were hafted to wooden shafts and used as weapon tips. Lazuén [16] suggested that hunting with stone-tipped weapons was pervasive throughout southern Europe as part of organised Middle Palaeolithic subsistence strategies. It would also seem that Neanderthals coordinated their hunting behaviours to exploit game that congregated in large herds [14]. Reconstructions of the palaeo-landscape and animal ethology by White et al. [23] show that Neanderthals did not necessarily pre-select and isolate individual animals from a herd to pursue and kill—in the way many current hunter-gatherers do. Instead, they ambushed whole herds, slaughtering at random amongst the animals. When it came to processing the carcasses for transport and consumption, however, they were highly selective. It thus appears that Neanderthals were “excellent tacticians, casual executioners and discerning diners” [23] (p. 10).
Mithen [24] (p. 198) wrote the following about Neanderthal hunting:
“It is widely thought that their main hunting weapons appear to have been short thrusting spears, tipped with stone points. The points themselves required considerable skill to make, but the weapons themselves remained quite simple in design. With such weapons, the Neanderthals appear to have frequently used a confrontational hunting technique. This seems to have been a major cause of the high frequency of bone fractures found on Neanderthal skeletal remains and the high mortality of young adults. Why Neanderthals did not invent more effective hunting weapons and methods to avoid such injuries remains unclear. We have no evidence for throwing spears, for bows and arrows, for spear-throwers, for pits, traps and snares”.
With this contribution, we analyse seven stone point assemblages from sites around the confluence of the Rhône and Ardèche-Payre tributaries on the south eastern margins of the Massif Central Mountains in France, (Figure 1). The assemblages include stone points from Abri du Maras (MIS 3 and 5), Saint-Marcel (MIS 3 and 5), Grotte du Figuier (MIS 3–4), and Payre (MIS 6 and 7) (Figure 1). We use the revised tip cross-section area (TCSA) method [15] as testable hypothesis-building tool to assess the following assumptions about Neanderthal hunting weapons:
2. Our Approach
Whereas the design of weapon components may vary in terms of style, weapon tips must articulate effectively with the physiological and cognitive constraints of the hunters as well as with their hunting ecology [30]. In addition, weapons must suit the chosen subsistence, personal risk management, and mobility strategies of the hunters who use them. Design aspects that impact their ballistic success or failure will therefore co-vary with the different selective forces [33,34,35,36,37]. The TCSA statistic is ballistically relevant, because: (a) it is key to the success of most hunting weapons by representing a tip’s ability to cut a hole through the hide of a prey animal [15,38]; and (b) it predicts the penetration depth of weapons—with smaller TCSA values reaching higher velocities and penetrating deeper than larger, heavier tips under similar launching conditions [39]. TCSA values (0.5 × maximum width × maximum thickness) cannot determine how any individual artefact was used. However, the method enables the assessment of large samples for best-fit ballistic probability over a range of weapon types [36,38,40], generating directly comparable quantitative data with little room for calculation or explanatory error [41].
The method, its interpretative potential, its shortcomings and improvements have been synthesised recently [15], so that we do not repeat that discussion here. Instead, we directly apply the calibrated TCSA values with statistical integrity for artefacts used to tip poisoned arrows, un-poisoned arrows, javelins and single-handed stabbing spears (Table 1) to our samples from south eastern France. The artefact sample setting the TCSA standard for bimanual-thrusting spears is still too small to be robust, for example, the TCSA range for four iron-tipped thrusting spears housed at the KwaZulu-Natal Museum is 180–360 mm2, and that of eight European Pleistocene wooden spears amounts to 408–878 mm2 [15]. For this study, we therefore use TCSA values of >200 mm2 as a working standard for bimanual thrusting spears.
With the introduction of a discrete TCSA category for javelin tips—a weapon system still in use by hunter-gatherers across the globe and therefore not to be dismissed in the context of deep-time hunting—the dart-tip category lost its statistical integrity [36,37]. What is more, the original dart-tip category does not consider the possibility of two distinct dart-tip TCSA values wherein large terrestrial spear-throwers were used for hunting sheep-sized and larger animals, and small Arctic spear-throwers to hunt marine mammals and birds [42,43]. We cannot resolve these issues here, and although the samples for other weapon-tip types have been augmented and their TCSA values calibrated [15], we still use the Thomas [44] and Shott [45] samples, and their TCSA range calculated by Shea [40], as representative of dart hunting. Using the scheme in Table 1, results in an almost complete overlap between the probability of dart use vs. the probability of javelin use. Thus, whilst relatively robust for hypothesising about the other weapon-delivery systems, TCSA results on their own are currently less able to highlight a distinction between javelin hunting and dart hunting. The most parsimonious approach to deal with this quandary is to accept the most conservative interpretation for overlapping TCSA values, namely javelin hunting, unless there is corroborating evidence for the use of spear-throwers.
We also assess hypothetical Neanderthal weapon-assisted hunting strategies on a wider scale. First, by comparing our results for south eastern France with those previously published for concurrent Neanderthal or probable/assumed Neanderthal assemblages and second, by looking at synchronous H. sapiens assemblages. We suggest that the groundwork laid in the resulting framework can serve to stimulate discussion and further research until data from many more assemblages are added to shape a robust framework for the evolution of weapon-assisted hunting during the Old World Pleistocene.
3. Our Assemblages
For the assemblages below we calculated the TCSA values for all the material with two convergent edges, regardless of the technical axis. Déjétée points and triangular flakes were also selected. The difference between a point (retouched or not) and a convergent scraper, for example, was made based on the value of the tip angle (with a graph to isolate groups if visible).
3.1. Payre
The two oldest assemblages in our study are from Payre, a collapsed cave site first excavated in the 1960s by Combier [46] and more recently by Moncel [47]. Today it is an open-air site of ≥80 m2 with a 5m-thick stratigraphic sequence with seven units (from bottom to top: G, F, E, D, C, B and A). Four of these units feature occupation layers (G, F, E and D). The site is known for providing Middle Palaeolithic layers with Neanderthal remains, bracketed between flowstone [48,49,50]. U-series on flowstone, ESR/U-series on teeth and thermoluminescence (TL) on flint suggest a chronology ranging from ~300 ka to ~140 ka [51,52]. Occupations at the base of the sequence (units G and F) are concurrent with the transition between MIS 8 and MIS 7, and the top (units E and D) with the end of MIS 6. Some of the ages obtained from the base flowstone are, however, younger or fall within the same range as the TL ages for overlying layer G [53], and these ages must be used with caution as minimum estimates. The archaeological units are attributed to the early Middle Palaeolithic [54,55,56]. We analysed the point assemblages from layer G (n = 152) associated with MIS 7, and layer D associated with MIS 6 (n = 141).
3.2. Saint-Marcel
This cave site is located on the left side of the Ardèche River, in Saint-Marcel d’Ardèche, not far from the Abri du Maras. Under the entrance, excavations by Gilles from 1974 to 1988 revealed a 6.50m-thick infill, corresponding to detrital cones built up as a result of the collapse of the entrance [57]. Stratigraphic and sedimentological studies identified about forty layers, grouped into two complexes (upper and lower). The upper complex (layers c to u) contains several recurrent Mousterian occupations showing no behavioural change despite sedimentary breaks, except for level u at the bottom. The uppermost part of the upper complex (layers c to f) belongs to the Late Middle Palaeolithic, deposited during a temperate and wet period (MIS 4/beginning of MIS 3). In layer f, red deer bones with cut marks produced dates of 37,850 ± 550 BP (OxA-19623), 37,850 ± 600 BP (OxA-19624) and 41,300 ± 1700 BP (OxA-19625), which together with sedimentological evidence, indicate a Late Mousterian occupation [58]. Levels k to u (from the bottom to the upper complex) correspond to MIS 5e-a according to biochronology [59]. We analysed points from layers i and j of the MIS 3 occupation (n = 25), and from layer u (n = 8) corresponding to MIS 5e.
3.3. Abri du Maras
Located in the Ardèche gorge, Abri du Maras is a rock shelter that was initially excavated by Gilles and Combier [46]. Since the 2000s, excavations have been conducted by M-H. Moncel. The new stratigraphy consists of layers numbered from 1 at the top to 6 at the bottom, with most archaeological material recovered from layers 5 and 4. Layer 4 is subdivided into levels 4.1 and 4.2, representing the two main occupation phases. Previous dating suggests a chronology ranging from MIS 5 to MIS 3 obtained from U-series ages of bones from the bottom of layer 4 and the top of layer 5 ranging from ~90 ka to ~70 ka [60]. Combined ESR/U-series dating yielded ages ranging from 90 ± 9 ka (layer 5) to 40 ± 3 ka (layer 4.1) [61]. The new ESR/U-series ages obtained for layer 4.2 are similar to those previously obtained for layers 4.1 and 4.2, ranging from 40 ± 3 to 55 ± 2 ka. Together with IRSL ages, they confirm the attribution of this part of the sequence to MIS 3, perhaps representing some of the latest Neanderthal occupations in the area [58,61]. We analysed the points from layers 4.1 and 4.2 as a single assemblage (n = 95) associated with MIS 3, and those from the underlaying layer 5 (n = 62) as associated with the MIS 5a-b occupation of the shelter.
3.4. Grotte du Figuier
The Figuier Cave with its three rooms is located on the bank of the Ardèche River. The 8m-high entrance is oriented to the south and opens into room one with a ~150 m2 surface area that was excavated by Chiron and Raymond during the end of the 19th and beginning of the 20th centuries. The Upper Palaeolithic at the site spans from the Aurignacian to the Magdalenian with Homo sapiens in rooms one and three, and Solutrean engravings [62]. The stratigraphy proposed by Combier [46] also includes two Middle Palaeolithic layers, one attributed to the Quina Mousterian, separated from the Upper Palaeolithic layers by a sterile sediment. More recent excavations were carried out in 2007–2008 by M-H. Moncel. Correlations between stratigraphic sequences in the three rooms allow for the identification of five layers on top of a sterile, micaceous sand base [62,63]. Previous dating of layer 4 (room one) to 52 ± 9 ka [61] suggests occupation during MIS 4–3. Two U-series ages were obtained for rooms two and three on a bone and a tooth of 34 ± 7 and 13 ± 1 ka, provide a minimum estimate only [53]. We analysed points from the room two Middle Palaeolithic levels (n = 21).
4. TCSA Results and Contextual Interpretation for the Assemblages from South Eastern France
4.1. Payre MIS 7
The oldest point assemblage, Payre MIS 7, has a median TCSA value of 197 mm2 (Table 2), indicating that if stone points were used to tip hunting weapons the trend was to produce tips that would be most effective as large single-handed stabbing spears or relatively small bimanual thrusting spears. Of all our samples, this assemblage shows the highest TCSA frequencies for stabbing spears (64.1%) and thrusting spears (44.7%) (Table 2). The frequencies for all other weapon-tip categories in the assemblage fall below the 15% threshold [41], so that such weapons were probably not part of stone-tipped hunting technologies if these were used during MIS 7 at Payre. Stabbing spears can also be thrown over short distances, and thrusting spears are ethno-historically associated with groups of hunters bringing down large and/or dangerous animals [15], both weapons are used during ambush hunting. We hypothesise that stabbing spears would have been effective for hunting the medium- and small-sized deer, whilst thrusting spears would have been useful for hunting the large (>300 kg) horses and aurochs. Using a combination of the two weapon-delivery types, perhaps with some members of the group using stabbing spears thrown over short distances (5–10 m) and others using thrusting spears for killing injured large animals at close quarters, would have had adaptive advantages for Neanderthal hunters—as opposed to using thrusting spears only.
For this assemblage it is, however, worth noting that the core technologies are not devoted to symmetrical point production. Instead, discoidal cores provided mostly elongated flakes and dejetée triangular flakes or points. Thus far, use trace results have provided evidence of mammal processing, but no conclusive evidence for convergent artefacts being hafted or used to tip hunting weapons [2,64]. Faunal data, however, indicate that during MIS 7 hunting or meat harvesting activities at Payre centred around wild horses (Equus ferus), red and roe deer (Cervus elaphus and Capreolus capreolus) and aurochs (Bos primigenius). The age profiles of the animals indicate that year-around hunting took place, but that horses were probably not hunted in summer and deer and aurochs probably not in winter [65]. If this was indeed the case, we may wonder what the people of Payre used as hunting weapons during MIS 7—if not stone-tipped spears, wooden spears are a possibility and may not have been preserved.
4.2. Payre MIS 6
During this phase, the TCSA median value for the point assemblage decreases relative to the MIS 7 phase. Now being 150 mm2, it is more consistent with a preferred production and use of tips most suited for single-handed stabbing spears. The TCSA frequency indicating the possible use of stabbing spears, however, decreases to 46.1% and for thrusting spears to 37.6% (Table 2). At the same time, the possibility of hunting with light-weight javelins becomes a minor possibility with a frequency of 17%, just over the 15% threshold for probable use. People still hunted wild horses and aurochs, but based on the animals’ age estimates it seems that hunting now mostly took place from summer to autumn, no longer during winter and spring [65]. Although the frequency is low, we may see the first tentative use of light-weight javelins by Neanderthal hunters in south eastern France, should they have used stone-tipped weaponry. Perhaps, being free from the constraints of heavy winter clothing and possible snowy or marshy winter-spring landscapes, running after wounded prey and throwing weapons over longer distances became increasingly attractive. For this point sample, corroborating use trace analyses are lacking.
