
Landscape Response to Dynamic Human Pressure in the Paliouras Lagoon, Halkidiki Peninsula, Macedonia, Greece
 , , , and
, , , and
Abstract
:1. Introduction
1.1. Study Area, Vegetation and Climate, Human Occupation
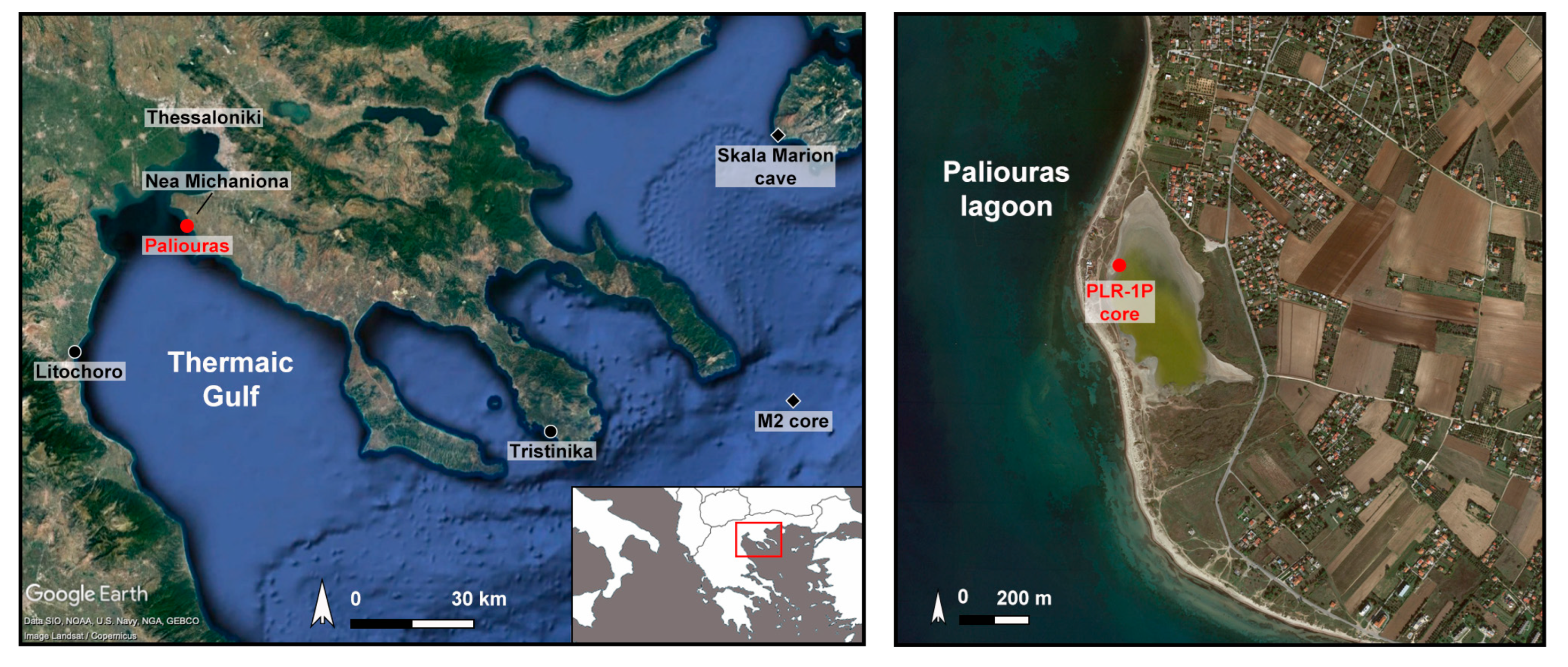
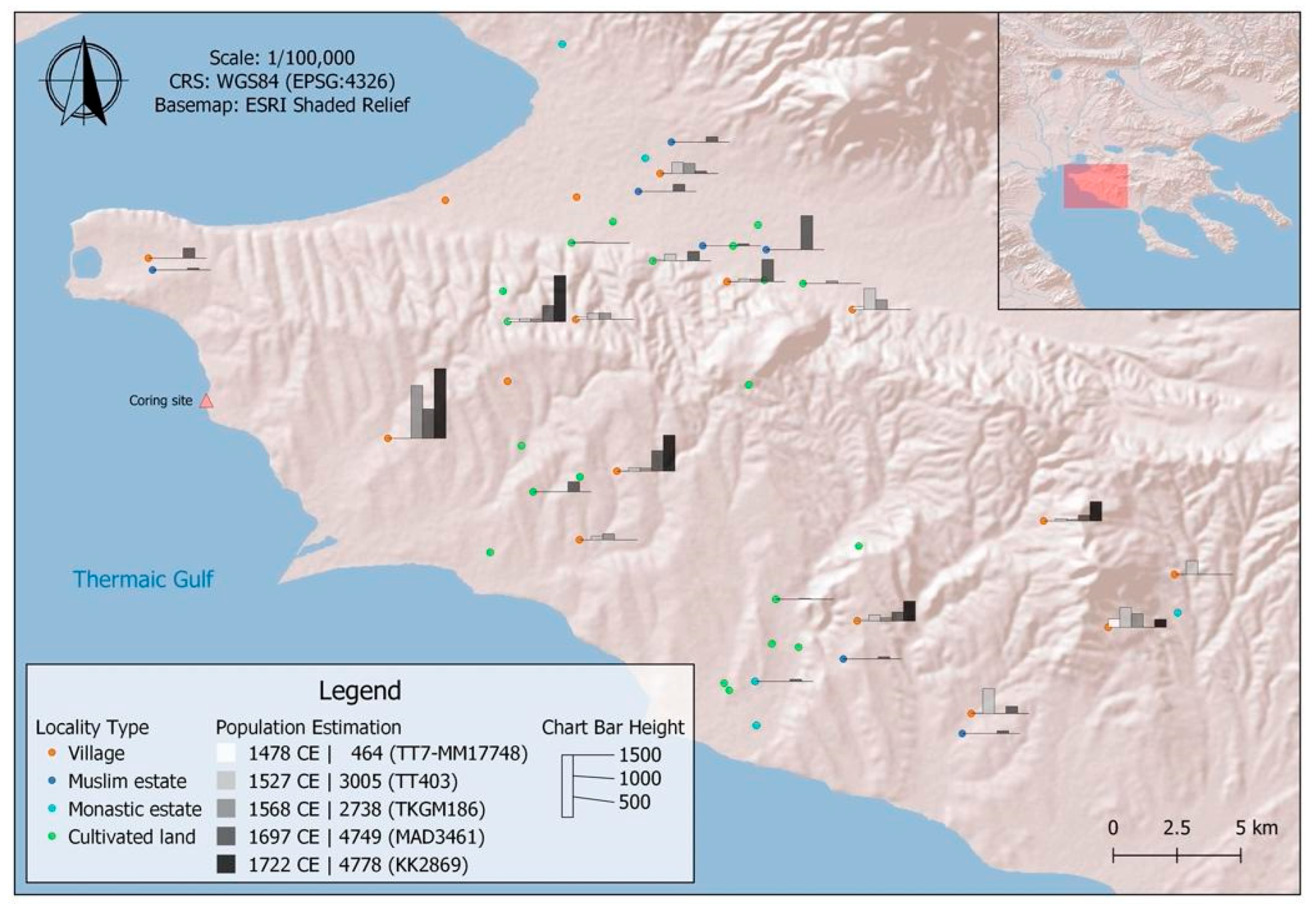
1.1.1. Site
1.1.2. Vegetation
1.1.3. Climate
1.1.4. Archaeological and Historical Setting
2. Materials and Methods
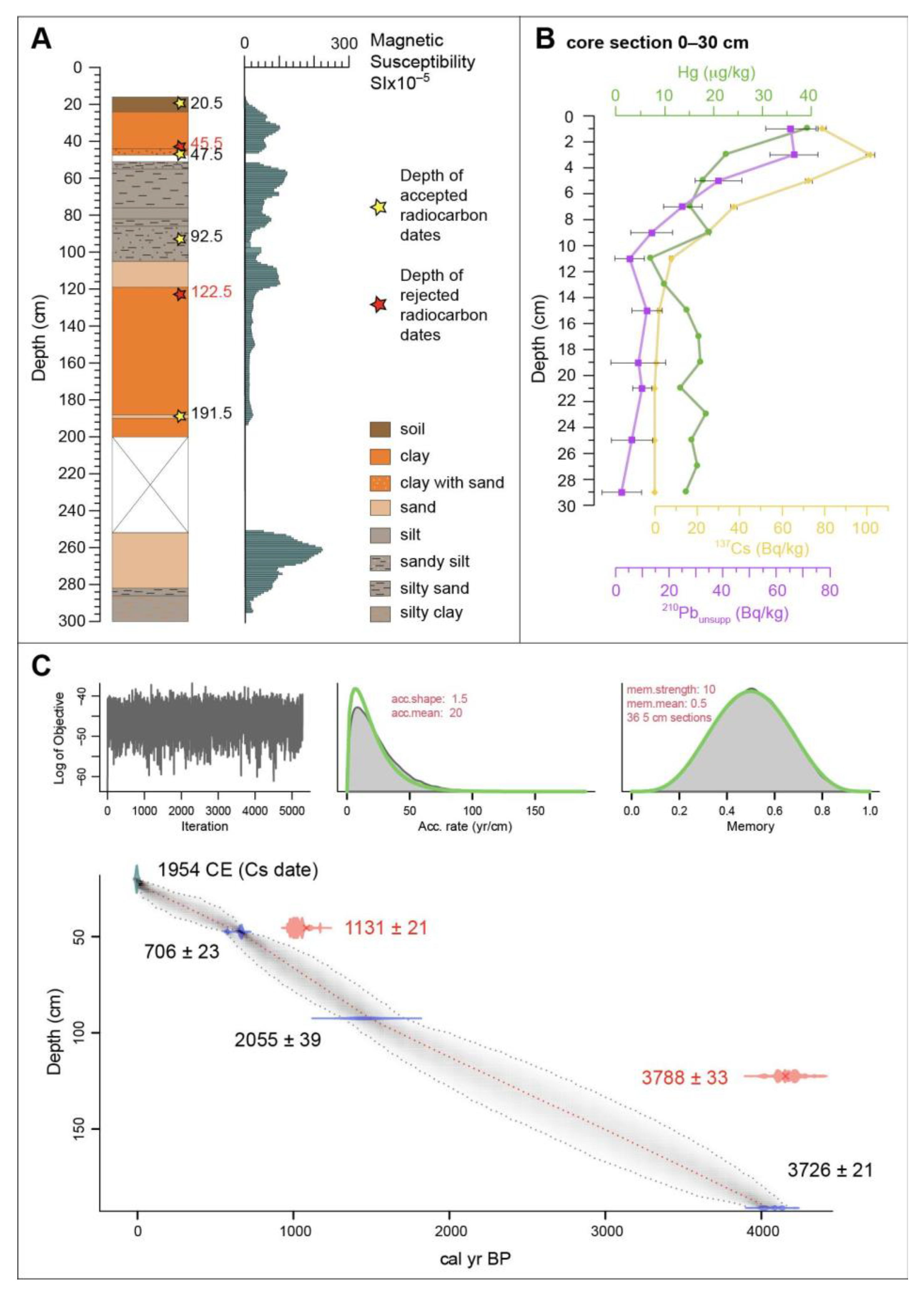
2.1. Lithology, Magnetic Susceptibility, and Mollusc Analysis
2.2. Age Depth Model

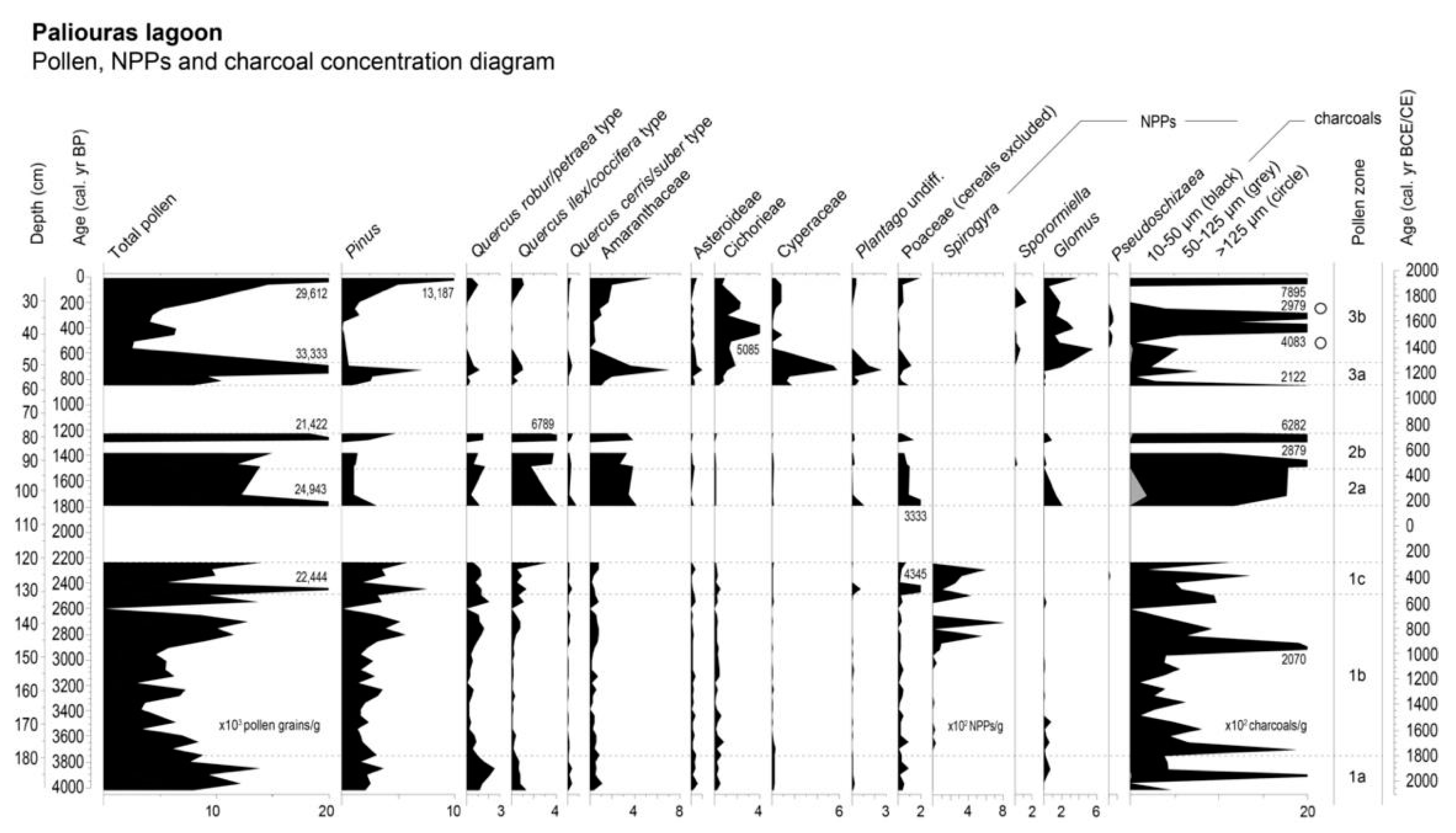
2.3. Pollen Analysis
3. Results
3.1. Lithology, Magnetic Susceptibility, and Molluscs
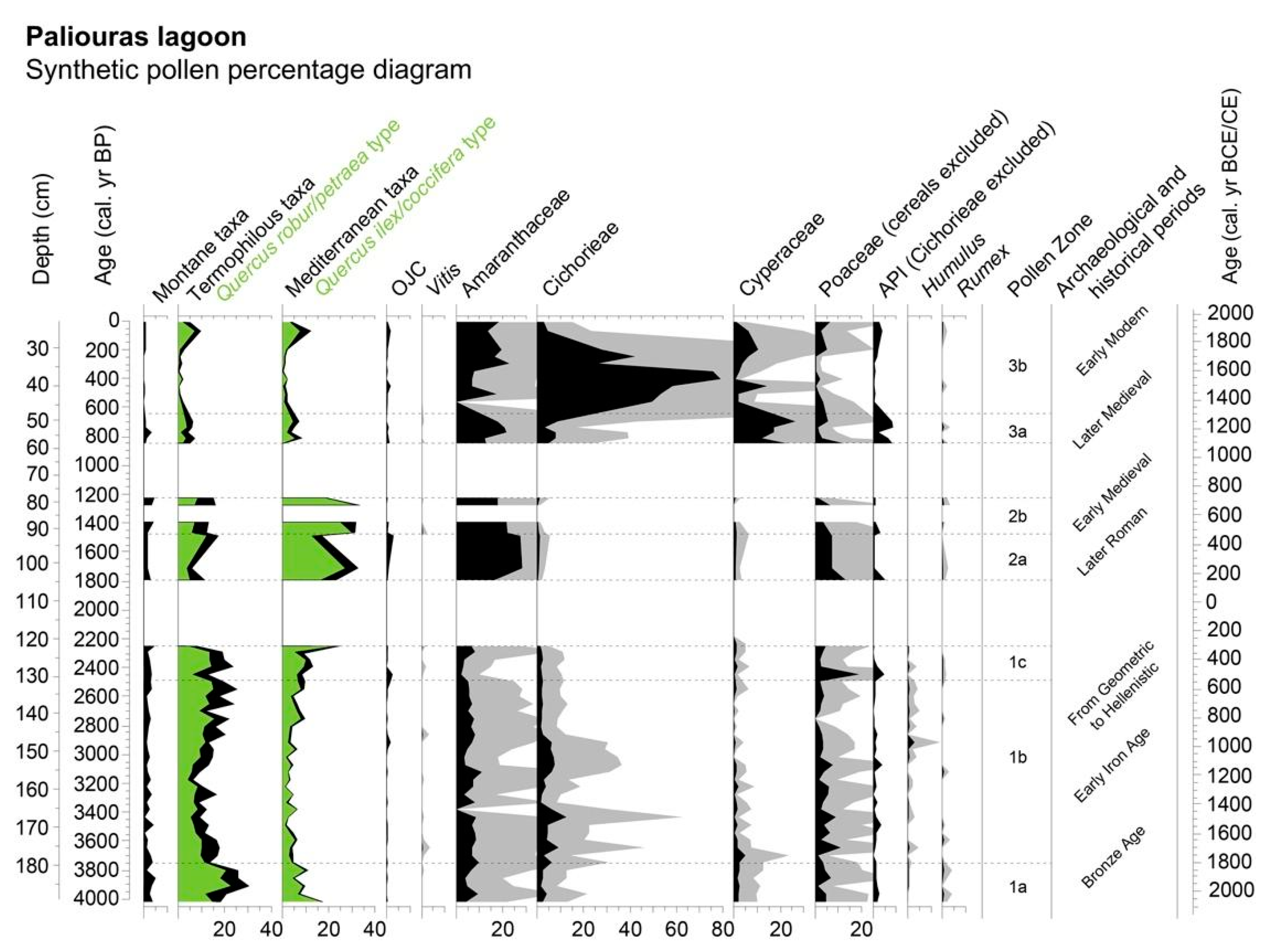
Pollen
4. Discussion
4.1. From the Bronze Age to the Hellenistic Period
4.2. The Later Roman and the Early Medieval Times
4.3. The Later Medieval and Early Modern Times
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, N.; Eastwood, W.J.; Kuzucuoglu, C.; Fiorentino, G.; Caracuta, V. Climatic, vegetation and cultural change in the eastern Mediterranean during the mid-Holocene environmental transition. Holocene 2011, 21, 147–162. [Google Scholar] [CrossRef]
- Kouli, K.; Masi, A.; Mercuri, A.; Florenzano, A.; Sadori, L. Regional Vegetation Histories: An Overview of the Pollen Evidence from the Central Mediterranean. In Environment and Society in the Long Late Antiquity; Izdebski, A., Mulryan, M., Eds.; Brill: Leiden, The Netherlands, 2019; pp. 69–82. [Google Scholar]
- Mercuri, A.M.; Florenzano, A.; Burjachs, F.; Giardini, M.; Kouli, K.; Masi, A.; Picornell-Gelabert, L.; Revelles, J.; Sadori, L.; Servera-Vives, G.; et al. From influence to impact: The multifunctional land-use in Mediterranean prehistory emerging from palynology of archaeological sites (8.0–2.8 ka BP). Holocene 2019, 29, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Clarke, C.L.; Greve Alsos, I.; Edwards, M.E.; Paus, A.; Gielly, L.; Haflidason, H.; Mangerud, J.; Regnell, C.; Hughes, P.D.M.; Svendsen, J.I.; et al. A 24,000-year ancient DNA and pollen record from the Polar Urals reveals temporal dynamics of arctic and boreal plant communities. Quat. Sci. Rev. 2020, 247, 106564. [Google Scholar] [CrossRef]
- Izdebski, A.; Guzowski, P.; Poniat, R.; Masci, L.; Palli, J.; Vignola, C.; Bauch, M.; Cocozza, C.; Fernandes, R.; Ljungqvist, F.C.; et al. Palaeoecological data indicates land-use changes across Europe linked to spatial heterogeneity in mortality during the Black Death pandemic. Nat. Ecol. Evol. 2022, 6, 297–306. [Google Scholar] [CrossRef]
- Seim, A.; Marquer, L.; Bisson, U.; Hofmann, J.; Herzig, F.; Kontic, R.; Lechterbeck, J.; Muigg, B.; Neyses-Eiden, M.; Rzepecki, A.; et al. Historical spruce abundance in Central Europe: A combined dendrochronological and palynological approach. Front. Ecol. Evol. 2022, 10, 909453. [Google Scholar] [CrossRef]
- Pérez-Obiol, R.; Sadori, L. Similarities and dissimilarities, synchronisms and diachronisms in the Holocene vegetation history of the Balearic Islands and Sicily. Veget. Hist. Archaeobot. 2007, 16, 259–265. [Google Scholar] [CrossRef]
- Heurtley, W.A. An archaeological reconnaissance of Greek Macedonia (west of the Struma) in the Neolithic, Bronze, and Early Iron Ages. In Prehistoric Macedonia; 1 map, 112 figures, 24 plates; University Press: Cambridge, UK, 1940. [Google Scholar]
- Kouli, K. Vegetation development and human activities in Attiki (SE Greece) during the last 5000 years. Veg. Hist. Archaeobot. 2012, 21, 267–278. [Google Scholar] [CrossRef]
- Morellón, M.; Anselmetti, F.S.; Ariztegui, D.; Brushulli, B.; Sinopoli, G.; Wagner, B.; Sadori, L.; Gilli, A.; Pambuku, A. Human–climate interactions in the central Mediterranean region during the last millennia: The laminated record of Lake Butrint (Albania). Quat. Sci. Rev. 2016, 136, 134–152. [Google Scholar] [CrossRef] [Green Version]
- Panajiotidis, S.; Papadopoulou, M.L. Human-landscape interactions in Halkidiki (NC Greece) over the last 3.5 millennia, revealed through palynological, and archaeological-historical archives. J. Archaeol. Sci. 2016, 7, 138–145. [Google Scholar] [CrossRef]
