
Lateglacial to Mid-Holocene Vegetation History in the Eastern Vale of Pickering, Northeast Yorkshire, UK: Pollen Diagrams from Palaeolake Flixton

Abstract
:1. Introduction
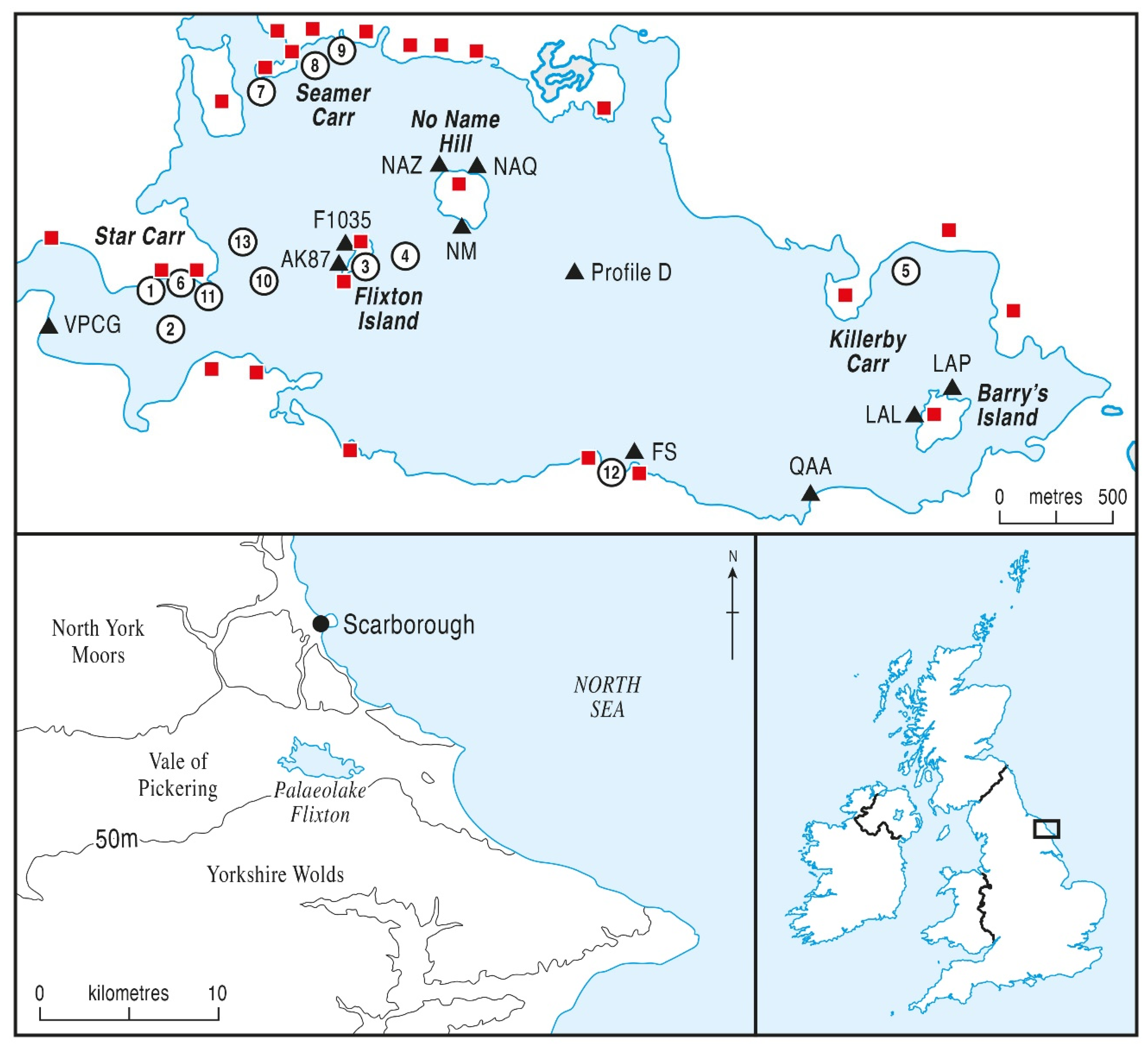
2. Study Area and Sites
3. Materials and Methods
3.1. Sampling
3.2. Pollen and Microcharcoal Analysis
3.3. Radiocarbon Dating
4. Results
4.1. Lake Flixton Centre (Profile D)
4.1.1. Pollen Assemblage Zones
- D1
- 660–620 cm Betula-Salix
- D2
- 620–564 cm Betula-Juniperus-Salix
- D3
- 564–542 cm Betula-Salix-Juniperus
- D4