4.3. Saint-Marcel MIS 5e
This sample is too small (n = 8) to draw any robust inferences from, but it continues the trend of probable point manufacture for one-handed stabbing spears with a TCSA median value of 146 mm2 (Table 1). Half of the points in this assemblage conform to ballistic expectations for use in this manner, 37.5% of the pieces could have functioned effectively as tips for bimanual thrusting spears, and the possible javelin-tip component rises to 25% (Table 2). Throughout the sequence, the faunal assemblage is dominated by cervids. In layer u (MIS 5), European fallow deer (Dama dama) are most prolific.
4.4. Abri du Maras MIS 5a-b
The median TCSA value of 108 mm2 is consistent with a focus on producing stone points most effective for use as tips for stabbing spears (Table 1). Compared to the Saint-Marcel sample, however, the decrease in probable thrusting-spear tip production (22.5%), and the increase in the production of tips suitable for javelins (32.3%), are more pronounced (Table 2). For the first time there is a possibility that tips suitable for spear-thrower darts may have been in the mix at 24.2%, but with an ~98% overlap with the javelin-tip category parsimony dictates that additional strands of evidence are required before dart hunting is inferred.
The MIS 5 faunal assemblage at Abri du Maras is dominated by medium sized reindeer (Rangifer tarandus) at >65%. Other species such as medium-sized red deer (Cervus elaphus), and large wild horse (Equus ferus) and steppe bison (Bison priscus) were also sometimes targeted during summer when Neanderthal groups used the shelter as a long-term, seasonal campsite [66]. Reindeer are alert, highly mobile and well dispersed so that they are fairly difficult to hunt. The most effective way to hunt them is to intercept migrations, kill in excess, and preserve the surplus meat and fat [67]. The specialisation in summer-time reindeer hunting may have stimulated Neanderthal groups to increase their use of javelins. Such a strategy would mean that they could initiate hunting from a greater distance to compensate for the alert mobility of reindeer herds. Once the herd became confused, and some animals wounded, hunters could come in for the kill at a shorter range with stabbing and thrusting spears to finish off their preferred targets. Detailed use trace analyses on this point assemblage have not been conducted, and corroborating evidence for dart hunting is absent. We therefore suggest that, based on the TCSA results, the most likely hunting scenario associated with this point assemblage is the use of stabbing spears, supported by both javelins and thrusting spears, to optimise reindeer ambush hunting.
4.5. Grotte du Figuier MIS 3–4
The median TCSA value of 108 mm2 for this assemblage is the same as for the Abri du Maras MIS 5a-b assemblage (Table 2). There is a clear preference for the knapping of points most suitable for tipping single-handed stabbing spears (57.1%), but, different from all the preceding assemblages, none of the points were suitable for tipping bimanual thrusting spears. Instead, 28.6% of the pieces would be most suited for javelin hunting when used to tip hunting weapons, with a 27.8% probability of dart hunting. Again, given the massive overlap in TCSA values for these two weapon systems, and lacking evidence of spear-throwers, the conservative interpretation is that whilst stabbing spears were the weapons of choice, the people who used Grotte du Figuier during the MIS 3–4 phase also hunted with lightweight javelins.
Bison and wild horses were still hunted, but now Alpine ibex (Capra ibex) were pursued almost as frequently as reindeer, and chamois (Rupicapra rupicapra) were also targeted by the people who used the site as a regular short-term camp [62,68]. Ibex herds are relatively small (≤20 animals) compared to horse or reindeer herds that may number hundreds of animals (swelling into the thousands during seasonal mass migrations), and the chamois is a small goat-antelope (<50 kg). Expanding hunting breadth to include these animals that may have been pursued individually rather than ambushed in herds, may have further stimulated the use of javelins as a hunting strategy.
4.6. Saint-Marcel MIS 3
The TCSA median value of 100 mm2 for this assemblage is also most consistent with points being knapped for tipping stabbing spears (Table 1)—should they have been intended for use in a composite hunting technology. Points best suited ballistically to tip stabbing spears comprise 48% of the assemblage (Table 2). Frequencies for potential javelin hunting and/or dart hunting is the same at (36%), the highest for their relative weapon-delivery types amongst all our assemblages. Medium-sized red deer is the most abundant prey animal, followed by roe deer, fallow deer (Dama dama), ibex (in the form of Capra caucasica, west Caucasian tur), extinct giant deer (Megaloceros giganteus), horse (Equus germanicus), aurochs, European ass (Equus hydruntinus) and wild boar (Sus scrofa) [69]. Seasonality indexes show that red deer were hunted all year round with most slaughters occurring during the autumn, mainly with young animals and adults killed in herds [68,70]. It seems that hunting with stone-tipped weapons from medium (javelin) to short distances (stabbing spears with short throwing capacity) was the preferred hunting strategy for this context, instead of hunting from closer quarters with stabbing and thrusting spears.
4.7. Abri du Maras MIS 3
For our last sample, the TCSA median value of 94 mm2 is also most consistent with tips used for hunting with stabbing spears, but now it is skewed towards the smaller range of these weapons (Table 1). The frequency for possible javelin tips is 30.5% and for possible dart tips 21% (Table 2). What stands out for this assemblage is the 20% possibility for bow hunting with un-poisoned stone-tipped arrows. The mammal assemblage is even more dominated by reindeer (88%), compared to the MIS 5a-b occupation of this site, with minor occurrences of horse, bison, giant deer, red deer and ibex, but now lagomorphs are also present [71]. Although a full-scale use trace analysis of all the points and point fragments has not been completed, some of the artefacts have impact traces and polishes consistent with their use as weapon tips [31]. Hardy et al. [31] interpreted the presence of twisted fibres on a few tools, together with their relatively small size, as possible evidence for string-thrown darts in a Neanderthal hunting context. Currently, there is no corroborating evidence for the use of either spear-throwers or bows at the site, and there are no definitive human remains, so that a direct association with Neanderthals remains elusive. Once again, we have to take the most cautious route, and interpret possible hunting with stone-tipped weapons at Abri du Maras during MIS 3 as a preference for the dual-use of stabbing spears and lightweight javelins. The presence of small game such as lagomorphs, and relatively good potential for both dart and arrow hunting, however, indicate a potentially interesting scenario for this context that requires further exploration.
5. The TCSA Results of Our Analysis in the Greater Context
Below we present the hypothetical reconstruction for the use of stone-tipped hunting weapons based on TCSA values. This broad interpretation represents only an initial framework to be continuously fleshed out, tested, strengthened and constrained with additional data and lines of evidence. We start with the earliest MIS stages working through time to MIS 3 and introduce the associated fossil hominins or assumed populations responsible for the production and use of the assemblages.
5.1. Marine Isotope Stage 7 and Older
Of the three assemblages directly associated with Homo neanderthalensis fossil remains, the Payre assemblage has the most pieces best suited for use as bimanual thrusting spears, but several tips would also have been suited for tipping single-handed stabbing spears (Figure 2). At Biache-Saint-Vaast in north-western France [72], it seems that Neanderthals were more geared towards hunting with stabbing spears only, with a similar trend represented at Tabun in Israel [40,41]. The older wooden spears from Schöningen in Germany and Clacton in England, presumably made by pre-Neanderthals, have relatively massive TCSA values that are most consistent with bimanual thrusting spears [15,41], even though they may also have been hurled over short distances [73,74].
Of the African assemblages, the TCSA value distribution for points from Kathu Pan in the Northern Cape of South Africa is most similar to that of Payre (Figure 2). Although this assemblage is not directly associated with hominin fossil material, and if we accept the association of the artefacts with the age estimate of >464 ka [76], it is reasonable to suggest that these artefacts were knapped by Homo heidelbergensis [77]. H. heidelbergensis fossils were recorded from Elandsfontein and Hoedjiespunt in South Africa during MIS 7 [78]. The Canteen Kopje assemblage from South Africa dating to ~300 ka [79] could have been made by either H. heidelbergensis or early H. sapiens. The latter is represented at Florisbad, South Africa, by ~259 ka [80,81]. Most of these pointed artefacts could serve well as tips for stabbing spears, with some also suitable for tipping bimanual thrusting weapons. The small-bodied, small-brained H. naledi may have shared the South African landscape with the larger humans between ~335–236 ka [82], but thus far there is no evidence that they hunted with stone-tipped weapons [83]. The Ethiopian assemblage from the Gademotta Formation—probably also made by either H. heidelbergensis or early H. sapiens—falls mostly in the TCSA range of stabbing spear tips.
5.2. Marine Isotope Stage 6
By MIS 6 the Pyre point assemblage produced by H. neanderthalensis still spans both stabbing and thrusting spears, but the median TCSA value is now more towards the mean for stabbing spears compared to being at the upper margin of this weapon-tip range during MIS 7 (Figure 3). Of all the MIS 6 assemblages Pyre, however, still has the highest frequency of artefacts with TCSA values conforming to the possible use of tipping bimanual thrusting spears. The other two sites associated with Neanderthal and probable Neanderthal occupation show a similar pattern. However, for Bouheben in France we used the descriptive statistics published in Villa and Lenoir [84] so that outliers and how they affect the mean/median values are invisible, and the Tabun sample (n = 12) is probably too small to draw any robust conclusions from.
For Africa, we currently have directly comparable data for four assemblages from South Africa only. The TCSA values for all of these indicate hunting with stabbing spears, but a simultaneous employment of bimanual thrusting spears, should the points have been used to tip hunting weapons. The use of thrusting spears seems especially frequent at the pan-scape sites of Rooidam at 36.2% and Florisbad at 42.1% [36,85,86], and less so at the river-ravine site of Olieboomspoort at 26.6% [36,87]. By this time, H. sapiens is accepted to be the only humans living in southern Africa, so that if TCSA values are indeed able to reveal something about weapon-assisted hunting behaviours, it would seem that H. neanderthalensis in the Levant and France and H. sapiens populations in southern Africa followed similar hunting strategies in terms of their choice and variation in hunting weapons. Interestingly, all the assemblages now have convergent pieces that would have been suited ballistically for javelin tips. Especially assemblages with >15% of its TCSA values falling in this category, such as Payre in France (17%) and Rooidam in South Africa (22.3%) may represent a new trend, wherein populations started to experiment with lighter spears that can be thrown at greater velocity over longer distances.
5.3. Marine Isotope Stage 5
MIS 5 is a climatically complex phase with oscillating interglacial (a, c, e) and glacial (b and d) substages, and the only assemblage from this phase directly associated with H. neanderthalensis remains is that of Tabun in Israel [40]. Its mean TCSA value is still within the stabbing spear range, but it is lower compared to the MIS 6 assemblage from the same site (Figure 4). The mean TCSA value for the small assemblage from Abrigo de la Quebrada in Spain [88,89] is the lowest of the assemblages probably associated with Neanderthal groups, whilst that of Saint-Marcel stands out as the one with the largest proportion falling in the thrusting spear-tip category. Interpretation of all three of these assemblages can only be thought of as preliminary, based on the small number of pointed artefacts recorded (Tabun n = 9, Abrigo de la Quebrada n = 6, Saint-Marcel n = 8). The Abri du Maras assemblage has a wide distribution, centred on making tips suitable for stabbing spears, but also includes pieces that would work well for tipping heavier thrusting weapons or lighter javelins (Figure 4).
The assemblage from Skhul B in Israel is directly associated with H. sapiens remains but has a roughly similar age estimate and TCSA value distribution as the Tabun assemblage, which is associated with a Neanderthal population at the time. All the other assemblages with available TCSA data come from southern Africa, showing complex variability in the use of possible stone-tipped weapon choices across the region. This variability and its possible implications have been discussed previously [36,41,43], and we do not repeat those details here.
5.4. Marine Isotope Stage 4
The Grotte du Figuier chamber two assemblage spans MIS 3–4. Here we first compare it to other assemblages with MIS 3–4 age estimates and those from MIS 4 contexts, before doing the same with MIS 3 below. For the first time, we see an assemblage in south eastern France assumed to have a Neanderthal context with no pointed artefacts suited hypothetically for hunting with bimanual thrusting spears (Figure 5). This is in contrast with the H. neanderthalensis associated assemblage from Kebara in Israel that has several artefacts suited for such hunting [40,41]. At Rosh Ein Mor, for which the human population remains unknown, there is a proportion of points most suitable for thrusting spears. In all three of these assemblages some of the points would also be well-suited for use as javelin tips based on their TCSA values (Figure 5).