- Emmanouilidis, A.; Panagiotopoulos, K.; Kouli, K.; Avramidis, P. Late-Holocene paleoenvironmental and land-use changes in Western Greece based on a sediment record from Klisova lagoon. Holocene 2022, 32, 485–500. [Google Scholar] [CrossRef]
- Susini, D.; Vignola, C.; Goffredo, R.; Totten, D.M.; Masi, A.; Smedile, A.; De Martini, P.M.; Cinti, F.R.; Sadori, L.; Forti, L.; et al. Holocene palaeoenvironmental evolution of the southern margin of the Salpi lagoon (Apulia, southern Italy). Quat. Intern. 2022. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Triantaphyllou, M.; Karkanas, P.; Kouli, K.; Syrides, G.; Vouvalidis, K.; Palyvos, N.; Tsourou, T. Paleoenvironmental evolution and prehistoric human–environment, in the embayment of Palamari (Skyros Island, Greece) during Middle-Late Holocene. Quat. Int. 2010, 216, 41–53. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Kouli, K.; Tsourou, T.; Koukousioura, O.; Pavlopoulos, K.; Dermitzakis, M.D. Paleoenvironmental changes since 3000 BC in the coastal marsh of Vravron (Attica, SE Greece). Quat. Int. 2010, 216, 14–22. [Google Scholar] [CrossRef]
- Vignola, C.; Hättestrand, M.; Bonnier, A.; Finné, M.; Izdebski, A.; Katrantsiotis, C.; Kouli, K.; Liakopoulos, G.C.; Norström, E.; Papadaki, M.; et al. Mid-late Holocene vegetation history of the Argive Plain (Peloponnese, Greece) as inferred from a pollen record from ancient Lake Lerna. PLoS ONE 2022, 17, 0271548. [Google Scholar] [CrossRef] [PubMed]
- Koukousioura, O.; Triantafyllou, M.V.; Dimiza, M.D.; Pavlopoulos, K.; Syrides, G.; Vouvalidis, K. Benthic foraminiferal evidence for and Paleoenvironmental evolution of Holocene coastal plains in the Aegean Sea (Greece). Quat Int 2012, 261, 105–117. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Pavlopoulos, K.P.; Kouli, K.; Koukousioura, O.; Dimiza, M.D.; Aidona, E.; Syrides, G.; Pallikarakis, A.; Goiran, J.P.; Fouache, E. Multiproxy paleoenvironmental reconstruction: The Piraeus coastal plain case study. Bull. Geol. Soc. Greece 2016, 50, 478–487. [Google Scholar] [CrossRef]
- Kouli, K.; Triantaphyllou, M.; Pavlopoulos, K.; Tsourou, T.; Karkanas, P.; Dermitzakis, M.D. Palynological investigation of Holocene palaeoenvironmental changes in the coastal plain of Marathon (Attica, Greece). Geobios. 2009, 42, 43–51. [Google Scholar] [CrossRef]
- Kouli, K.; Triantaphyllou, M.V.; Koukousioura, O.; Dimiza, M.D.; Parinos, C.; Panagiotopoulos, I.P.; Tsourou, T.; Gogou, A.; Mavrommatis, N.; Syrides, G.; et al. Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece). Water 2021, 13, 1505. [Google Scholar] [CrossRef]
- Koukousioura, O.; Kouli, K.; Vouvalidis, K.; Aidona, E.; Karadimou, G.; Syrides, G. A multi-proxy approach for reconstructing environmental dynamics since the mid Holocene in Lake Ismarida (Thrace, N. Greece). Rev. Micropal. 2020, 68, 100443. [Google Scholar] [CrossRef]
- Tsigarida, E.B.; Xydopoulos, I. Halkidiki Landscape, Archaeology, and Ethnicity. In Mines, Olives and Monasteries: Aspects of Halkidiki’s Environment History; Gounaris, B.C., Ed.; Epikentro Publishers and PHAROS Books: Thessaloniki, Greece, 2015. [Google Scholar]
- Koukousioura, O.; Dimiza, M.D.; Kyriazidou, E.; Triantaphyllou, M.V.; Syrides, G.; Aidona, E.; Vouvalidis, K.; Panagiotopoulos, I.P.; Papadopoulou, L. Environmental evolution of the Paliouras coastal lagoon in the eastern Thermaikos gulf (Greece) during Holocene. Environ. Earth Sci. 2019, 78, 313. [Google Scholar] [CrossRef]
- Psimoulis, P.; Ghilardi, M.; Fouache, E.; Stiros, S. Subsidence and evolution of the Thessaloniki plain, Greece, based on historical levelling and GPS data. Eng. Geol. 2006, 90, 55–70. [Google Scholar] [CrossRef]
- Poulos, S.E.; Chronis, G.T.; Collins, M.B.; Lykousis, V. Thermaikos Gulf Coastal System, NW Aegean Sea: An overview of water sediment fluxes in relation to air–land–ocean interactions and human activities. J. Mar. Syst. 2000, 25, 47–76. [Google Scholar] [CrossRef]