- 542–500 cm Betula-Pinus-Salix-Juniperus
- D5
- 500–465 cm Betula-Pinus-Salix-Juniperus-Ulmus
- D6
- 465–425 cm Corylus-Betula
- D7
- 425–319 cm Corylus-Ulmus-Quercus-Betula
- D8
- 319–206 cm Corylus-Ulmus-Quercus-Alnus
- D9
- 206–200 cm Quercus-Corylus-Alnus
4.1.2. Interpretation
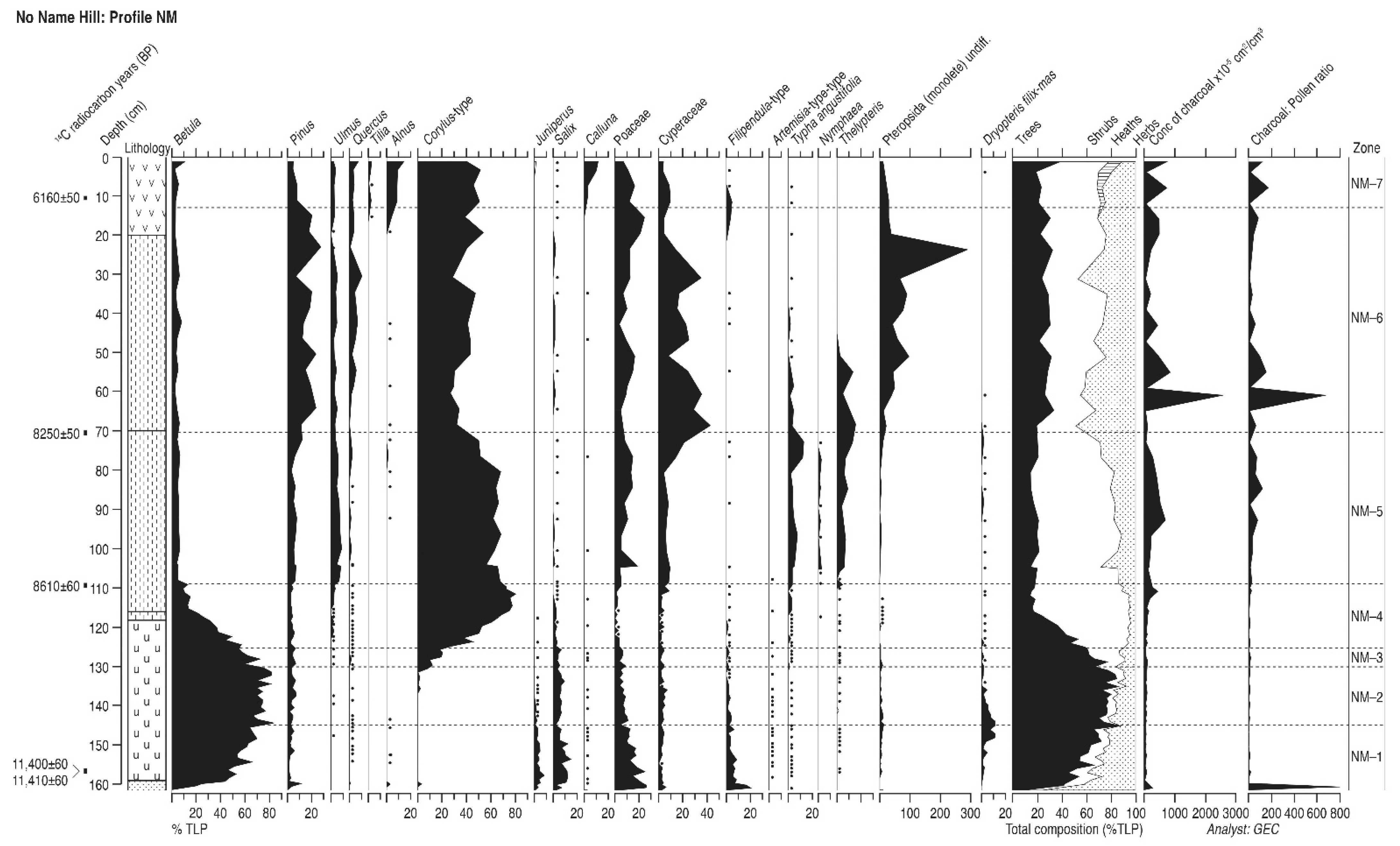
4.2. No-Name Hill (Profile NM)
4.2.1. Pollen Assemblage Zones
- NM-1
- 162–144 cm Betula-Salix-Juniperus
- NM-2
- 144–129 cm Betula-Salix
- NM-3
- 129–125 cm Betula-Corylus-Salix
- NM-4
- 125–109 cm Corylus-Betula
- NM-5