It was during the MIS 4 phase that, based on the current assemblages with directly comparable data, the TCSA values indicate a pronounced difference between point assemblages manufactured and used by Neanderthal or assumed Neanderthal populations in the Levant and south eastern France, and those made and used by H. sapiens in southern Africa (Figure 5). TCSA values for the oldest southern African assemblage from White Paintings Shelter in Botswana has a median TCSA value that falls just within the upper limits for javelin tips, and a mean TCSA value at the lower limit for hunting with stabbing spears. Most of the subsequent assemblages distributed across the region have median TCSA values falling within the arrow range with some pieces also well-suited for tipping javelins. MIS 4 in southern Africa therefore represents the earliest known phase during which H. sapiens groups may have been hunting habitually with bows and arrows—an inference supported by multiple strands of evidence from independent studies and approaches [90,91,92,93,94]. If the artefacts measured were used to tip hunting weapons, amongst other things, these groups did not only hunt with bows and arrows, but also with javelins as suggested by the spread of the TCSA values and some outliers.
By the transition from the glacial MIS 4 to the interglacial MIS 3, however, the TCSA values for the youngest assemblage from southern Africa (Sibudu Cave dating to ~55–59 ka) indicate a return to hunting mostly with stabbing spears (Figure 5). Noteworthy here is also a return to discoidal and Levallois flaking at long-sequence Middle Stone Age sites such as Sibudu Cave during this phase [95]. A trend that already started by the end of the previous phase (Sibudu assemblage dated to ~62–65 ka) that was strongly blade based with backed implements [96]. At Sibudu Cave faunal data too indicate a gradual shift from hunting smaller forest/woodland prey to large, herding plains game such as zebra [97].
5.5. Marine Isotope Stage 3
We include here again the assemblages that span MIS 3–4. Similar to the previous inter-glacial (MIS 5), the TCSA values for the MIS 3 assemblages show variability around the use of spear hunting (Figure 6). The bona fide Neanderthal artefacts from Le Moustier in central-western France stand out as the only MIS 3 assemblage with TCSA values almost exclusively associated with bimanual thrusting spears. This assemblage is curated at the Harvard Peabody Museum, and we do not know how it was selected, so that future analyses on a more comprehensive sample may provide different results. The other two assemblages directly associated with H. neanderthalensis remains, namely Bagratashen 1 and Kebara, both show TCSA values most consistent with stabbing-spear hunting in tandem with the possible use of javelins (Figure 6).
Most assemblages assumed to have been made and used by Neanderthals have sizable TCSA frequencies falling within the hypothetical range of javelin tips. For example, assemblages from the Spanish sites of Amalda and Abrigo de la Quebrada [89,98,99] have mean and median TCSA values that fall within this category, whilst that from Boker Tachtit in Israel [100] is just within the range of stabbing spears with a large proportion of tips also suitable for hunting with javelins. The assemblages from Mirak in Iran [101] and Tor Faraj in Jordan [102] both have mean TCSA values most consistent with hunting with one-handed stabbing spears. The assemblage from Abri du Maras stands out amongst the assumed Neanderthal assemblages with its TCSA range also spanning that of arrows and darts, and those from Rosh Ein Mor and Mirak roughly follow the Neanderthal and assumed Neanderthal trend (Figure 6).
Amongst the assemblages associated or assumed to be associated with H. sapiens, that of Üçagizli in Turkey and Ksar Akil in Lebanon [40] stand out with median TCSA values within the arrow range—similar to Abri du Maras (Figure 6). The French Chatelperonian assemblage that consists of points from the sites of Trou de La Chevre, La Ferrassie and Grotte du Portel points [40] and Rose Cottage Cave in South Africa [36], both have median and mean TCSA values consistent with a preference for javelin hunting. For the remaining South African assemblages, namely that of Umbeli Belli, Umhlatuzana and Sibudu Cave, the marked differences in TCSA ranges between H. sapiens and Neanderthal assemblages observed for MIS 4 are no longer visible in the MIS 3 record. It seems that if stone tips were used to tip hunting weapons during this phase, people in southern Africa preferred to hunt with stabbing spears, often tipped with points produced with Levallois-like knapping methods similar to some of the assumed Neanderthal groups.
Figure 6.
Comparative analysis of TCSA values for assemblages dating to MIS 3. The hominin abbreviation key is the same as for Figure 5. The key to boxplot information is the same as for Figure 2. Data sources: Bagratashen [103]; Kebara [40,41]; Amalda [98,99]; Abrigo de la Quebrada [89]; Boker Tachtit [100]; Tor Faraj [102]; Mirak [101]; Üçagizli, Chatelperonian, Ksar Akil [40]; South African assemblages [36].
Figure 6.
Comparative analysis of TCSA values for assemblages dating to MIS 3. The hominin abbreviation key is the same as for Figure 5. The key to boxplot information is the same as for Figure 2. Data sources: Bagratashen [103]; Kebara [40,41]; Amalda [98,99]; Abrigo de la Quebrada [89]; Boker Tachtit [100]; Tor Faraj [102]; Mirak [101]; Üçagizli, Chatelperonian, Ksar Akil [40]; South African assemblages [36].

6. Discussion
In our introduction we listed five assumptions about Neanderthal weapon-assisted hunting that the TCSA approach may help re-assess. Below we discuss each of these topics based on the results of our study presented above. Other approaches and additional lines of evidence may provide different outcomes or support our inferences.
6.1. Assumption 1: Neanderthal Stone-Tipped Weaponry Was Mostly Limited to Double-Handed Thrusting Spears
Berger and Trinkaus [25] (p. 841) suggested that if stone points were used as weapon tips in Neanderthal contexts, “the resultant spears would have been thick and heavy, usable only as thrusting spears and not as effective throwing projectiles”, and that the Neanderthal wooden spears were best suited for “close quarter predation on ungulates”. They link the use of thrusting spears with morphological evidence for infrequent throwing behaviour in Neanderthal populations despite the strength of their upper limbs, and with high frequencies of head and neck trauma suffered by Neanderthals [25]. Churchill [26] and Schmitt et al. [104] also see the humeral morphology of Neanderthals and early H. sapiens as consistent with the habitual and forceful use of bimanual thrusting spears.
Our TCSA work on bimanual thrusting spears of known use [15], and the TCSA values of the Middle Palaeolithic wooden spears (Figure 2) corroborate their inferences, showing that some pointed stone artefacts from Neanderthal assemblages and the wooden spears are best-suited ballistically for use as bimanual thrusting spears. In the assemblages from south eastern France, such tips are relatively frequent in the ≥MIS 7–5e assemblages, where they constitute the second-most represented weapon-tip type after one-handed stabbing spears (Table 2). They become less frequent in the younger Neanderthal or assumed Neanderthal assemblages. The current TCSA data therefore supports the notion that early Neanderthal groups may have hunted habitually with bimanual thrusting spears. However, it does not support a scenario wherein this was their exclusive way of hunting, especially after MIS 5e. Instead, our TCSA data indicate that strategies around stone-tipped weapon use amongst Neanderthal or assumed Neanderthal groups changed through time to also include other approaches to spear-hunting.
6.2. Assumption 2: Neanderthals Used Stone-Tipped Spears for Both Thrusting and Short-Distance Throwing
Hardy et al. [27] examined artifacts from two sites in the Crimea, namely Starosele dating to ~80–40 ka and Buran Kaya III with an age estimate of 37–32 ka, assuming that they were made and used by Neanderthals based on the ‘cultural material’ and contemporaneous H. neanderthalensis remains from elsewhere. They suggested the use of both thrusting and throwing in terms of spear use. In the context of a functional analysis of Mousterian and Micoquian assemblages from Sesselfelsgrotte in Germany dating to ~48–36 ka assumed to be of Neanderthal origin, Rots [28] also found proxy evidence for both thrusting and throwing spear use on pointed artefacts. She argues that the same hunting weapon may have been used for both purposes but highlights that the weight of Neanderthal spears would limit their effective throwing distance to well below 10 m [28]. In both their discussions no distinction is made between bimanual thrusting and single-handed stabbing spears. The mechanics and biomechanics of one-handed stabbing are fundamentally different from bimanual thrusting [105], and the two hunting strategies may be associated with different evolutionary trajectories in hunting behaviours [104], and/or with adaptive advantages [37,41]. It is therefore important to consider both weapon-assisted hunting strategies in the Neanderthal context.
Our TCSA analyses support the use of bimanual thrusting weapons and single-handed stabbing spears that can be thrown effectively over short (< 10 m) distances for Neanderthal groups. For example, the TCSA results of the point assemblages from south eastern France demonstrate that in all seven instances there were more artefacts suited ballistically for tipping single-handed stabbing spears than any other weapon-tip category (Table 2). In the three older assemblages (Payre MIS 7, Payer MIS 6, Saint-Marcel MIS 5e) this category was followed by spear tips suited for bimanual thrusting. The four younger assemblages (Abri du Maras MIS 5 a-b, Grotte du Figuier MIS 3–4, Saint-Marcel MIS 3, Abri du Maras MIS 3), however, follow a different pattern wherein tips best-suited for javelin hunting becomes the second-most represented category (Table 2). We suggest that this indicates a development in hunting strategies on the south eastern margins of the Massif Central Mountains in France from contact or close-quarter hunting to hunting from a relatively safer distance. This trend also becomes evident in other Neanderthal or assumed Neanderthal assemblages dating to MIS 3 (Figure 6), so that we may accept that the trend observed for south eastern France also played out elsewhere in Europe and the Levant.
It is necessary at this stage to reflect on the adaptive advantages of hunting with single-handed stabbing spears or assegais. Before the recent introduction of firearms to Africa, various types of assegais represented the bulk of hunting and warfare weaponry across the continent because of the effectiveness and flexible nature of this weapon [15]. For example, in hunting scenarios single-handed stabbing spears or assegais:
- (a)
- Provide better balance and movement to weapon-yielding hunters compared to large and heavy bimanual thrusting spears;
- (b)
- Provide enough shaft and tip strength to stab powerfully and effectively at dangerous antagonists without hunters losing their weapons;
- (c)
- Can be flung from a short distance providing some flexibility in risk management;
- (d)
- Free up one hand to carry a backup or defence weapon such as a club or another spear or spear type.
Because each person can carry no more than one bimanual thrusting spear and no more than two or three stabbing spears, these weapons are unsuitable for hunting as a single person or in a pair. Instead, all spear-hunting strategies require multi-person cooperation [41]. The communal success of such social weapon-use strategies may explain why the stabbing-spear category became the preferred weapon for Neanderthal groups at least since MIS 7. Greater flexibility and development in Neanderthal or assumed Neanderthal stone-tipped weapon use may be indicated by the TCSA outcomes that suggest that stabbing spears may have been used in tandem with bimanual thrusting spears early on, and later together with lighter javelins that could be thrown over longer distances (Table 2). Bona fide TCSA indicators for habitual javelin hunting, however, only appear later in the Neanderthal record, and most often at sites without corroborating H. neanderthalensis fossil remains—a point we return to below (Figure 5 and Figure 6). This interpretation is supported by work that shows a shift in Middle Palaeolithic technical behaviour between the earlier (MIS 9–6) and the later (MIS 4–3) phases [54,55].
6.3. Assumption 3: Neanderthals Did Not Make or Use Mechanically Projected Technologies Such as the Spear-Thrower and Dart or the Bow and Arrow for Killing at a Distance
Based on neurological differences between H. neanderthalensis and H. sapiens, Coolidge et al. [29] argue that Neanderthals did not produce or use complex weaponry, such as harpoons launched with spear-throwers or bow and arrow sets [106,107]. According to them it suggests that Neanderthals may have been limited by an inability to aim long-distance weapons accurately, because of constraints involving hand/movement control, vision, and spatial awareness and judgment [108]. One of us have reached similar conclusions in work relating to a co-evolutionary approach to human cognition [30], wherein we built from Bruner et al. [109]. They explain the role of the precuneus in a process called ‘visuospatial integration’, which is integral to bimanual manipulation and operating over time and distance—as required for bow hunting—and which varies between H. neanderthalensis and H. sapiens [110,111,112,113].
Our TCSA analyses reveals that for the assemblages from south eastern France dating to MIS 5e and older, the frequencies of tips with values falling in the dart and arrow categories are all below the 15% benchmark of probable use. For the later assemblages the possibility of dart use with spear-throwers increases to 21–36% (Table 2). Yet, because of the overlap between the TCSA ranges for dart tips and javelin tips (Table 1), and because there is little to no definitive corroborating evidence for spear-thrower use in the Neanderthal record, we must follow a parsimonious interpretation wherein the data is most consistent with javelin hunting. The MIS 3 assemblage from Abri du Maras is the only one with artefacts that have TCSA values in the arrow range for >15%. Again, it is only during MIS 3 that other Neanderthal or assumed Neanderthal assemblages also show similarly low TCSA values, especially in the case of Kebara in Israel and Abrigo de la Quebrada in Spain (Figure 6). Here it is noteworthy that the Kebara assemblage is directly associated with H. neanderthalensis remains, but those from Abri du Maras and Abrigo de la Quebrada thus far lacks fossil hominin evidence. Thus, based on current evidence, it cannot be claimed that Neanderthals hunted with long-distance weapon systems such as the spear-thrower and dart or the bow and arrow—instead, the possibility of javelin hunting requires more exploration.