- Syrides, G. Lithostratigraphical, Biostratigraphical and Paleogeographical Study of the Neogene-Quaternary Sedimentary Sequence of Chalkidiki Peninsula. Ph.D. Thesis, School of Geology, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1990. [Google Scholar]
- Athanasiadis, N. Dasiki Fytokoinoniologia (Forest Phytosociology); Giahoudis: Thessaloniki, Greece, 1985. [Google Scholar]
- Theodoropoulos, K.O. determination of the phytosociological units of the university forest Taxiarchi Halkidikis O kathorismos ton fytokoinoniologikon monadon tou panepistimiakou dasous Taxiarchi Halkidikis (Definition and Classification of the Plant Sociological Units of the University Forest of Taxiarchis Halkidiki). Unpublished. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1991. [Google Scholar]
- Athanasiadis, N.; Theodoropoulos, K.; Gerasimidis, A.; Eleftheriadou, E.; Tsiripidis, I.; Korakis, G. Monades vlastisis, tis zonis, ton aeifyllon platyfyllon tou Agiou Orous (Vegetation Units of the Evergreen Broadleaved Zone of the Holly Mountain); Eidiki Ekdosi Thessaloniki Politistiki Protevousa tis Evropis: Thessaloniki, Greece, 1998; p. 87. [Google Scholar]
- Pavlidis, G. I chloris kai i vlastisis tis chersonisou Sithonias Halkidikis (Flora and Vegetation of the Sithonia Peninsula in Halkidiki). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1976. [Google Scholar]
- Tsitsoni, T.; Karagiannakidou, V. Site quality and stand structure in Pinus halepensis forests of North Greece. Forestry 2000, 73, 51–64. [Google Scholar] [CrossRef]
- Katirtzidou, M.; Latinopoulos, P. Allocation of surface and subsurface water resources to competing uses under climate changing conditions: A case study in Halkidiki, Greece. Water Supply 2018, 18, 1151–1161. [Google Scholar] [CrossRef]
- Mamara, A.; Anadranistakis, M.; Argiriou, A.A.; Szentimrey, T.; Kovacs, T.; Bezes, A.; Bihari, Z. High resolution air temperature climatology for Greece for the period 1971–2000. Meteorol. Appl. 2017, 24, 191–205. [Google Scholar] [CrossRef] [Green Version]
- Tiverios, M. Greek colonisation of the Northern Aegean. In Greek Colonisation: An Account of Greek Colonies and Other Settlements Overseas; Tsetskhladze, G., Ed.; Brill: Leiden, The Netherlands, 2008. [Google Scholar]
- Poulter, A.G. The Transition to Late Antiquity: On the Danube and Beyond; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Snively, C.S. Macedonia in Late Antiquity. In A Companion to Ancient Macedonia; Roisman, J., Worthington, I., Eds.; Blackwell Publishing: West Sussex, UK, 2010. [Google Scholar]
- Smyrlis, K. Settlement and Environment in Halkidiki, Ninth to Fifteenth Century AD. In Mines, Olives and Monasteries. Aspects of Halkidiki’s Environmetal History; Gounaris, B.C., Ed.; Epikentro Publishers and Pharos Books: Thessaloniki, Greece, 2015; pp. 109–121. [Google Scholar]
- Karagiannē, P. Hoi Vyzantinoi Oikismoi Stē Makedonia mesa apo ta Archaiologika Dedomena (4os-5os Diōnas); University Studio Press: Thessalonikē, Greece, 2010. [Google Scholar]
- Soustal, P. Makedonien, Südlicher Teil; 2 Bände; Austrian Academy of Science Press: Vienna, Austria, 2022. [Google Scholar]
- Gogoue, A.; Triantaphyllou, M.V.; Xoplaki, E.; Izdebski, A.; Parinos, C.; Dimiza, M.D.; Bouloubassi, I.; Luterbacher, J.; Kouli, K.; Martrat, B.; et al. Climate variability and socio-environmental changes in the northern Aegean (NE Mediterranean) during the last 1500 years. Quat Sci Rev. 2016, 136, 209–228. [Google Scholar] [CrossRef]
- Kolovos, E.; Kotzageorgis, P. Halkidiki in the Early Modern Period: Towards an Environmental History. In Mines, Olives and Monasteries. Aspects of Halkidiki’s Environmetal History; Gounaris, B.C., Ed.; Epikentro Publishers and Pharos Books: Thessaloniki, Greece, 2015; pp. 123–161. [Google Scholar]
- Gounaris, V.K. Thessaloniki, 1830–1912: History, Economy and Society. In The Good Kingdom of Thessaloniki. History and Culture; Chasiotis, I.K., Ed.; Observer: Thessaloniki, Greece, 1997; pp. 117–133. [Google Scholar]