- 109–70 cm Corylus-Pinus-Ulmus
- NM-6
- 70–13 cm Corylus-Pinus-Ulmus-Quercus
- NM-7
- 13–0 cm Corylus-Alnus-Pinus-Ulmus-Quercus
4.2.2. Interpretation
4.3. No-Name Hill (Profile NAQ)
4.3.1. Pollen Assemblage Zones
- NAQ-1
- 207–189 cm Betula-Salix-Juniperus
- NAQ-2
- 189–171 cm Betula-Pinus-Salix-Juniperus
- NAQ-3
- 171–166 cm Betula-Pinus-Salix
- NAQ-4
- 166–162 cm Betula-Pinus-Corylus
- NAQ-5
- 162–154 cm Corylus-Betula-Pinus
- NAQ-6
- 154–136 cm Corylus-Pinus-Betula
4.3.2. Interpretation
4.4. No-Name Hill (Profile NAZ)
4.4.1. Pollen Assemblage Zones
- NAZ-1
- 47–39 cm Betula-Pinus-Salix
- NAZ-2
- 39–26 cm Betula-Pinus-Corylus
- NAZ-3
- 26–10 cm Corylus-Pinus-Betula
4.4.2. Interpretation
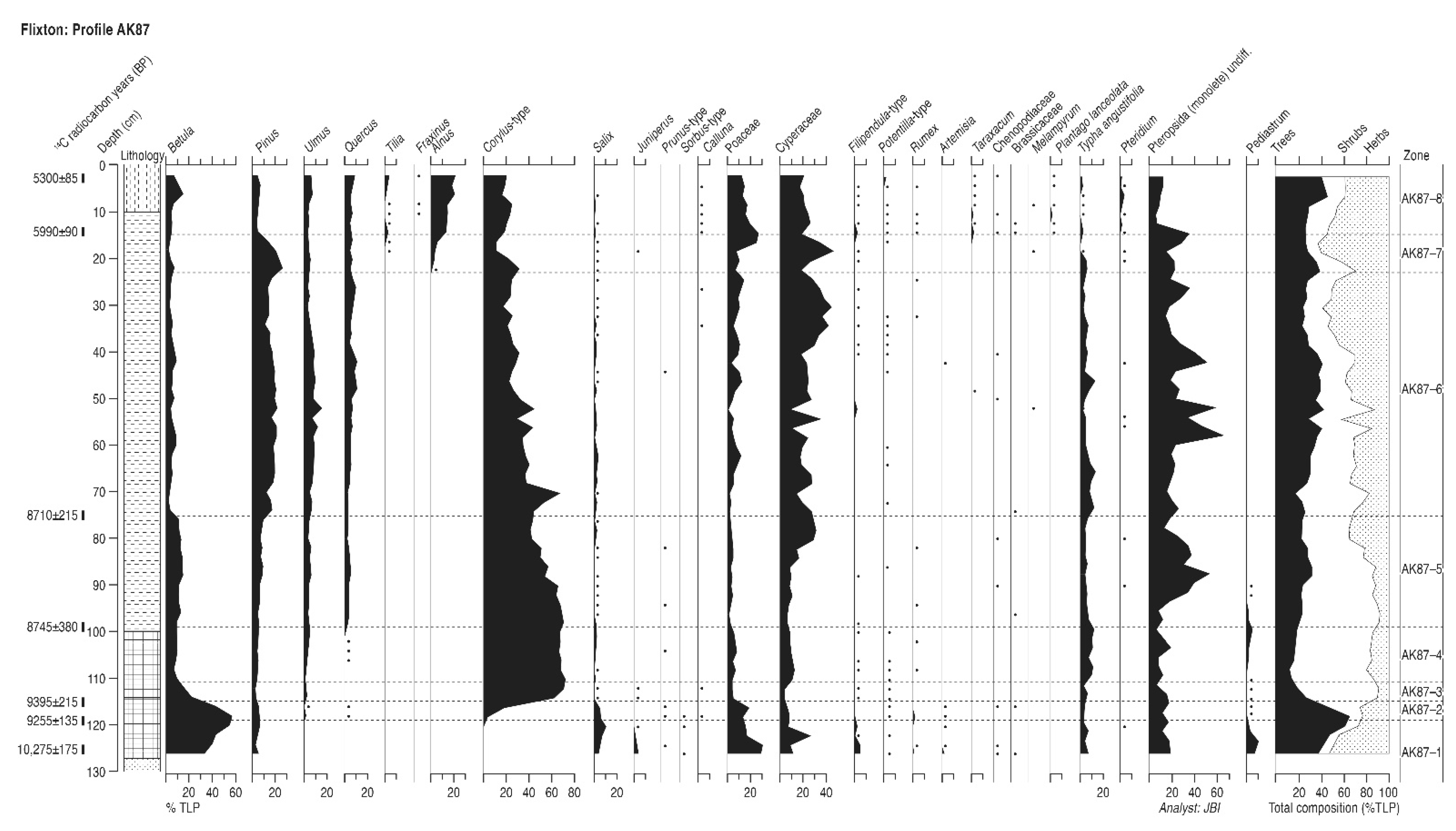
4.5. Flixton Island (Profile AK87)
4.5.1. Pollen Assemblage Zones
- AK87-1
- 126–119 cm Betula-Salix-Juniperus
- AK87-2
- 119–115 cm Betula-Corylus-Salix
- AK87-3
- 115–111 cm Betula-Corylus
- AK87-4
- 111–99 cm Corylus-Betula-Pinus-Ulmus
- AK87-5
- 99–75 cm Corylus-Betula-Pinus-Ulmus-Quercus
- AK87-6
- 75–23 cm Pinus-Corylus-Ulmus-Quercus
- AK87-7