6.4. Assumption 4: Neanderthals Used String-Thrown Darts
Above we indicated that there is little to no corroborating evidence for spear-thrower use in the Neanderthal record. We have also presented Hardy et al.’s [31] interpretation that twisted fibres observed on a few small, pointed artefacts may be seen as proxy evidence for Neanderthals hunting with string-thrown darts at Abri du Maras during MIS 3. The Hardy study did not consider hunting with lightweight javelins, which would result in the production of relatively small weapon tips, similar in dimension to dart tips (Table 1). Thus, the string could have simply been used to tighten javelin tips in their shafts.
Our TCSA results show that even if darts were used in the manner described by Hardy et al. [31], the preferred stone-tipped hunting weapon used at the site during this phase was the trustworthy stabbing spear, possibly supported by javelin hunting (Table 2). Whilst there is currently no supporting evidence for the use of darts or arrows at Abri du Maras, or any of the other Neanderthal or assumed Neanderthal sites, seen together with the presence of small game such as lagomorphs, the Abri du Maras assemblage can be considered a ‘smoking gun’ for the use of complex hunting technologies. However, the timing and the fact that there are no directly associated H. neanderthalensis remains, require a more nuanced consideration than simply attributing it to Neanderthals based on age or technology.
6.5. Assumption 5: Neanderthal Weapons Were Similar to Those Used by Broadly Contemporaneous Homo sapiens Hunters in Africa
Villa and Soriano [32] set out to show that “Neanderthals were not inferior hunters” and that they used hunting weapons similar to those used by their concomitant H. sapiens counterparts in South Africa. They argue that both Middle Palaeolithic (Neanderthal) and Middle Stone Age (H. sapiens) assemblages indicate the use of hand-delivered spears only—including backed pieces that dominate southern African MIS 4 assemblages. In contrast to their findings, work conducted by Africa-based researchers at the same time and after their study provides multi-stranded proxy evidence suggesting bow-hunting behaviour in southern Africa from perhaps as early as ~80 ka (see evidence listed in [15,90]; but see [32] for different view). Our comparative TCSA analyses support an inference of broadly similar weapon-assisted hunting behaviours for H. neanderthalensis in Europe and the Levant and probable H. heidelbergensis and early H. sapiens groups in Africa before and until roughly MIS 6 (Figure 2 and Figure 3). By MIS 6, both groups seem to start experimenting with lighter javelins, whilst still depending heavily on the adaptive advantages of the stabbing spear.
For MIS 5—the period between ~130 ka and 71 ka—the TCSA data do not support the assumption that Neanderthal weapon use in Eurasia was the same as that of H. sapiens in Africa. During this phase, African H. sapiens weapon-assisted hunting strategies became more variable (Figure 4), possibly in response to climatic oscillations between the interglacial and glacial substages. The variation may also reflect increasingly flexible trends in socio-technical behaviour amongst the H. sapiens groups [30,114]. The comparative TCSA results for the subsequent glacial MIS 4 demonstrate clear differences between most H. sapiens assemblages from southern Africa and the contemporaneous Neanderthal or assumed Neanderthal assemblages (Figure 5). During this phase it seems that the bow and arrow became the weapon of choice in southern Africa, supplemented with javelins and some stabbing spears [36].
By MIS 3 the TCSA results indicate that hunting with stone-tipped weapons became largely variable in both Neanderthal and H. sapiens populations (Figure 6). The evolutionary narrative of Middle Palaeolithic Neanderthal weapon use in Eurasia compared to Middle Stone Age H. sapiens weapon use in Africa can therefore not be reduced to being ‘similar’ across the board. In the TCSA record we see trends of similarity before MIS 5, divergence in hunting with stone-tipped weapons during MIS 5–4, seemingly converging again during MIS 3 when H. sapiens appear and stay on the European landscape.
7. Conclusions
The TCSA method cannot determine the function of each artefact measured. However, if we accept that some convergent stone tools were used to tip hunting weapons, it becomes a powerful, standardised tool for generating directly comparable data about variability in weapon-assisted hunting strategies through time and across space. Because the ballistic characteristics of weapon tips (represented by their TCSA values) articulate directly with the contextual effectiveness of weapon-delivery systems, they can reveal aspects of adaptive success. Depending on the level of analysis and interpretation, comparative TCSA data can be used to generate hypotheses about intra- and inter-site weapon use on a regional or a global scale. With this contribution we reassessed some interpretations and assumptions about Neanderthal hunting with stone-tipped weaponry. We showed that in south eastern France, between MIS 7 starting by ~243 ka and MIS 3 ending by ~29 ka, the TCSA values of stone point assemblages indicate interesting developments that require future testing. For example:
- By using TCSA standards that distinguish between three spear-hunting strategies, we can argue that early Neanderthals probably hunted with both bimanual thrusting spears and single-handed stabbing spears.
- Collectively, the TCSA and faunal records of Payre suggest that its early Neanderthal population probably adapted their spear-use strategies (relying more or less on bimanual thrusting and single-handed stabbing) according to prey type and hunting season.
- The adaptive advantages of using smaller stabbing spears, and the fact that they could be thrown more effectively over short distances, seems to have encouraged experimentation with lighter javelin-type weapons, perhaps starting as early as MIS 6, but becoming increasingly pronounced in younger populations.
Especially during MIS 3 (~57–29 ka) the TCSA record for assemblages assumed to have been made and used by Neanderthals show a tendency towards hunting with lightweight javelins or even darts and/or arrows in combination with single-handed stabbing spears. Most of these assemblages are not directly associated with H. neanderthalensis fossil remains. One of us [41], has argued that for humans to throw lightweight javelins over longer distances with the necessary force and accuracy to become effective hunting weapons, several factors needed to converge. For example, a powerful thumb-tip to finger-tip prehension, the ability to throw objects from a distance at high speed and with great accuracy, and neurocognitive underpinnings associated with the ability to mentally map force transmission through time and across space [112]. The latter requires effective visuo-spatial integration associated with brain areas such as the precuneus that expanded to its full capacity only in H. sapiens by ~100 ka or during MIS 5 [115], when we see TCSA data showing an increase in possible javelin use in southern Africa associated with a H. sapiens population only.
Successful hand-eye coordination, spatial awareness and distance judgment probably developed incrementally throughout our genus, and the evolution of increasingly efficient visuo-spatial feedback was necessary for developing and choosing between the use of different weapon-delivery systems. Our TCSA data suggest that both H. neanderthalensis and H. sapiens started to independently develop a capacity for hunting at a distance. Yet, whereas javelin and bow hunting (supported by the use of stabbing spears) may have become habitual for H. sapiens during MIS 5–4 in the southern African record, these strategies seem not to have evolved to the same extent in the concurrent Neanderthal populations. It is therefore possible that the variation in weapon choice between the two populations reflect differences in neurocognitive development between ~130 ka and 57 ka. TCSA data for MIS 3, however, indicate a convergence in weapon choice between Neanderthal or assumed Neanderthal groups and H. sapiens groups.
The African TCSA record indicates that by MIS 3 H. sapiens hunters returned to the preferred use of stabbing spears in tandem with javelin hunting. This may reflect the opening of the landscape after the MIS 4 glacial and hunting larger animals in the context of group hunting, instead of focussing on smaller prey hunted alone, or with only a few hunters participating. African H. sapiens MIS 3 weapons were often tipped with stone points made with Levallois-like knapping techniques—similar to the weapon tips of contemporaneous Neanderthal populations further north.
Genetic research highlights multiple admixture events between H. neanderthalensis and H. sapiens populations, from as early as ~120 ka in the Near East [116], most likely in western Asia from ~60–50 ka [117], and in present-day Romania at ~42–37 ka [118,119]. Even if interaction or co-existence was limited to 1400–2900 years during the Châtelperronian (~43–40 ka) in France and northern Spain [120], it still spans 56–116 human generations at 25 years per generation. We do not know yet whether some of the H. sapiens groups who entered Eurasia from Africa also used Levallois knapping techniques. Picin et al. [121] unpack dilemmas and possibilities associated with so-called ‘transitional industries’ of the Near East and Europe containing Levallois elements, suggesting that it is perhaps not feasible to ascribe such assemblages to either Neanderthals or H. sapiens only. Socio-economical overlap and exchange, behavioural flexibility, as well as genetic mixing between the populations must be considered as an explanation for some of the aspects associated with the MIS 3 archaeological record in Eurasia. This is especially necessary when behavioural trends—such as hunting with long-distance weapons—are observed without the presence of decisive human remains.
Author Contributions
Conceptualization, M.L.; methodology, M.L.; validation, M-H.M; formal analysis, M.L.; investigation, M.L. and M.-H.M.; resources, M.L. and M.-H.M.; data curation M.-H.M.; writing—original draft preparation, M.L.; writing—review and editing, M.-H.M.; visualization, M.L.; project administration, M.-H.M.; funding acquisition, M.-H.M. All authors have read and agreed to the published version of the manuscript.
Funding
Fieldwork on the Middle Palaeolithic sites of the southeast margins of the Massif Central were awarded to M-H. Moncel by the “Service Régional de l’Archéologie, Auvergne-Rhône-Alpes”, French Ministry of Culture.
Data Availability Statement
Data for the French assemblages can be requested from M-H. Moncel. Comparative raw data for weapon tips of known use can be downloaded from https://link.springer.com/article/10.1007/s10816-022-09595-1 (accessed on 19 February 2023).