- Koliopoulos, I. The shaping of the new Macedonia (1798–1870). In The History of Macedonia; Koliopoulos, I., Ed.; Musesum of the Macedonian Struggle: Thessaloniki, Greece, 2007; pp. 167–179. [Google Scholar]
- Leipe, T.; Moros, M.; Kotilainen, A.; Vallius, H.; Kabel, K.; Endler, M.; Kowalski, N. Mercury in Baltic Sea sediments-natural background and anthropogenic impact. Chem. Erde-Geo-Chem. 2013, 73, 249–259. [Google Scholar] [CrossRef]
- Moros, M.; Andersen, T.J.; Schulz-Bull, D.; Häusler, K.; Bunke, D.; Snowball, I.; Kotilainen, A.; Zillén, L.; Jensen, J.B.; Kabel, K.; et al. Towards an event stratigraphy for Baltic Sea sediments deposited since AD 1900: Approaches and challenges. Boreas 2017, 46, 129–142. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Reimer, P.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725. [Google Scholar] [CrossRef]
- Heaton, T.J.; Köhler, P.; Butzin, M.; Bard, E.; Reimer, R.W.; Austin, W.E.N.; Ramsey, C.B.; Grootes, P.M.; Hughen, K.A.; Kromer, B.; et al. Marine20—The marine radiocarbon age calibration curve (0–55, 000 cal BP). Radiocarbon 2020, 62, 779–820. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis; John Wiley: Chichester, UK, 1989. [Google Scholar]
- Stockmarr, J. Tablets with spores used in absolute pollen analysis. Pollen Spores 1971, 13, 615–621. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de Botanique Historique et Palynologie; Universite’ d’Aix Marseille III: Marseille, France, 1992–1995–1998. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004. [Google Scholar]
- Berglund, B.E.; Ralska-Jasiewiczowa, M. Pollen Analysis and Pollen Diagrams. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; John Wiley & Sons: Chichester, UK, 1986; pp. 455–496. [Google Scholar]
- Grimm, E.C. Tilia and Tilia-graph: Pollen spreadsheet and graphics programs. In Proceedings of the 8th International Palynological Congress, Aixen-Provence, France, 6–12 September 1992; p. 56. [Google Scholar]
- Andersen, S.T. Identification of Wild Grasses and Cereal Pollen; Danmarks Geologiske Undersøgelse Årbog: Copenhagen, Danemark, 1979; pp. 69–92. [Google Scholar]
- Florenzano, A.; Marignani, M.; Rosati, L.; Fascetti, S.; Mercuri, A.M. Are Cichorieae an indicator of open habitats and pastoralism in current and past vegetation studies? Plant Biosyst. 2015, 149, 154–165. [Google Scholar] [CrossRef]
- Smit, A. A scanning electron microscopical study of the pollen morphology in the genus Quercus. Acta Bot. Neerl 1973, 22, 655–665. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E. Olea, Juglans and Castanea: The OJC group as pollen evidence of the development of human-induced environments in the Italian peninsula. Quat. Int. 2013, 303, 24–42. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E.; Torri, P. Anthropogenic Pollen Indicators (API) from archaeological sites as local evidence of human-induced environments in the Italian Peninsula. Ann. Bot. 2013, 3, 143–153. [Google Scholar]
- Sadori, L.; Giardini, M. Charcoal analysis, a method to study vegetation and climate of the Holocene: The case of Lago di Pergusa, Sicily (Italy). Geobios 2007, 40, 173–180. [Google Scholar] [CrossRef]
- Kokkinidou, D.; Trantalidou, K. Neolithic and Bronze Age Settlement in Western Macedonia. Annu. Br. Sch. Athens 1991, 86, 93–106. [Google Scholar] [CrossRef]
- Tsitsoni, T. Conditions determining natural regeneration after wildfires in the Pinus halepensis (Miller, 1768) forests of Kassandra Peninsula (North Greece). For. Ecol. Manag. 1997, 92, 199–208. [Google Scholar] [CrossRef]
- Palandjian, D.; Gitas, I.Z.; Wright, R. Burned area mapping and post-fire impact assessment in the Kassandra peninsula (Greece) using Landsat TM and Quickbird data. Geocart. Int. 2009, 24, 193–205. [Google Scholar] [CrossRef]