- 23–15 cm Pinus-Corylus-Ulmus-Quercus-Alnus
- AK87-8
- 15–2 cm Alnus-Corylus-Ulmus-Quercus-Tilia
4.5.2. Interpretation
4.6. Flixton Island (Profile 1035)
4.6.1. Pollen Assemblage Zones
- F1035-1
- 50 cm Corylus-Betula-Pinus
- F1035-2
- 31–25 cm Corylus-Betula-Ulmus
- F1035-3
- 25–21 cm Corylus-Ulmus-Pinus-Quercus
- F1035-4
- 21–17 cm Corylus-Pinus-Ulmus-Quercus-Betula
- F1035-5
- 17–12 cm Corylus-Betula-Ulmus-Quercus-Alnus
4.6.2. Interpretation
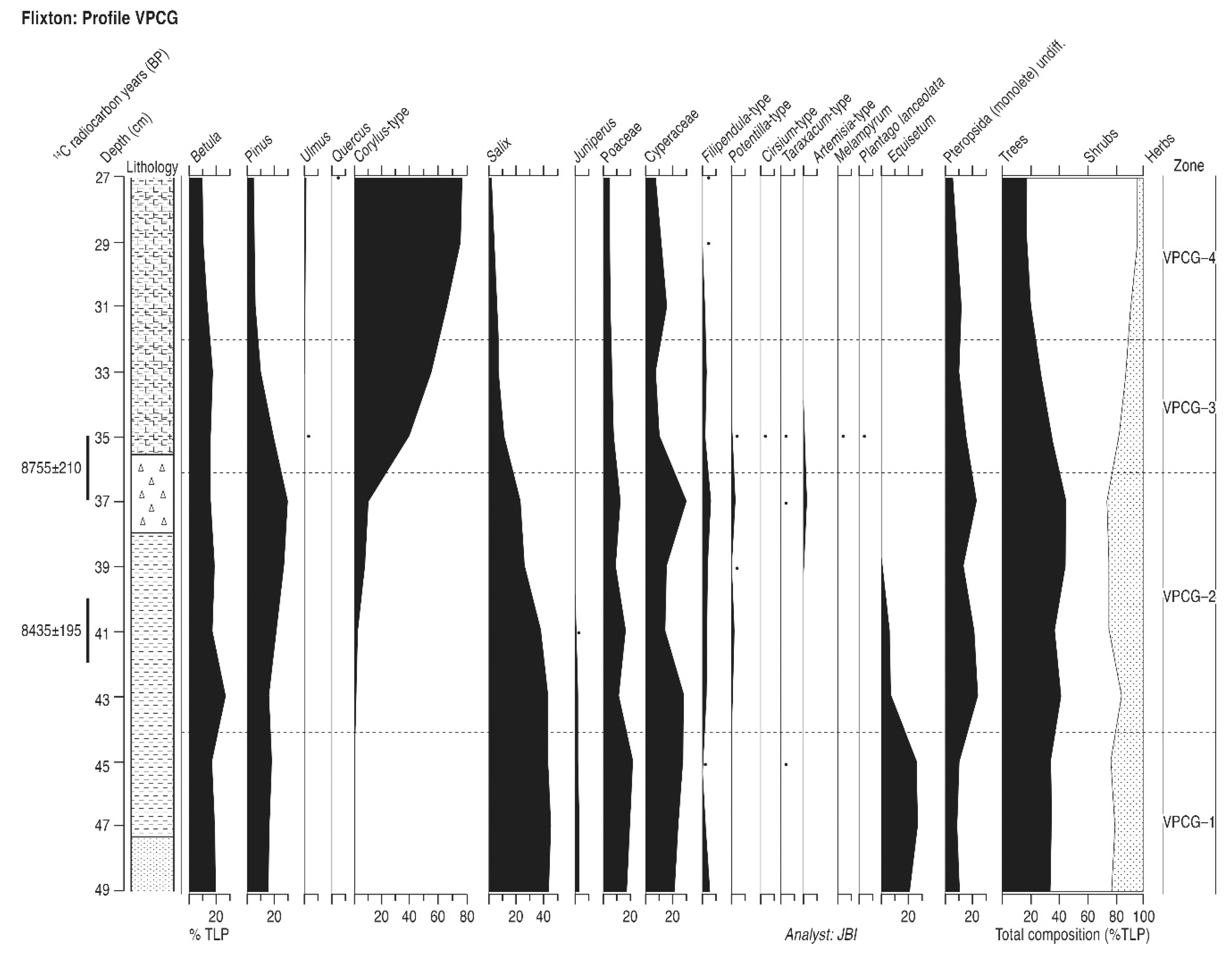
4.7. Flixton Carr (Profile VPCG)
4.7.1. Pollen Assemblage Zones