Acknowledgments
We thank Ran Barkai for inviting us to participate in this special volume, John Shea for sharing data with M.L., and four anonymous reviewers for their time and input that helped to improve our contribution. Any mistakes or omissions, however, remain our own.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hardy, B.L. Climatic variability and plant food distribution in Pleistocene Europe: Implications for Neanderthal diet and subsistence. Quat. Sci. Rev. 2010, 29, 662–679. [Google Scholar] [CrossRef]
- Hardy, B.L.; Moncel, M.H. Neanderthal use of fish, mammals, birds, starchy plants and wood 125–250,000 years ago. PLoS ONE 2011, 6, e23768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, A.G.; Brooks, A.S.; Piperno, D.R. Plant foods and the dietary ecology of Neanderthals and early modern humans. J. Hum. Evol. 2014, 69, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Nabais, M.; Dupont, C.; Zilhão, J. The exploitation of crabs by Last Interglacial Iberian Neanderthals: The evidence from Gruta da Figueira Brava (Portugal). Front. Environ. Archaeol. 2023, 2, 1097815. [Google Scholar] [CrossRef]
- Thieme, H.; Veil, S.; Meyer, W.; Moller, J.; Plisson, H. Neue Untersuchungen zum eemzeitlichen Elefanten-Jagdplatz Lehringen, Ldkr. Verden. Die Kunde 1985, 36, 11–58. [Google Scholar]
- Boëda, E.; Geneste, J.M.; Griggo, C.; Mercier, N.; Muhesen, S.; Reyss, J.L.; Taha, A.; Valladas, H. A Levallois point embedded in the vertebra of a wild ass (Equus africanus): Hafting, projectiles and Mousterian hunting weapons. Antiquity 1999, 73, 394–402. [Google Scholar] [CrossRef]
- Gaudzinski-Windheuser, S.; Kindler, L.; MacDonald, K.; Roebroeks, W. Hunting and processing of straight-tusked elephants 125,000 years ago: Implications for Neanderthal behavior. Sci. Adv. 2023, 9, eadd8186. [Google Scholar] [CrossRef] [PubMed]
- Lazuén, T. Please do not shoot the pianist. Criteria for recognizing ancient lithic weapon use. J. Archaeol. Sci. 2014, 46, 1–5. [Google Scholar] [CrossRef]
- Hovers, E.; Belfer-Cohen, A. Are lithics and fauna a match made in (prehistoric) heaven? J. Paleolit. Archaeol. 2020, 3, 108–125. [Google Scholar] [CrossRef]
- Klein, R.G. Archeology and the evolution of human behavior. Evol. Anthropol. 2000, 9, 17–36. [Google Scholar] [CrossRef]
- Stiner, M.C. Honor among Thievesa Zooarchaeological Study of Neandertal Ecology; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Shea, J.J. Spear points from the Middle Paleolithic of the Levant. J. Field Archaeol. 1988, 15, 441–450. [Google Scholar]
- Shea, J.J.; Brown, K.S.; Davis, Z.J. Controlled experiments with Middle Palaeolithic spear points: Levallois points. In Experimental Archaeology: Replicating Past Objects, Behaviors, and Processes; Mathieu, J.R., Ed.; BAR International Series: Oxford, UK, 2002; Volume 1035, pp. 55–72. [Google Scholar]
- Dusseldorp, G.L. A View to a Kill: Investigating Middle Palaeolithic Subsistence Using an Optimal Foraging Perspective; Sidestone Press: Leiden, The Netherlands, 2009. [Google Scholar]
- Lombard, M.; Lotter, M.G.; Caruana, M.V. The tip cross-sectional area (TCSA) method strengthened and constrained with ethno-historical material from sub-Saharan Africa. J. Archaeol. Method Theory 2022. [Google Scholar] [CrossRef]
- Lazuén, T. European Neanderthal stone hunting weapons reveal complex behaviour long before the appearance of modern humans. J. Archaeol. Sci. 2012, 39, 2304–2311. [Google Scholar] [CrossRef]
- Rots, V. Projectiles and hafting technology. In Multidisciplinary Approaches to the Study of Stone Age Weaponry; Iovita, R., Sano, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 167–185. [Google Scholar]
- Gaudzinski-Windheuser, S.; Noack, E.S.; Pop, E.; Herbst, C.; Pfleging, J.; Buchli, J.; Jacob, A.; Enzmann, F.; Kindler, L.; Iovita, R.; et al. Evidence for close-range hunting by last interglacial Neanderthals. Nat. Ecol. Evol. 2018, 2, 1087–1092. [Google Scholar] [CrossRef]
- Britton, K.; Grimes, V.; Niven, L.; Steele, T.E.; McPherron, S.; Soressi, M.; Kelly, T.E.; Jaubert, J.; Hublin, J.J.; Richards, M.P. Strontium isotope evidence for migration in late Pleistocene Rangifer: Implications for Neanderthal hunting strategies at the Middle Palaeolithic site of Jonzac, France. J. Hum. Evol. 2011, 61, 176–185. [Google Scholar] [CrossRef]
- Julien, M.A.; Rivals, F.; Serangeli, J.; Bocherens, H.; Conard, N.J. A new approach for deciphering between single and multiple accumulation events using intra-tooth isotopic variations: Application to the Middle Pleistocene bone bed of Schöningen 13 II-4. J. Hum. Evol. 2015, 89, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Kuitems, M.; van der Plicht, J.; Drucker, D.G.; Van Kolfschoten, T.; Palstra, S.W.; Bocherens, H. Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their paleoecological implications. J. Hum. Evol. 2015, 89, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Rivals, F.; Julien, M.A.; Kuitems, M.; Van Kolfschoten, T.; Serangeli, J.; Drucker, D.G.; Bocherens, H.; Conard, N.J. Investigation of equid paleodiet from Schöningen 13 II-4 through dental wear and isotopic analyses: Archaeological implications. J. Hum. Evol. 2015, 89, 129–137. [Google Scholar] [CrossRef] [PubMed]
- White, M.; Pettitt, P.; Schreve, D. Shoot first, ask questions later: Interpretative narratives of Neanderthal hunting. Quat. Sci. Rev. 2016, 140, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Mithen, S. The hunter-gatherer prehistory of human-animal interactions. Anthrozoös 1999, 12, 195–204. [Google Scholar] [CrossRef]
- Berger, T.D.; Trinkaus, E. Patterns of trauma among the Neandertals. J. Archaeol. Sci. 1995, 22, 841–852. [Google Scholar] [CrossRef]
- Churchill, S.E. Of assegais and bayonets: Reconstructing prehistoric spear use. Evol. Anthropol. 2002, 11, 185–186. [Google Scholar] [CrossRef]
- Hardy, B.L.; Kay, M.; Marks, A.E.; Monigal, K. Stone tool function at the paleolithic sites of Starosele and Buran Kaya III, Crimea: Behavioral implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10972–10977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rots, V. The functional analysis of the Mousterian and Micoquian assemblages of Sesselfelsgrotte, Germany: Aspects of tool use and hafting in the European Late Middle Palaeolithic. Quartär–Int. Jahrb. Zur Erforsch. Des Eiszeitalt. Und Der Steinzeit 2009, 56, 37–66. [Google Scholar]
- Coolidge, F.L.; Wynn, T.; Overmann, K.A.; Hicks, J.M. Cognitive archaeology and the cognitive sciences. In Human Paleoneurology; Bruner, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 177–208. [Google Scholar]
- Lombard, M.; Högberg, A. Four-field co-evolutionary model for human cognition: Variation in the Middle Stone Age/Middle Palaeolithic. J. Archaeol. Method Theory 2021, 28, 142–177. [Google Scholar] [CrossRef]
- Hardy, B.L.; Moncel, M.H.; Daujeard, C.; Fernandes, P.; Béarez, P.; Desclaux, E.; Navarro, M.G.C.; Puaud, S.; Gallotti, R. Impossible Neanderthals? Making string, throwing projectiles and catching small game during Marine Isotope Stage 4 (Abri du Maras, France). Quat. Sci. Rev. 2013, 82, 23–40. [Google Scholar] [CrossRef]
- Villa, P.; Soriano, S. Hunting weapons of Neanderthals and early modern humans in South Africa: Similarities and differences. J. Anthropol. Res. 2010, 66, 5–38. [Google Scholar] [CrossRef]
- Beck, C. Projectile point types as valid chronological units. In Unit Issues in Archaeology: Measuring Time, Space, and Material; Ramenofsky, A.F., Steffen, A., Eds.; University of Utah Press: Salt Lake City, UT, USA, 1998; pp. 21–40. [Google Scholar]
- Knecht, H. Projectile Technology; Plenum Press: New York, NY, USA, 1997. [Google Scholar]
- Yu, P.L. From atlatl to bow and arrow: Implicating projectile technology in changing systems of hunter-gatherer mobility. In Archaeology and Ethnoarchaeology of Mobility; Sellet, F., Greaves, R., Yu, P.L., Eds.; University Press Florida: Gainsville, FL, USA, 2006; pp. 201–220. [Google Scholar]
- Lombard, M. Variation in hunting weaponry for more than 300,000 years: A tip cross-sectional area study of Middle Stone Age points from southern Africa. Quat. Sci. Rev. 2021, 264, 107021. [Google Scholar] [CrossRef]
- Lombard, M. Re-considering the origins of Old World spearthrower-and-dart hunting. Quat. Sci. Rev. 2022, 293, 107677. [Google Scholar] [CrossRef]
- Hughes, S.S. Getting to the point: Evolutionary change in prehistoric weaponry. J. Archaeol. Method Theory 1998, 5, 345–408. [Google Scholar] [CrossRef]
- Sitton, J.; Story, B.; Buchanan, B.; Eren, M.I. Tip cross-sectional geometry predicts the penetration depth of stone-tipped projectiles. Sci. Rep. 2020, 10, 13289. [Google Scholar] [CrossRef]
- Shea, J.J. The origins of lithic projectile point technology: Evidence from Africa, the Levant, and Europe. J. Archaeol. Sci. 2006, 33, 823–846. [Google Scholar] [CrossRef]
- Lombard, M. A standardized approach to the origins of lightweight-javelin hunting. Lithic Technol. 2022, 1–11, in press. [Google Scholar] [CrossRef]
- Cattelain, P. Hunting during the Upper Paleolithic: Bow, spearthrower, or both? In Projectile Technology; Knecht, H., Ed.; Plenum Press: New York, NY, USA, 1997; pp. 213–240. [Google Scholar]
- Lombard, M.; Churchill, S. Revisiting Middle Stone Age hunting at≠GI, Botswana: A tip cross-sectional area study. S. Afr. Field Archaeol. 2022, 17, 11381. [Google Scholar] [CrossRef]
- Thomas, D.H. Arrowheads and atlatl darts: How the stones got the shaft. Am. Antiq. 1978, 43, 461–472. [Google Scholar] [CrossRef]
- Shott, M.J. Stones and shafts redux: The metric discrimination of chipped-stone dart and arrow points. Am. Antiq. 1997, 62, 86–101. [Google Scholar] [CrossRef]
- Combier, J. Le Paléolithique de l’Ardèche dans son Cadre Paléoclimatique, 4th ed.; l’Institut de Préhistoire de l’Université de Bordeaux: Bordeax, France, 1967. [Google Scholar]
- Moncel, M.-H. Des Occupations Humaines de la Moyenne Vallée du Rhône de la Fin du Pléistocène Moyen et du Début du Pléistocène Supérieur; Mémoire de la Société Préhistorique Française: Paris, France, 2008. [Google Scholar]
- Moncel, M.H.; Condemi, S. Des restes humains dans le site Paléolithique moyen ancien de Payre (Ardèche): Dents et pariétal. Bull. De La Société Préhistorique Française 1997, 94, 168–171. [Google Scholar] [CrossRef]
- Smith, T.M.; Austin, C.; Green, D.R.; Joannes-Boyau, R.; Bailey, S.; Dumitriu, D.; Fallon, S.; Grün, R.; James, H.F.; Moncel, M.-H.; et al. Wintertime stress, nursing, and lead exposure in Neanderthal children. Sci. Adv. 2018, 4, eaau9483. [Google Scholar] [CrossRef] [Green Version]
- Verna, C.; Détroit, F.; Kupczik, K.; Arnaud, J.; Balzeau, A.; Grimaud-Hervé, D.; Bertrand, S.; Riou, B.; Moncel, M.-H. The Middle Pleistocene hominin mandible from Payre (Ardèche, France). J. Hum. Evol. 2020, 144, 102775. [Google Scholar] [CrossRef]
- Masaoudi, H. Application des Méthodes du Déséquilibre des Familles de l’uranium (230Th/234U) et de la Résonance de Spin Éléctronique (ESR) à la Datation des Sites d’Orgnac 3, de Payre et de l’abri des Pêcheurs (Ardèche); Muséum National d’Histoire Naturelle: Paris, France, 1995. [Google Scholar]
- Valladas, H.; Mercier, N.; Ayliffe, L.K.; Falguères, C.; Bahain, J.J.; Dolo, J.M.; Froget, L.; Joron, J.L.; Masaoudi, H.; Reyss, J.L.; et al. Radiometric dates for the Middle Palaeolithic sequence of Payre (Ardèche, France). Quat. Geochron. 2008, 3, 377–389. [Google Scholar] [CrossRef] [Green Version]
- Richard, M.; Pons-Branchu, E.; Genuite, K.; Jaillet, S.; Joannes-Boyau, R.; Wang, N.; Genty, D.; Cheng, H.; Price, G.J.; Pierre, M.; et al. Timing of Neanderthal occupations in the southeastern margins of the Massif Central (France): A multi-method approach. Quat. Sci. Rev. 2021, 273, 107241. [Google Scholar] [CrossRef]
- Baena, J.; Moncel, M.-H.; Cuartero, F.; Chacón Navarro, M.G.; Rubio, D. Late Middle Pleistocene genesis of Neanderthal technology in Western Europe: The case of Payre site (south-east France). Quat. Int. 2017, 436, 212–238. [Google Scholar] [CrossRef]