- Valamoti, S.M.; Gkatzogia, E.; Hristova, I.; Marinova, E. Iron Age Cultural Interactions, Plant Subsistence and Land Use in Southeastern Europe Inferred from Archaeobotanical Evidence of Greece and Bulgaria. In Archaeology across Frontiers and Borderlands. Fragmentation and Connectivity in the North Aegean and the Central Balkans from the Bronze Age to the Iron Age; Gimatzidis, S., Pieniążek, M., Mangaloğlu-Votruba, S., Eds.; Austrian Academy of Sciences: Vienna, Austria, 2018; pp. 269–290. [Google Scholar]
- Valamoti, S.M.; Gkatzogia, E.; Ntinou, M. Did Greek colonization bring olive growing to the north? An integrated archaeobotanical investigation of the spread of Olea europaea in Greece from the7th to the 1st millennium BC. Veget. Hist. Archaeobot. 2018, 27, 177–195. [Google Scholar] [CrossRef]
- Jahns, S. On the Holocene vegetation history of the Argive Plain (Peloponnese, southern Greece). Veget. Hist. Archaeobot. 1993, 2, 187–203. [Google Scholar] [CrossRef]
- Zangger, E.; Timpson, M.E.; Yazvenko, S.B.; Kuhnke, F.; Knauss, J. The Pylos Regional Archaeological Project, II: Landscape evolution and site preservation. Hesperia 1997, 66, 549–641. [Google Scholar] [CrossRef]
- Greig, J.R.A.; Turner, J. Some pollen diagrams from Greece and their archaeological significance. J. Archaeol. Sci. 1974, 1, 177–194. [Google Scholar] [CrossRef]
- Leontarakis, P.K.; Xatzianastasiou, L.I.; Theodorou, J.A. Biological Aspects of the Lagoon Cockle, Cerastoderma glaucum (Poiret 1879), in a Coastal Lagoon in Keramoti, Greece in the Northeastern Mediterranean. J. Shellfish Res. 2008, 27, 1171–1175. [Google Scholar] [CrossRef]
- Mirzoeva, A.T.; Demchenko, N.A. Morphological response of lagoon cockle Cerastoderma glaucum (Poiret, 1789) to eutrophication in the Sea of Azov. In Proceedings of the Conference Series: Earth and Environmental Science, Kryvyi Rih, Ukraine, 24–27 May 2022; Volume 1049, p. 012059. [Google Scholar] [CrossRef]
- Cline, E.H. The Oxford Handbook of the Bronze Age Aegean (ca. 3000–1000 BC); Oxford UniversityPress: Oxford, UK, 2010. [Google Scholar]
- Psomiadis, D.; Dotsika, E.; Albanakis, K.; Ghaleb, B.; Hillaire-Marcel, C. Speleothem record of climatic changes in the northern Aegean region (Greece) from the Bronze Age to the collapse of the Roman Empire. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 489, 272–283. [Google Scholar] [CrossRef]
- Finné, M.; Woodbridge, J.; Labuhn, I.; Roberts, C.N. Holocene hydro-climatic variability in the Mediterranean: A synthetic multi-proxy reconstruction. Holocene 2019, 29, 847–863. [Google Scholar] [CrossRef]
- Andreou, S. Northern Aegean. In The Oxford Handbook of the Bronze Age Aegean; Cline, E., Ed.; Oxford University Press: Oxford, UK, 2010; pp. 643–659. [Google Scholar]
- Kyrikou, S.; Kouli, K.; Triantaphyllou, M.V.; Dimiza, M.D.; Gogou, A.; Panagiotopoulos, I.P.; Anagnostou, C.; Karageorgis, A.P. Late Glacial and Holocene vegetation patterns of Attica: A high-resolution record from Elefsis Bay, southern Greece. Quat. Int. 2020, 545, 28–37. [Google Scholar] [CrossRef]
- Marinova, E.; Tonkov, S.; Bozilova, E.; Vajsov, I. Holocene anthropo-genic landscapes in the Balkans: The palaeobotanical evidence from southwestern Bulgaria. Veget. Hist. Archaeobot. 2012, 21, 413–427. [Google Scholar] [CrossRef]
- Lazarova, M.; Koutsios, A.; Kontopoulos, N. Holocene vegetation history of the Kotihi Lagoon (north-western Peloponnesus, Greece). Quat Int. 2012, 261, 138–145. [Google Scholar] [CrossRef]
- Tiverios, M.A.; Paspalas, S.A. Andros and its Colonies. J. Mediterr. Archaeol. 2012, 25, 273–281. [Google Scholar]
- Trabaud, L.; Galtié, J.F. Effects of fire frequency on plant communities and landscape pattern in the Massif des Aspres (southern France). Landsc. Ecol. 1996, 11, 215–224. [Google Scholar] [CrossRef]
- Atherden, M. Human impact on the vegetation of southern Greece and problems of palynological interpretation: A case study from Crete. In Landscape and Land Use in Postglacial Greece; Halstead, P., Frederick, C., Eds.; Sheffield Academic Press: Sheffield, UK, 2000; pp. 62–78. [Google Scholar]