- VPCG-1
- 49–44 cm Salix-Betula-Pinus-Juniperus
- VPCG-2
- 44–36 cm Salix-Pinus-Betula-Corylus
- VPCG-3
- 36–32 cm Corylus-Betula-Pinus-Salix
- VPCG-4
- 32–27 cm Corylus-Betula
4.7.2. Interpretation
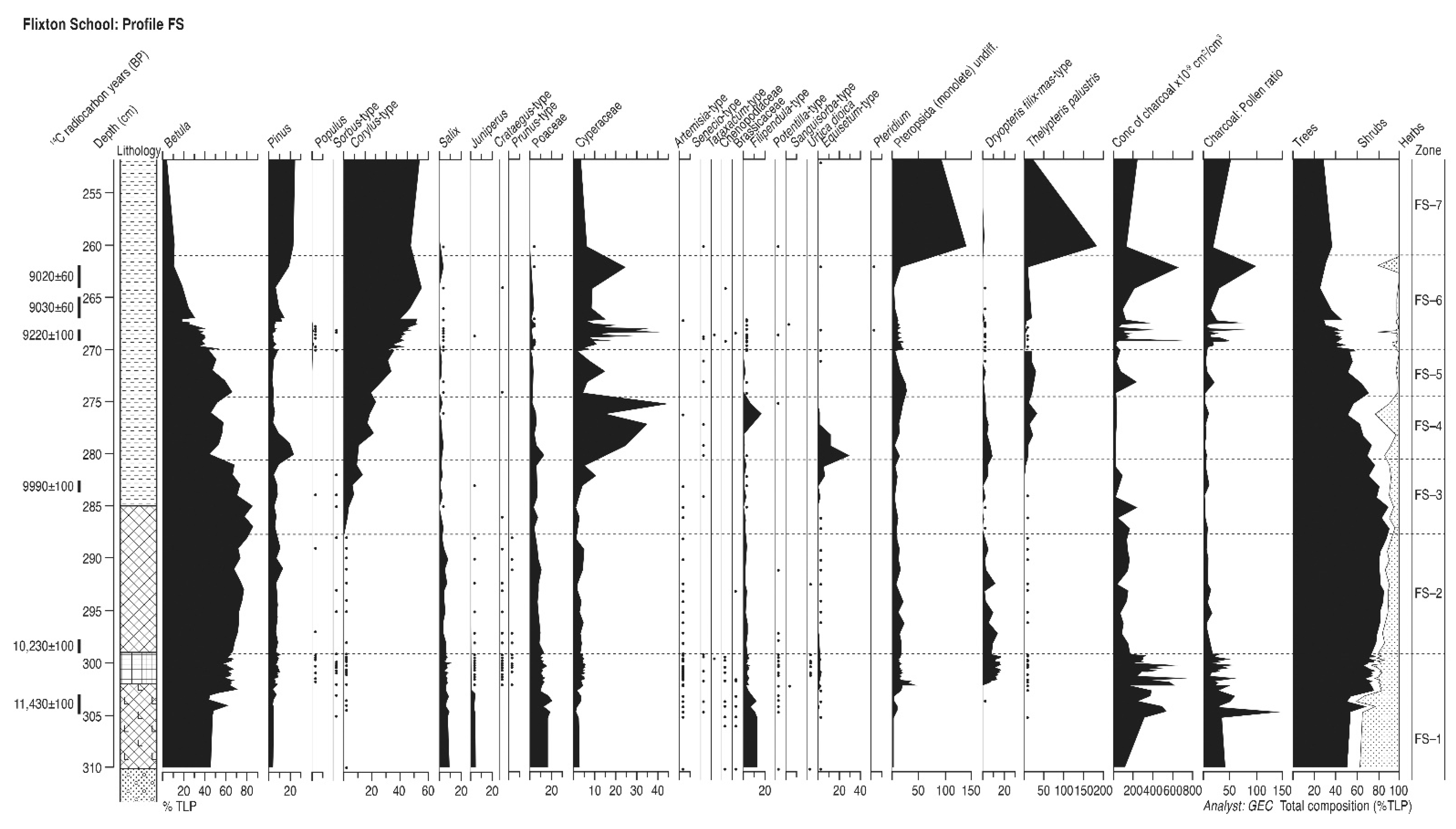
4.8. Flixton School (Profile FS)
4.8.1. Pollen Assemblage Zones
- FS-1
- 310–299 cm Betula-Salix-Pinus
- FS-2
- 299–288 cm Betula-Pinus-Salix
- FS-3
- 288–281 cm Betula-Corylus-Pinus
- FS-4
- 281–274 cm Betula-Pinus-Corylus-Salix
- FS-5
- 274–270 cm Betula-Corylus
- FS-6
- 270–261 cm Corylus-Betula-Pinus