- Carmignani, L.; Moncel, M.-H.; Fernandes, P.; Wilson, L. Technological variability during the Early Middle Palaeolithic in Western Europe: Reduction systems and predetermined products at the Bau de l’Aubesier and Payre (South-East France). PLoS ONE 2017, 12, e0178550. [Google Scholar] [CrossRef] [Green Version]
- Daffara, S.; Borel, A.; Moncel, M.-H. Conditioning of the raw materials on discoid exploitation strategies during the Early Middle Palaeolithic: The example of Payre level D (South-East France). Archaeol. Anthropol. Sci. 2019, 11, 4681–4695. [Google Scholar] [CrossRef]
- Debard, E. Le Quaternaire du Bas-Vivarais: Dynamique Sédimentaire, Paléoclimatologie et Chronologie d’après l’étude Sédimentologique des Remplissages d’avens, de Porches de Grottes et d’abris sous Roche: Comparaisons Avec le Velay Oriental; Université Claude Bernard: Lyon, France, 1988. [Google Scholar]
- Szmidt, C.C.; Moncel, M.-H.; Daujeard, C. New data on the Late Mousterian in Mediterranean France: First radiocarbon (AMS) dates at Saint-Marcel Cave (Ardèche). Comptes Rendus Palevol 2010, 9, 185–199. [Google Scholar] [CrossRef]
- Crégut-Bonnoure, E.; Boulbes, N.; Daujeard, C.; Fernandez, P.; Valensi, P. Nouvelles données sur la grande faune de l’Eémien dans le Sud-Est de la France. Quaternaire. Rev. De L’association Française Pour L’étude Du Quat. 2010, 21, 227–248. [Google Scholar] [CrossRef] [Green Version]
- Moncel, M.-H.; Michel, V. Première datation radiométrique par U-Th d’un niveau moustérien de l’Abri du Maras (Ardèche, France). Bull. De La Société Préhistorique Française 2000, 97, 371–375. [Google Scholar] [CrossRef]
- Richard, M.; Falguères, C.; Pons-Branchu, E.; Bahain, J.J.; Voinchet, P.; Lebon, M.; Valladas, H.; Dolo, J.M.; Puaud, S.; Rué, M.; et al. Contribution of ESR/U-series dating to the chronology of late Middle Palaeolithic sites in the middle Rhône valley, southeastern France. Quat. Geochron. 2015, 30, 529–534. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Puaud, S.; Daujeard, C.; Lartigot-Campin, A.-S.; Millet, J.-J.; Theodoropoulou, A.; Cregut-Bonnoure, É.; Gely, B.; Vercoutère, C.; Desclaux, E.; et al. La Grotte du Figuier (Saint-Martin-d’Ardèche): Bilan des travaux récents sur un site du Paléolithique moyen et supérieur de la moyenne vallée du Rhône (Sud-Est de la France). Bull. De La Société Préhistorique Française 2012, 109, 35–67. [Google Scholar]
- Moncel, M.-H.; Daujeard, C.; Cregut-Bonnoure, É.; Boulbes, N.; Puaud, S.; Debard, É.; Bailon, S.; Desclaux, E.; Escude, É.; Roger, T.; et al. Nouvelles données sur les occupations humaines du début du Pléistocène supérieur de la moyenne vallée du Rhône (France). Les sites de l’Abri des Pêcheurs, de la Baume Flandin, de l’Abri du Maras et de la Grotte du Figuier (Ardèche). Quaternaire 2010, 21, 385–411. [Google Scholar]
- Moncel, M.-H.; Chacón, M.G.; Coudenneau, A.; Fernandes, P. Points and convergent tools in the European Early Middle Palaeolithic site of Payre (SE, France). J. Archaeol. Sci. 2009, 36, 1892–1909. [Google Scholar] [CrossRef]
- Rivals, F.; Moncel, M.-H.; Patou-Mathis, M. Seasonality and intra-site variation of Neanderthal occupations in the Middle Palaeolithic locality of Payre (Ardèche, France) using dental wear analyses. J. Archaeol. Sci. 2009, 36, 1070–1078. [Google Scholar] [CrossRef]
- Marín, J.; Daujeard, C.; Saladié, P.; Rodríguez-Hidalgo, A.; Vettese, D.; Rivals, F.; Boulbes, N.; Crégut-Bonnoure, E.; Lateur, N.; Gallotti, R.; et al. Neanderthal faunal exploitation and settlement dynamics at the Abri du Maras, level 5 (south-eastern France). Quat. Sci. Rev. 2020, 243, 106472. [Google Scholar] [CrossRef]
- Geist, V. Of reindeer and man, modern and Neanderthal: A creation story founded on a historic perspective on how to conserve wildlife, woodland caribou in particular. Rangifer 2003, 23, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Daujeard, C.; Moncel, M.-H. On Neanderthal subsistence strategies and land use: A regional focus on the Rhone Valley area in southeastern France. J. Anthropol. Archaeol. 2010, 29, 368–391. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Daujeard, C. The variability of the Middle Palaeolithic on the right bank of the Middle Rhône Valley (southeast France): Technical traditions or functional choices? Quat. Int. 2012, 247, 103–124. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Daujeard, C.; Crégut-Bonnoure, E.; Fernandez, P.; Faure, M.; Guérin, C. L’occupation de la grotte de Saint-Marcel (Ardèche, France) au Paléolithique moyen: Stratégie d’exploitation de l’environnement et type d’occupation de la grotte. L’exemple des couches i, j et j’. Bull. De La Société Préhistorique Française 2004, 101, 257–304. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Chacón, M.G.; Vettese, D.; Courty, M.-A.; Daujeard, C.; Eixea, A.; Fernandes, P.; Allué, E.; Hardy, B.; Rivals, F.; et al. Late Neanderthals short-term and specialized occupations at the Abri du Maras (South-East France, level 4.1, MIS 3). Anthropol. Archaeol. Sci. 2021, 13, 45. [Google Scholar] [CrossRef]
- Rots, V. Insights into early Middle Palaeolithic tool use and hafting in Western Europe. The functional analysis of level IIa of the early Middle Palaeolithic site of Biache-Saint-Vaast (France). J. Archaeol. Sci. 2013, 40, 497–506. [Google Scholar] [CrossRef]
- Voormolen, B. Ancient Hunters, Modern Butchers: Schöningen 13II-4, a Kill-Butchery Site Dating from the Northwest European Lower Palaeolithic. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2008. [Google Scholar]
- Gaudzinski, S.; Roebroeks, W. Adults only: Reindeer hunting at the Middle Palaeolithic site Salzgitter Lebenstedt, northern Germany. J. Hum. Evol. 2000, 38, 497–521. [Google Scholar] [CrossRef]
- Sahle, Y.; Hutchings, W.K.; Braun, D.R.; Sealy, J.C.; Morgan, L.E.; Negash, A.; Atnafu, B. Earliest stone-tipped projectiles from the Ethiopian Rift date to> 279,000 years ago. PLoS ONE 2013, 8, e78092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porat, N.; Chazan, M.; Grün, R.; Aubert, M.; Eisenmann, V.; Horwitz, L.K. New radiometric ages for the Fauresmith industry from Kathu Pan, southern Africa: Implications for the Earlier to Middle Stone Age transition. J. Archaeol. Sci. 2010, 37, 269–283. [Google Scholar] [CrossRef]
- Wilkins, J.; Schoville, B.J.; Brown, K.S.; Chazan, M. Evidence for early hafted hunting technology. Science 2012, 338, 942–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusseldorp, G.; Lombard, M.; Wurz, S. Pleistocene Homo and the updated stone age sequence of South Africa. S. Afr. J. Sci. 2013, 109, 46–52. [Google Scholar] [CrossRef]
- Lotter, M.G.; Gibbon, R.J.; Kuman, K.; Leader, G.M.; Forssman, T.; Granger, D.E. A geoarchaeological study of the middle and upper Pleistocene levels at Canteen Kopje, Northern Cape Province, South Africa. Geoarchaeology 2016, 31, 304–323. [Google Scholar] [CrossRef]
- Grün, R.; Brink, J.S.; Spooner, N.A.; Taylor, L.; Stringer, C.B.; Franciscus, R.G.; Murray, A.S. Direct dating of Florisbad hominid. Nature 1996, 382, 500–501. [Google Scholar] [CrossRef]
- Bruner, E.; Lombard, M. The skull from Florisbad: A paleoneurological report. J. Anthropol. Sci. 2020, 98, 89–97. [Google Scholar]
- Dirks, P.H.; Roberts, E.M.; Hilbert-Wolf, H.; Kramers, J.D.; Hawks, J.; Dosseto, A.; Duval, M.; Elliott, M.; Evans, M.; Grün, R.; et al. The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. Elife 2017, 6, e24231. [Google Scholar] [CrossRef] [Green Version]
- Dusseldorp, G.L.; Lombard, M. Constraining the likely technological niches of late Middle Pleistocene hominins with Homo naledi as case study. J. Archaeol. Method Theory 2021, 28, 11–52. [Google Scholar] [CrossRef]
- Villa, P.; Lenoir, M. Hunting weapons of the Middle Stone Age and the Middle Palaeolithic: Spear points from Sibudu, Rose Cottage and Bouheben. S. Afr. Humanit. 2006, 18, 89–122. [Google Scholar]
- Kuman, K.A. Florisbad and ≠Gi: The Contribution of Open-Air Sites to Study of the Middle Stone Age in Southern Africa. Ph.D. Thesis, University of Pennsylvania, Philadelphia, PA, USA, 1989. [Google Scholar]
- Eltzholtz, A.K. Investigating Temporal Change in Fauresmith Technology: Insights from Rooidam 2, Northern Cape Province, South Africa. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2020. [Google Scholar]
- Val, A.; De la Peña, P.; Duval, M.; Bansal, S.; Colino, F.; Culey, J.; Hodgskiss, T.; Morrissey, P.; Murray, A.; Murungi, M.; et al. The place beyond the trees: Renewed excavations of the Middle Stone Age deposits at Olieboomspoort in the Waterberg Mountains of the South African Savanna Biome. Archaeol. Anthropol. Sci. 2021, 13, 116. [Google Scholar] [CrossRef]
- Real, C.; Eixea, A.; Sanchis, A.; Morales, J.V.; Klasen, N.; Zilhão, J.; Villaverde, V. Abrigo de la Quebrada level IV (Valencia, Spain): Interpreting a Middle Palaeolithic palimpsest from a zooarchaeological and lithic perspective. J. Paleolit. Archaeol. 2020, 3, 187–224. [Google Scholar] [CrossRef] [Green Version]
- Eixea, A.; Giner, B.; Jardón, P.; Zilhão, J.; Villaverde, V. Elementos líticos apuntados en el yacimiento de Paleolítico Medio del Abrigo de la Quebrada (Chelva, Valencia): Caracterización tecno-tipológica y análisis de las macrofracturas. Espacio, tiempo y forma. Prehist. Arqueol. 2015, 8, 78–109. [Google Scholar]
- Lombard, M.; Shea, J.J. Did Pleistocene Africans use the spearthrower-and-dart? Evol. Anthropol. 2021, 30, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lombard, M. Quartz-tipped arrows older than 60 ka: Further use-trace evidence from Sibudu, KwaZulu-Natal, South Africa. J. Archaeol. Sci. 2011, 38, 1918–1930. [Google Scholar] [CrossRef]
- Backwell, L.; Bradfield, J.; Carlson, K.J.; Jashashvili, T.; Wadley, L.; d’Errico, F. The antiquity of bow-and-arrow technology: Evidence from Middle Stone Age layers at Sibudu Cave. Antiquity 2018, 92, 289–303. [Google Scholar] [CrossRef]
- de la Peña, P.; Taipale, N.; Wadley, L.; Rots, V. A techno-functional perspective on quartz micro-notches in Sibudu’s Howiesons Poort indicates the use of barbs in hunting technology. J. Archaeol. Sci. 2018, 93, 166–195. [Google Scholar] [CrossRef]
- Bradfield, J.; Lombard, M.; Reynard, J.; Wurz, S. Further evidence for bow hunting and its implications more than 60,000 years ago: Results of a use-trace analysis of the bone point from Klasies River Main site, South Africa. Quat. Sci. Rev. 2020, 236, 106295. [Google Scholar] [CrossRef]
- Cochrane, G.W. The transition from Howieson’s Poort to post-Howieson’s Poort industries in southern Africa. S. Afr. Archaeol. Soc. Goodwin Ser. 2008, 10, 157–167. [Google Scholar]
- de la Peña, P.; Wadley, L. Technological variability at Sibudu Cave: The end of Howiesons Poort and reduced mobility strategies after 62,000 years ago. PLoS ONE 2017, 12, e0185845. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.L.; Plug, I. Animal exploitation strategies during the south African middle stone Age: Howiesons Poort and post-Howiesons Poort fauna from sibudu cave. J. Hum. Evol. 2008, 54, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Rios-Garaizar, J. Economic organization among Neanderthals: Amalda (Basque Country, Spain) level VII Mousterian. Zephyrus 2010, 65, 15–37. [Google Scholar]
- Sánchez-Romero, L.; Benito-Calvo, A.; Marín-Arroyo, A.B.; Agudo-Pérez, L.; Karampaglidis, T.; Rios-Garaizar, J. New insights for understanding spatial patterning and formation processes of the Neanderthal occupation in the Amalda I cave (Gipuzkoa, Spain). Sci. Rep. 2020, 10, 8733. [Google Scholar] [CrossRef] [PubMed]
- Yaroshevich, A.; Kaufman, D.; Marks, A. Weapons in transition: Reappraisal of the origin of complex projectiles in the Levant based on the Boker Tachtit stratigraphic sequence. J. Archaeol. Sci. 2021, 131, 105381. [Google Scholar] [CrossRef]
- Feizi, N.; Nasab, H.V. Symmetry and Middle Paleolithic Points: A Case Study of Iran’s Middle Paleolithic Points. Research Square 2022. Available online: https://assets.researchsquare.com/files/rs-257471/v3_covered.pdf?c=1647444337 (accessed on 26 December 2022).