- Carrión, Y.; Kaal, J.; López-Sáez, J.A.; López-Merino, L.; Martínez Cortizas, A. Holocene vegetation changes in NW Iberia revealed by anthracological and palynological records from a colluvial soil. Holocene 2010, 20, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Göktürk, O.M.; Fleitmann, D.; Badertscher, S.; Cheng, H.; Edwards, R.L.; Leuenberger, M.; Fankhauser, A.; Tüysüz, O.; Kramers, J. Climate on the southern Black Sea coast during the Holocene: Implications from the Sofular Cave record. Quat. Sci. Rev. 2011, 30, 2433–2445. [Google Scholar] [CrossRef]
- Cheng, H.; Sinha, A.; Verheyden, S.; Nader, F.H.; Li, X.L.; Zhang, P.Z.; Yin, J.J.; Yi, L.; Peng, Y.B.; Rao, Z.G.; et al. The climate variability in northern Levant over the past 20,000 years. Geophys. Res. Lett. 2015, 42, 8641–8650. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, J.K. Euboians in Macedonia? Acloser look. Oxf. J. Archaeol. 1996, 15, 151–181. [Google Scholar] [CrossRef]
- Norström, E.; Katrantsiotis, C.; Finné, M.; Risberg, J.; Smittenberg, R.; Bjursäter, S. Biomarker hydrogen isotope composition (δD) as proxy for Holocene hydro-climatic change and seismic activity in SW Peloponnese, Greece. J Quat Sci. 2018, 33, 563–574. [Google Scholar] [CrossRef]
- Katrantsiotis, C.; Norström, E.; Smittenberg, R.H.; Finné, M.; Weiberg, E.; Hättestrand, M. Climate changes in the Eastern Mediterranean over the last 5000 years and their links to the high-latitude atmospheric patterns and Asian monsoons. Glob. Planet Chang. 2019, 175, 36–51. [Google Scholar] [CrossRef]
- van Geel, B.; Coope, G.R.; van der Hammen, T. Palaeoecology and stratigraphy of the Late Glacial type section at Usselo (The Netherlands). Rev. Palaeobot. Palynol. 1989, 60, 25–129. [Google Scholar] [CrossRef]
- Paperna, J.; Overstreet, R.M. Parasites and diseases of mullets (Mugilidae). In Aquaculture of Grey Mullets; Oren, O.H., Ed.; Cambridge University Press: Cambridge, UK, 1981; pp. 411–493. [Google Scholar]
- Fabbrocini, A.; Di Matteo, O.; D’Adamo, R. Abra segmentum (Mollusca:Pelecypoda) of the Lesina Lagoon (Southern Adriatic coast, Italy): Observationson variations in the population in relation to the main environmental parameters. Transit. Waters Bull. 2008, 1, 39–44. [Google Scholar]
- Atherden, M.; Hall, J.; Wright, J. A pollen diagram from the north-east Peloponnese, Greece: Implications for vegetation history and archaeology. Holocene 1993, 3, 351–356. [Google Scholar] [CrossRef]
- Weiberg, E.; Bevan, A.; Kouli, K.; Katsianis, M.; Woodbridge, J.; Bonnier, A.; Engel, M.; Finné, M.; Fyfe, R.; Maniatis, Y.; et al. Long-term trends of land use and demography in Greece: A comparative study. Holocene 2019, 29, 742–760. [Google Scholar] [CrossRef] [Green Version]
- Pagnoux, C.; Bouby, L.; Valamoti, S.M.; Bonhomme, V.; Ivorra, S.; Gkatzogia, E.; Karathanou, A.; Kotsachristou, D.; Kroll, H.; Terral, J.F. Local domestication or diffusion? Insights into viticulture in Greece from Neolithic to Archaic times, using geometric morphometric analyses of archaeological grape seeds. J. Archaeol. Sci. 2021, 125, 105263. [Google Scholar] [CrossRef]
- Mercuri, A.; Accorsi, C.; Bandini Mazzanti, M. The long history of Cannabis and its cultivation by the Romans in central Italy, shown by pollen records from Lago Albano and Lago di Nemi. Veget. Hist. Archaeobot. 2002, 11, 263–276. [Google Scholar] [CrossRef]
- Wilson, D.G. Plant remains from the Graveney boat and the early history of Humulus lupulus L. in W. Europe. New Phytol. 1975, 75, 627–664. [Google Scholar] [CrossRef]
- Behre, K.E. The history of beer additives in Europe—A review. Veget. Hist. Archaeobot. 1999, 8, 35–48. [Google Scholar] [CrossRef]
- Edwardson, J.R. Hops—Their botany, history, production and utilization. Econ. Bot. 1952, 6, 160–175. [Google Scholar] [CrossRef]
- Bar-Matthews, M.; Ayalon, A. Speleothems as paleoclimate indicators, a case study from Soreq Cave located in the Eastern Mediterranean region, Israel. In Past Climate Variability through Europe and Africa; Battarbee, R.W., Gasse, F., Stickley, C.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 363–391. [Google Scholar]