- FS-7
- 261–253 cm Corylus-Pinus-Betula
4.8.2. Interpretation
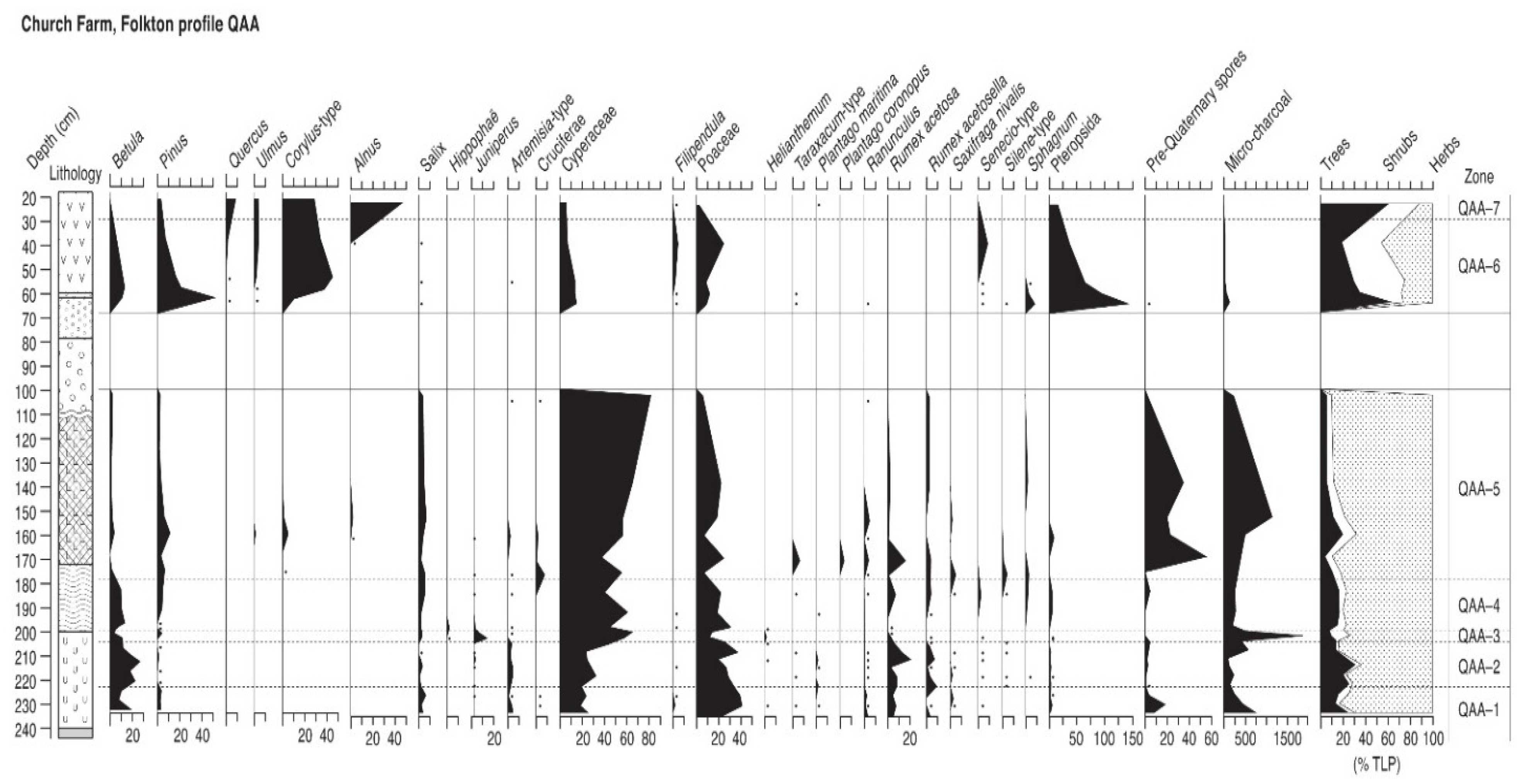
4.9. Church Farm Folkton (Profile QAA)
4.9.1. Pollen Assemblage Zones
- QAA-1
- 232–222 cm Betula-Salix-Pinus
- QAA-2
- 222–204 cm Betula-Salix
- QAA-3
- 204–197 cm Juniperus-Salix-Betula
- QAA-4
- 197–178 cm Betula-Salix-Pinus
- QAA-5
- 178–102 cm Salix-Pinus
- QAA-6
- 62–28 cm Corylus-Pinus-Betula
- QAA-7
- 28–22 cm Corylus-Alnus-Quercus-Ulmus
4.9.2. Interpretation
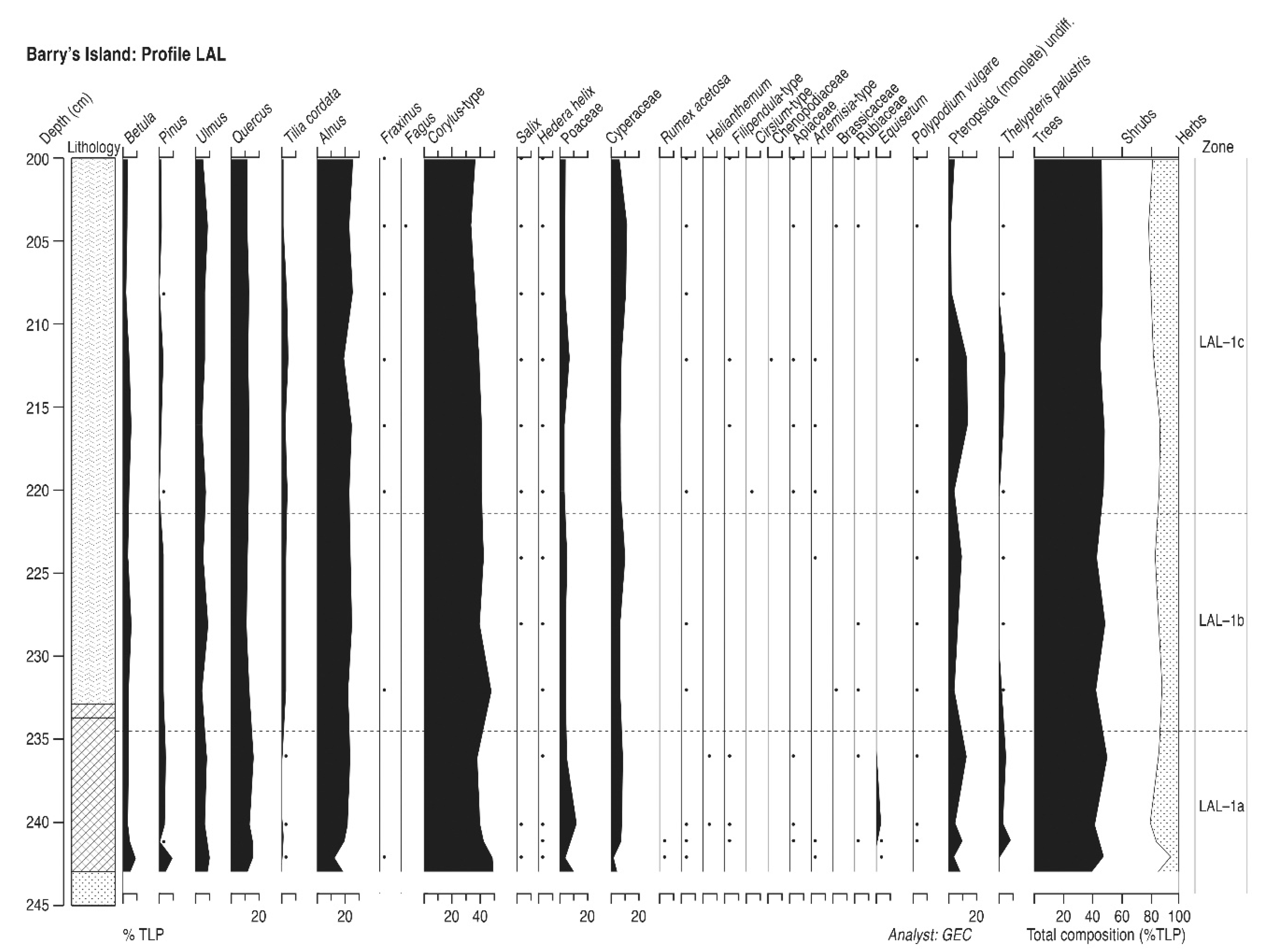
4.10. Barry’s Island (Profile LAL)