- Groucutt, H.S. Middle Palaeolithic point technology, with a focus on the site of Tor Faraj (Jordan, MIS 3). Quat. Int. 2014, 350, 205–226. [Google Scholar] [CrossRef]
- Egeland, C.P.; Gasparian, B.; Fadem, C.M.; Nahapetyan, S.; Arakelyan, D.; Nicholson, C.M. Bagratashen 1, a stratified open-air Middle Paleolithic site in the Debed river valley of northeastern Armenia: A preliminary report. Archaeol Res. Asia 2016, 8, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, D.; Churchill, S.E.; Hylander, W.L. Experimental evidence concerning spear use in Neandertals and early modern humans. J. Archaeol. Sci. 2003, 30, 103–114. [Google Scholar] [CrossRef]
- Milks, A.; Champion, S.; Cowper, E.; Pope, M.; Carr, D. Early spears as thrusting weapons: Isolating force and impact velocities in human performance trials. J. Archaeol. Sci. Rep. 2016, 10, 191–203. [Google Scholar] [CrossRef]
- Wynn, T.; Coolidge, F. Why not cognition? Curr. Anthropol. 2008, 49, 895–897. [Google Scholar] [CrossRef]
- Wynn, T.; Coolidge, F.L. A Stone-Age Meeting of Minds: Neandertals became extinct while Homo sapiens prospered. A marked contrast in mental capacities may account for these different fates. Am. Sci. 2008, 96, 44–51. [Google Scholar] [CrossRef]
- Coolidge, F.L.; Overmann, K.A. Numerosity, abstraction, and the emergence of symbolic thinking. Curr Anthropol. 2012, 53, 204–225. [Google Scholar] [CrossRef] [Green Version]
- Bruner, E.; Spinapolice, E.; Burke, A.; Overmann, K.A. Visuospatial integration: Paleoanthropological and archaeological perspectives. In Evolution of Primate Social Cognition; Di Paolo, L., Di Vincenzo, F., De Petrillo, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 299–326. [Google Scholar]
- Bruner, E.; Lozano Ruiz, M. Extended mind and visuo-spatial integration: Three hands for the Neandertal lineage. J. Anthropol. Sci. 2014, 92, 273–280. [Google Scholar] [PubMed]
- Bruner, E.; Lozano Ruiz, M. Three hands: One year later. J. Anthropol. Sci. 2015, 93, 191–195. [Google Scholar]
- Gärdenfors, P.; Lombard, M. Causal cognition, force dynamics and early hunting technologies. Front. Psychol. 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Lombard, M. On the minds of bow hunters. In Squeezing Minds from Stones; Coolidge, F.L., Overmann, K.A., Wynn, T., Eds.; University of Oxford Press: Oxford, UK, 2019; pp. 473–496. [Google Scholar]
- Högberg, A.; Lombard, M. ‘I can do it’ becomes ‘we do it’: Kimberley (Australia) and Still Bay (South Africa) points through a socio-technical framework lens. J. Paleolit. Archaeol. 2020, 3, 633–663. [Google Scholar] [CrossRef] [Green Version]
- Bruner, E.; Preuss, T.M.; Chen, X.; Rilling, J.K. Evidence for expansion of the precuneus in human evolution. Brain Struct. Funct. 2017, 222, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Kuhlwilm, M.; Gronau, I.; Hubisz, M.J.; De Filippo, C.; Prado-Martinez, J.; Kircher, M.; Fu, Q.; Burbano, H.A.; Lalueza-Fox, C.; de La Rasilla, M.; et al. Ancient gene flow from early modern humans into Eastern Neanderthals. Nature 2016, 530, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Subiaul, F.; Galef, B.; Zentall, T.; Whiten, A. Social learning in humans and nonhuman animals: Theoretical and empirical dissections. J. Comp. Psychol. 2012, 126, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Hajdinjak, M.; Moldovan, O.; Constantin, S.; Mallick, S.; Skoglund, P.; Patterson, N.; Rohland, N.; Lazaridis, I.; Nickel, B.; et al. An early modern human from Romania with a recent Neanderthal ancestor. Nature 2015, 524, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Gokcumen, O. Archaic hominin introgression into modern human genomes. Am. J. Phys. Anthropol. 2019, 171, 60–73. [Google Scholar] [CrossRef]
- Djakovic, I.; Soressi, M.; Key, A. Optimal linear estimation models predict 1400-2800 years of co-existence between Neandertals and Homo sapiens in western Europe. bioRxiv 2022. [Google Scholar] [CrossRef]
- Picin, A.; Moroni, A.; Benazzi, S. The arrival of Homo sapiens in the Near East and Europe. In Updating Neanderthals; Romagnoli, F., Rivals, F., Benazzi, S., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2022; pp. 321–347. [Google Scholar]
Figure 1.
Location of the sites from which assemblages were analysed for this study, and points from the oldest Payre unit Ga, MIS 7 (a,b) and youngest Abri du Maras units 4.1 and 4.2, MIS 3 (c–e) assemblages in our study. Maps by M Lombard and artefact photographs by M-H. Moncel.
Figure 1.
Location of the sites from which assemblages were analysed for this study, and points from the oldest Payre unit Ga, MIS 7 (a,b) and youngest Abri du Maras units 4.1 and 4.2, MIS 3 (c–e) assemblages in our study. Maps by M Lombard and artefact photographs by M-H. Moncel.

Figure 2.
Comparative analysis of TCSA values for assemblages dating to MIS 7 and older. Key to assemblages: HN = assemblages with Homo neanderthalensis fossil material; prob HN = assemblages with no known hominin fossil material, but probably produced by H. neanderthalensis; prob HH = assemblages with no known hominin fossil material, but probably produced by H. Heidelbergensis; eHS = early/archaic H. sapiens. Key to boxplot information: lines in boxes = median TCSA values, x in boxes = mean TCSA values, circles in/outside boxes = TCSA values for each tool measured, the dashed black lines sub-divide assemblages for easier reading. Data sources: Biache-Saint-Vaast [72]; Tabun [40,41]; Schöningen and Clacton [41]; Gademotta [75]; Kathu Pan [36].
Figure 2.