- Finné, M.; Holmgrena, K.; Sundqvista, H.S.; Weiberg, E.; Lindblom, M. Climate in the eastern Mediterranean, and adjacent regions, during the past 6000 years—A review. J. Arch. Sci. 2011, 38, 3153–3173. [Google Scholar] [CrossRef]
- Sadori, L.; Masi, A.; Ricotta, C. Climate-driven past fires in central Sicily. Plant Biosyst. 2015, 149, 166–173. [Google Scholar] [CrossRef]
- Kuhnt, T.; Schmiedl, G.; Ehrmann, W.; Andersen, H.Y. Stable isotopic composition of Holocene benthic foraminifers from the Eastern Mediterranean Sea: Past changes in productivity and deep water oxygenation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 268, 106–115. [Google Scholar] [CrossRef]
- Panagiotopoulos, K.; Aufgebauer, A.; Schäbitz, F.; Wagner, B. Vegetation and climate history of the Lake Prespa region since the Late Glacial. Quatern. Int. 2013, 293, 157–169. [Google Scholar] [CrossRef]
- Masi, A.; Francke, A.; Pepe, C.; Thienemann, M.; Wagner, B.; Sadori, L. Vegetation history and paleoclimate at Lake Dojran (FYROM/Greece) during the Late Glacial and Holocene. Clim Past. 2018, 14, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Kouli, K.; Dermitzakis, M.D. Contributions to European Pollen Database: Lake Orestiás (Kastoria, northern Greece). Grana 2010, 49, 154–156. [Google Scholar] [CrossRef]
- Kouli, K. Plant landscape and land use at the Neolithic lake settlement of Dispilió (Macedonia, Northern Greece). Plant Biosyst. 2015, 149, 145–204. [Google Scholar] [CrossRef]
- Donders, T.; Panagiotopoulos, K.; Koutsodendris, A.; Bertini, A.; Mercuri, A.M.; Masi, A.; Combourieu-Nebout, N.; Joannin, S.; Kouli, K.; Kousis, I.; et al. 1.36 million years of Mediterranean forest refugium dynamics in response to glacial–interglacial cycle strength. Proc. Natl. Acad. Sci. USA 2021, 118, e2026111118. [Google Scholar] [CrossRef]
- Bottema, S. Late Quaternary History of Northwestern Greece. Ph.D. Thesis, University of Gronigen, Groningen, The Netherlands, 1974. [Google Scholar]
- Bottema, S. The Holocene history of walnut, sweet-chestnut, manna-ash and plane tree in the Eastern Mediterranean. Pallas 2000, 52, 35–59. [Google Scholar]
- Bottema, S. On the history of the walnut (Juglans regia L.) in southeastern Europe. Acta Bot. Neerl. 1980, 29, 343–349. [Google Scholar] [CrossRef]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veget. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Del Lungo, S.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; et al. Rethinking the history of common walnut (Juglans regia L.) in Europe: Its origins and human interactions. PLoS ONE 2017, 12, e0172541. [Google Scholar] [CrossRef] [Green Version]
- Alcock, J.P. Food in the Ancient World; Greenwood Press: Westport, CT, USA, 2006. [Google Scholar]
- Behre, K.E. The history of rye cultivation in Europe. Veget. Hist. Archaeobot. 1992, 1, 141–156. [Google Scholar] [CrossRef]
- Jasny, N. Competition among Grains in Classical Antiquity. Am Hist Rev. 1942, 47, 747–764. [Google Scholar] [CrossRef]
- Squatriti, P. Rye’s rise and Rome’s fall: Agriculture and climate in Europe during Late Antiquity. In Environment and Society during the Long Late Antiquity; Izdebski, A., Mulryan, M., Eds.; Brill: Leiden, The Netherlands, 2018; pp. 342–354. [Google Scholar]
- Lüning, S.; Schulte, L.; Garcés-Pastor, S.; Danladi, I.B.; Gałka, M. The Medieval Climate Anomaly in the Mediterranean region. Paleoceanogr. Paleoclimatol. 2019, 34, 1625–1649. [Google Scholar] [CrossRef]
- Xoplaki, E.; Fleitmann, D.; Luterbacher, J.; Wagner, S.; Zorita, E.; Telelis, I.; Toreti, A.; Izdebski, A. The Medieval Climate Anomaly and Byzantium: A review of the evidence on climatic fluctuations, economic performance and societal change. Quat. Sci. Rev. 2016, 136, 229–252. [Google Scholar] [CrossRef] [Green Version]
- Lefort, J. Villages de Macédoine 1. La Chalcidique occidentale. Revue des études byzantines 42. 1984, p. 325. Available online: https://www.persee.fr/doc/rebyz_0766-5598_1984_num_42_1_2162_t1_0325_0000_1 (accessed on 12 November 2022).