Comparative analysis of TCSA values for assemblages dating to MIS 7 and older. Key to assemblages: HN = assemblages with Homo neanderthalensis fossil material; prob HN = assemblages with no known hominin fossil material, but probably produced by H. neanderthalensis; prob HH = assemblages with no known hominin fossil material, but probably produced by H. Heidelbergensis; eHS = early/archaic H. sapiens. Key to boxplot information: lines in boxes = median TCSA values, x in boxes = mean TCSA values, circles in/outside boxes = TCSA values for each tool measured, the dashed black lines sub-divide assemblages for easier reading. Data sources: Biache-Saint-Vaast [72]; Tabun [40,41]; Schöningen and Clacton [41]; Gademotta [75]; Kathu Pan [36].

Figure 3.
Comparative analysis of TCSA values for assemblages dating to MIS 6. Key: Same as for Figure 2, but HS = assemblages made by Homo sapiens, because these are the only known humans on the African landscape at the time. The key to boxplot information is the same as for Figure 2. Data sources: Bouheben [84]; Tabun [40,41]; South African assemblages [36].
Figure 3.
Comparative analysis of TCSA values for assemblages dating to MIS 6. Key: Same as for Figure 2, but HS = assemblages made by Homo sapiens, because these are the only known humans on the African landscape at the time. The key to boxplot information is the same as for Figure 2. Data sources: Bouheben [84]; Tabun [40,41]; South African assemblages [36].

Figure 4.
Comparative analysis of TCSA values for assemblages dating to MIS 5. Keyfor hominin abbreviations is the same as for Figure 3. The key to boxplot information is the same as for Figure 2. Data sources: Tabun [40]; Abrigo de la Quebrada [89]; Lehringen [41]; South African assemblages [36]; ≠Gi Pan [43].
Figure 4.
Comparative analysis of TCSA values for assemblages dating to MIS 5. Keyfor hominin abbreviations is the same as for Figure 3. The key to boxplot information is the same as for Figure 2. Data sources: Tabun [40]; Abrigo de la Quebrada [89]; Lehringen [41]; South African assemblages [36]; ≠Gi Pan [43].

Figure 5.
Comparative analysis of TCSA values for assemblages dating to MIS 4. Hominin abbreviation key: Same as for Figure 3, but ass HN = assemblages with no human fossil material but assumed to be associated with H. neanderthalensis based on technology or dating only, even though H. sapiens may have been on the landscape. The key to boxplot information is the same as for Figure 2. Data sources: Kebara [40,41]; Rosh Ein Mor [40]; Southern African assemblages [36].
Figure 5.
Comparative analysis of TCSA values for assemblages dating to MIS 4. Hominin abbreviation key: Same as for Figure 3, but ass HN = assemblages with no human fossil material but assumed to be associated with H. neanderthalensis based on technology or dating only, even though H. sapiens may have been on the landscape. The key to boxplot information is the same as for Figure 2. Data sources: Kebara [40,41]; Rosh Ein Mor [40]; Southern African assemblages [36].


{kind=link}
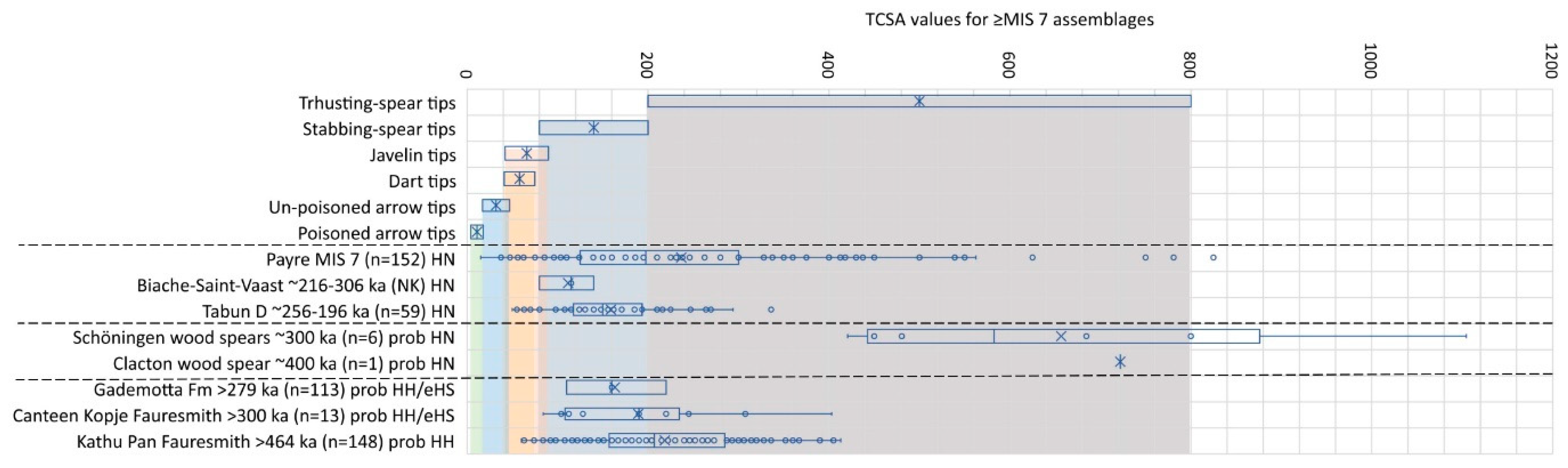{kind=link}
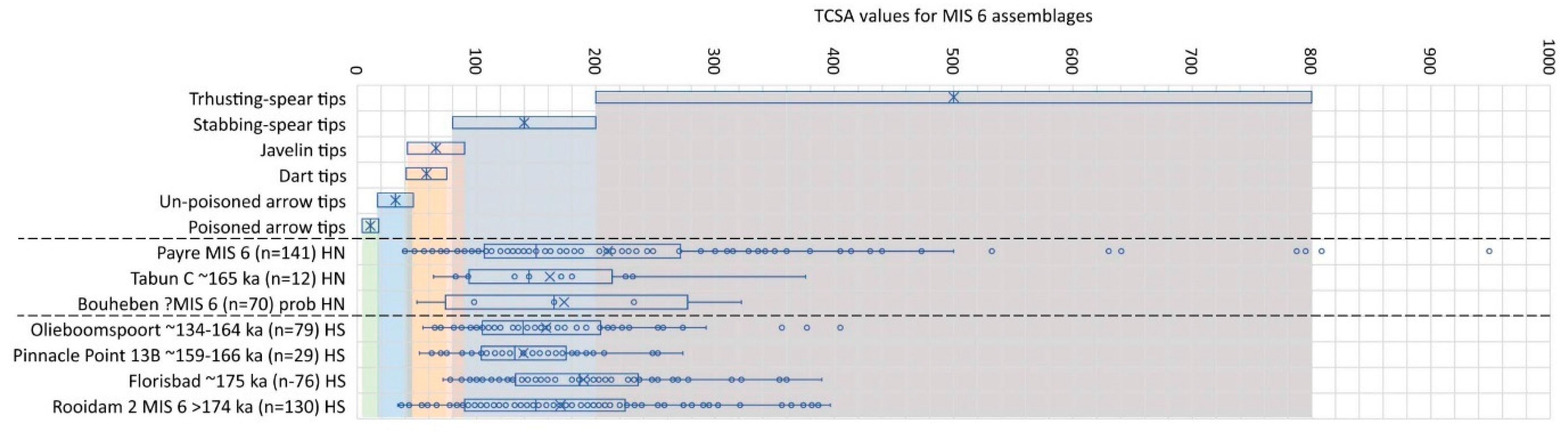{kind=link}
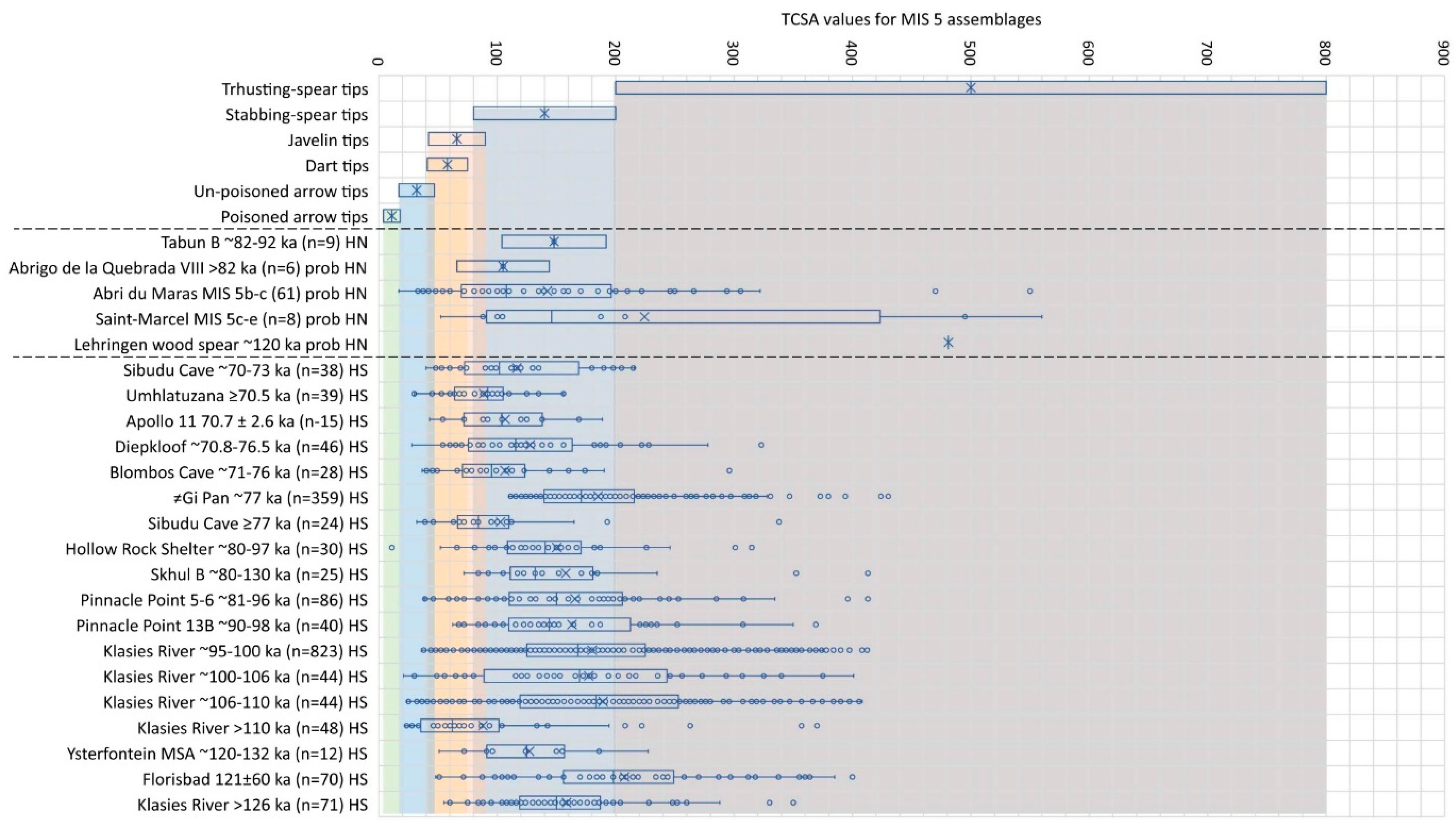{kind=link}
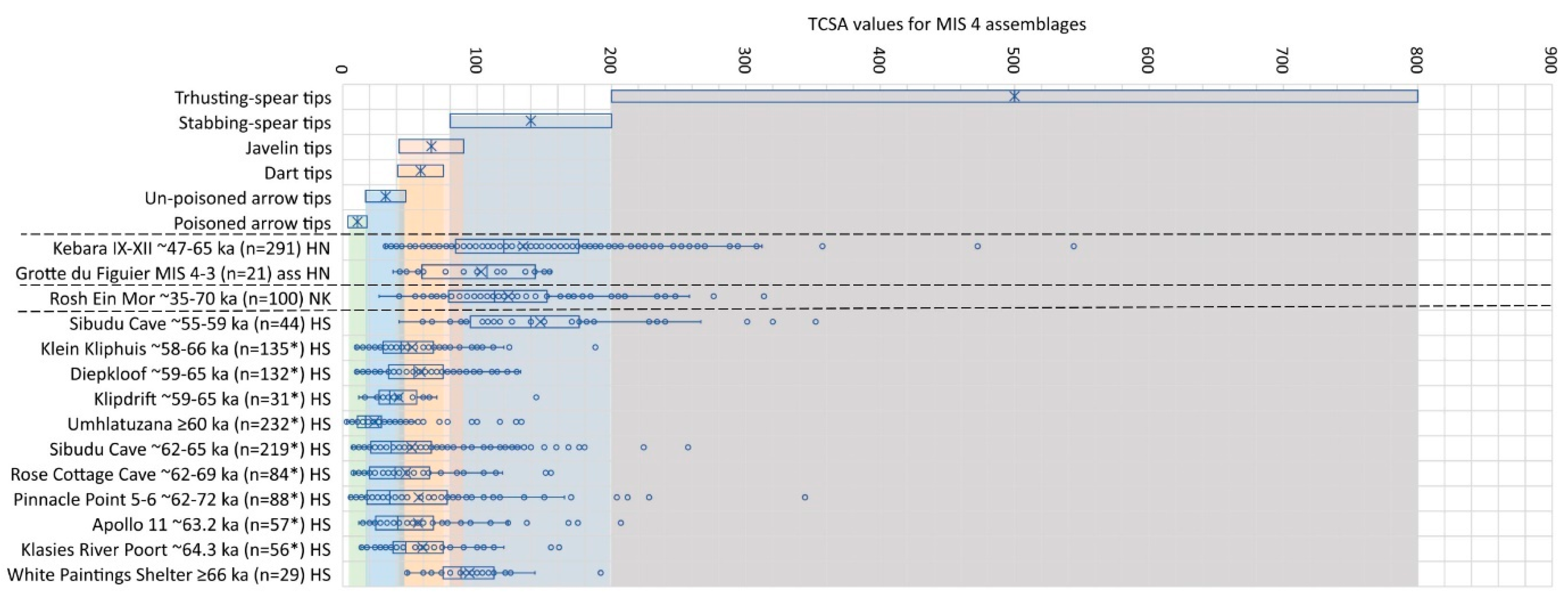{kind=link}
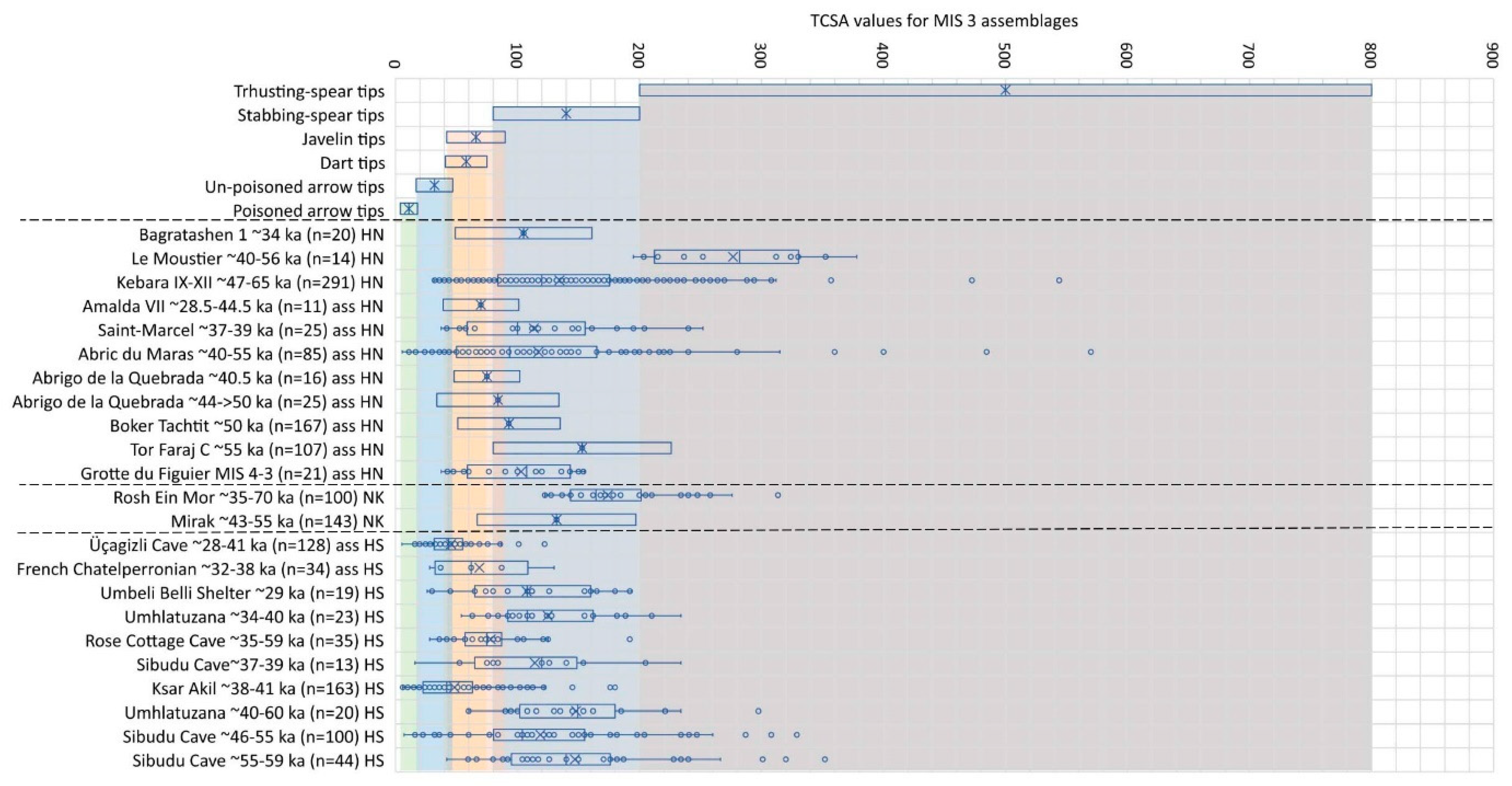{kind=link}
Table 1.
Newly calibrated TCSA standards for six weapon types used by recent hunter-gatherers [15].
Table 1.
Newly calibrated TCSA standards for six weapon types used by recent hunter-gatherers [15].
| TCSA Standard | N | Mean mm2 | SD mm2 | Range mm2 | Median mm2 |
|---|---|---|---|---|---|
| Poisoned arrow tips (tested/calibrated) | 565 | 11 | 7 | 4–18 | 8 |
| Un-poisoned arrow tips (tested/calibrated) | 338 | 32 | 15 | 17–47 | 31 |
| Dart tips (working standard) | 40 | 58 | 17 | 41–75 | 60 |
| Javelin tips (tested/calibrated) | 270 | 66 | 24 | 42–90 | 63 |
| Stabbing spear tips (tested/calibrated) | 141 | 140 | 60 | 80–200 | 128 |
| Thrusting spear tips (working standard) | 12 | 519 | 266 | >200 | 465 |
Table 2.
TCSA statistics (values in mm²) and percentage frequencies of potential weapon-delivery systems based on our analysis.
Table 2.
TCSA statistics (values in mm²) and percentage frequencies of potential weapon-delivery systems based on our analysis.
| Assemblage | TCSA Statistics in mm² | % Potential Weapon-Delivery System | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Median | Poisoned Arrow | Arrow | Dart | Javelin | Stabbing Spear | Thrusting Spear | |
| Abri du Maras, MIS 3 (n = 95) | 117 | 99 | 94 | 6.3 | 20 | 21 | 30.5 | 47.4 | 12.6 |
| Saint-Marcel, MIS 3 (n = 25) | 113 | 63 | 100 | 0 | 12 | 36 | 36 | 48 | 12 |
| Grotte du Figuier, MIS 3–4 (n = 21) | 103 | 42 | 108 | 0 | 9.5 | 27.8 | 28.6 | 57.1 | 0 |
| Abri du Maras, MIS 5a-b (n = 61) | 142 | 103 | 108 | 1.6 | 9.8 | 24.2 | 32.3 | 46.8 | 22.5 |
| Saint-Marcel, MIS 5e (n = 8) | 224 | 195 | 146 | 0 | 0 | 12.5 | 25 | 50 | 37.5 |
| Payre, MIS 6 (n = 141) | 210 | 165 | 150 | 0 | 1.4 | 0.7 | 17 | 46.1 | 37.6 |
| Payre MIS 7 (n = 152) | 237 | 152 | 197 | 0.7 | 1.3 | 7.2 | 9.9 | 64.1 | 44.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lombard, M.; Moncel, M.-H. Neanderthal Hunting Weapons Re-Assessed: A Tip Cross-Sectional Area Analysis of Middle Palaeolithic Point Assemblages from South Eastern France. Quaternary 2023, 6, 17. https://doi.org/10.3390/quat6010017
AMA Style
Lombard M, Moncel M-H. Neanderthal Hunting Weapons Re-Assessed: A Tip Cross-Sectional Area Analysis of Middle Palaeolithic Point Assemblages from South Eastern France. Quaternary. 2023; 6(1):17. https://doi.org/10.3390/quat6010017
Chicago/Turabian StyleLombard, Marlize, and Marie-Hélène Moncel. 2023. "Neanderthal Hunting Weapons Re-Assessed: A Tip Cross-Sectional Area Analysis of Middle Palaeolithic Point Assemblages from South Eastern France" Quaternary 6, no. 1: 17. https://doi.org/10.3390/quat6010017