- Lefort, J.; Bellier, P. Paysages de Macédoine: Leurs Caractères Leur Évolution À Travers Les Documents et les Récits des Voyageurs; De Boccard: Paris, France, 1986. [Google Scholar]
- Izdebski, A.; Pickett, J.; Roberts, N.; Waliszewski, T. The environmental, archaeological and historical evidence for climatic changes and their societal impacts in the Eastern Mediterranean in Late Antiquity. Quat. Sci. Rev. 2016, 136, 189–208. [Google Scholar] [CrossRef]
- Kouli, K. Tracing human impact on a mountainous plant landscape in Rhodopi Mt (N. Greece) during the last 1100 years. Rev. Micropaleontol. 2020, 68, 100442. [Google Scholar] [CrossRef]
- Kotzageorgis, P.P. Nomads (Yürüks) and Environment in Early Modern Halkidiki. Balk. Stud. 2015, 50, 95–118. [Google Scholar]
- Liakopoulos, G.C. The Integration of Settlers into Existing Socio-Environmental Settings: Reclaiming the Greek Lands After the Late Medieval Crisis. In Perspectives on Public Policy in Societal-Environmental Crises. Risk, Systems and Decisions; Izdebski, A., Haldon, J., Filipkowski, P., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Bottema, S. Palynological investigations in Greece with special reference to pollen as an indicator of human activity. Palaeohistoria 1982, 24, 257–288. [Google Scholar]
- Lebreton, V.E.; Messager, L.; Marquer, J.; Renault-Miskovsky, A. neotaphonomic experiment in pollen oxidation and its implications for archaeopalynology. Rev. Palaeobot. Palynol. 2010, 162, 29–38. [Google Scholar] [CrossRef]
- Tsiamis, C. Plague in Byzantine Times: A Medico-Historical Study; De Gruyter: Berlin, Germany; Boston, MA, USA, 2022. [Google Scholar]
- Vacalopoulos, A.E. History of Thessaloniki 316 BC–1983; Ant. Stamoulis: Thessaloniki, Greece, 1983. [Google Scholar]
- Chasiotis, I.K. Turkish-occupied Thessaloniki: The first period (15th century–1830). In To the good kingdoms of Thessaloniki. History and culture; Chasiotis, I.K., Ed.; Observer: Thessaloniki, Greece, 1997; pp. 103–116. [Google Scholar]
- Moustakas, K. Methodological issues in the approximation of population sizes of the Late Byzantine City. In Byzantine cities (8th–15th Century). Research Perspectives and New Interpretive Approaches; Kiousopoulou, T., Ed.; Publications of the Faculty of Philosophy of the University of Crete: Rethymno, Greece, 2012; pp. 225–251. [Google Scholar]
- Hütteroth, W.-D. Ecology in the Ottoman Lands. In The Cambridge History of Turkey; The Later Ottoman Empire, 1603–1839; Faroqhi, S.N., Ed.; Cambridge University Press: Cambridge, UK, 2008; Volume 3, pp. 18–43. [Google Scholar]
- Bellier, P.; Bondoux, R.C.; Cheynet, J.C.; Geyer, B.; Grélois, J.P.; Kravari, V. Paysages de Macédoine: Leurs Caractères, Leur évolution à Travers Les Documents et Les Récits Des Voyageurs, Présenté Par Jacques Lefort; Travaux et Mémoires du Centre de Recherche d’Histoire et Civilisation de Byzance; Collège de France, De Boccard: Paris, France, 1986; Title of Unpublished Work. [Google Scholar]
- Xoplaki, E.; Maheras, P.; Luterbacher, J. Variability of Climate in Meridional Balkans during the Periods 1675–1715 and 1780–1830 and its Impact on Human Life. Clim. Change 2001, 48, 581–615. [Google Scholar] [CrossRef]
- Lespez, L. Geomorphic responses to long-term land use changes in Eastern Macedonia (Greece). Catena 2003, 51, 181–208. [Google Scholar] [CrossRef]
- Roberts, N.; Moreno, A.; Valero-Garcés, B.L.; Corella, J.P.; Jones, M.; Allcock, S.; Woodbridge, J.; Morellón, M.; Luterbacher, J.; Xoplaki, E.; et al. Palaeolimnological evidence for an east- west climate see-saw in the Mediterranean since AD 900. Glob. Planet Change 2012, 84, 23–34. [Google Scholar] [CrossRef]
- Morellón, M.; Sinopoli, G.; Izdebski, A.; Sadori, L.; Anselmetti, F.; Hodges, R.; Regattieri, E.; Wagner, B.; Brushulli, B.; Ariztegui, D. Environment, Climate and Society in Roman and Byzantine Butrint. In Environment and Society in the Long Late Antiquity; Izdebski, A., Mulryan, M., Eds.; Brill: Leiden, The Netherlands, 2018. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (cm) | Lab ID | Dated Material | Radiocarbon Age | Probability | Cal. Age 2σ BP | Cal. Age 2σ BCE/CE |
|---|---|---|---|---|---|---|
| 19 | 137Cs | 1954 CE | ||||
| 20.5 | DSH10237_SO | Organic matter | - | 100% | −5.5 ± 1 | 1955.5 ± 1 CE |
| 45.5 * | DSH9902_SO | Organic matter | 1131 ± 21 | 95% | 1014 ± 110 | 936 ± 110 CE |
| 47.5 | DSH10238_SO | Organic matter | 706 ± 23 | 90% | 663 ± 34 | 1287 ± 34 CE |
| 92.5 | DSH9849_SH | Cerithium vulgatum shell | 2055 ± 39 | 93% | 2020.5 ± 203 | 70.5 ± 203 BCE |
| 122.5 * | DSH9900_SO | Organic matter | 3788 ± 33 | 90% | 4171.5 ± 177 | 2221.5 ± 177 BCE |
| 191.5 | DSH10652_SO | Bulk | 3726 ± 21 | 100% | 4068 ± 164 | 2118 ± 164 BCE |
| Year CE | Total Population (Estimation) | Ottoman Register Signature |
|---|---|---|
| 1478 | 464 | TT7–MM17748 |
| 1527 | 3005 | TT403 |
| 1568 | 2738 | TKGM186 |
| 1697 | 4749 | MAD3461 |
| 1722 | 4778 | KK2869 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masci, L.; Vignola, C.; Liakopoulos, G.C.; Kouli, K.; Koukousioura, O.; Aidona, E.; Moros, M.; Vouvalidis, K.; Izdebski, A.; Masi, A. Landscape Response to Dynamic Human Pressure in the Paliouras Lagoon, Halkidiki Peninsula, Macedonia, Greece. Quaternary 2022, 5, 54. https://doi.org/10.3390/quat5040054
Masci L, Vignola C, Liakopoulos GC, Kouli K, Koukousioura O, Aidona E, Moros M, Vouvalidis K, Izdebski A, Masi A. Landscape Response to Dynamic Human Pressure in the Paliouras Lagoon, Halkidiki Peninsula, Macedonia, Greece. Quaternary. 2022; 5(4):54. https://doi.org/10.3390/quat5040054
Chicago/Turabian StyleMasci, Lucrezia, Cristiano Vignola, Georgios C. Liakopoulos, Katerina Kouli, Olga Koukousioura, Elina Aidona, Matthias Moros, Konstantinos Vouvalidis, Adam Izdebski, and Alessia Masi. 2022. "Landscape Response to Dynamic Human Pressure in the Paliouras Lagoon, Halkidiki Peninsula, Macedonia, Greece" Quaternary 5, no. 4: 54. https://doi.org/10.3390/quat5040054