4.10.1. Pollen Assemblage Zones
- LAL-1
- 244–200 cm Corylus-Alnus-Quercus-Ulmus
4.10.2. Interpretation
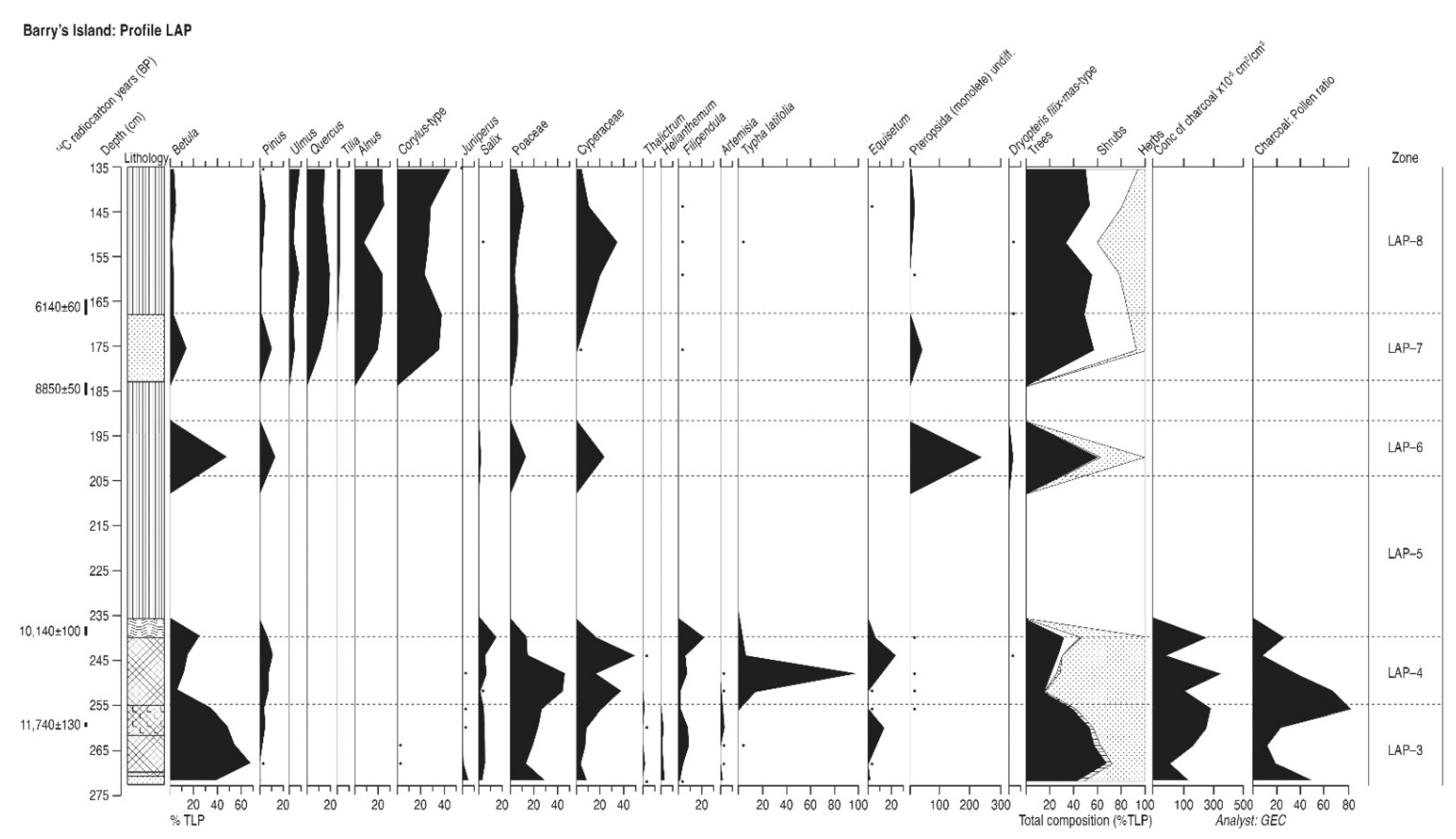
4.11. Barry’s Island (Profile LAP)
4.11.1. Pollen Assemblage Zones
- LAP-1
- 272–254 cm Betula-Salix-Pinus
- LAP-2
- 254–239 cm Pinus-Betula-Salix
- LAP-3
- 200 cm Betula-Pinus
- LAP-4
- 182–167 cm Corylus-Alnus-Quercus-Pinus
- LAP-5
- 167–135 cm Corylus-Alnus-Quercus-Ulmus
4.11.2. Interpretation
5. Discussion
5.1. Depositional Environments and Microfossil Data
5.2. Vegetation History Conspectus
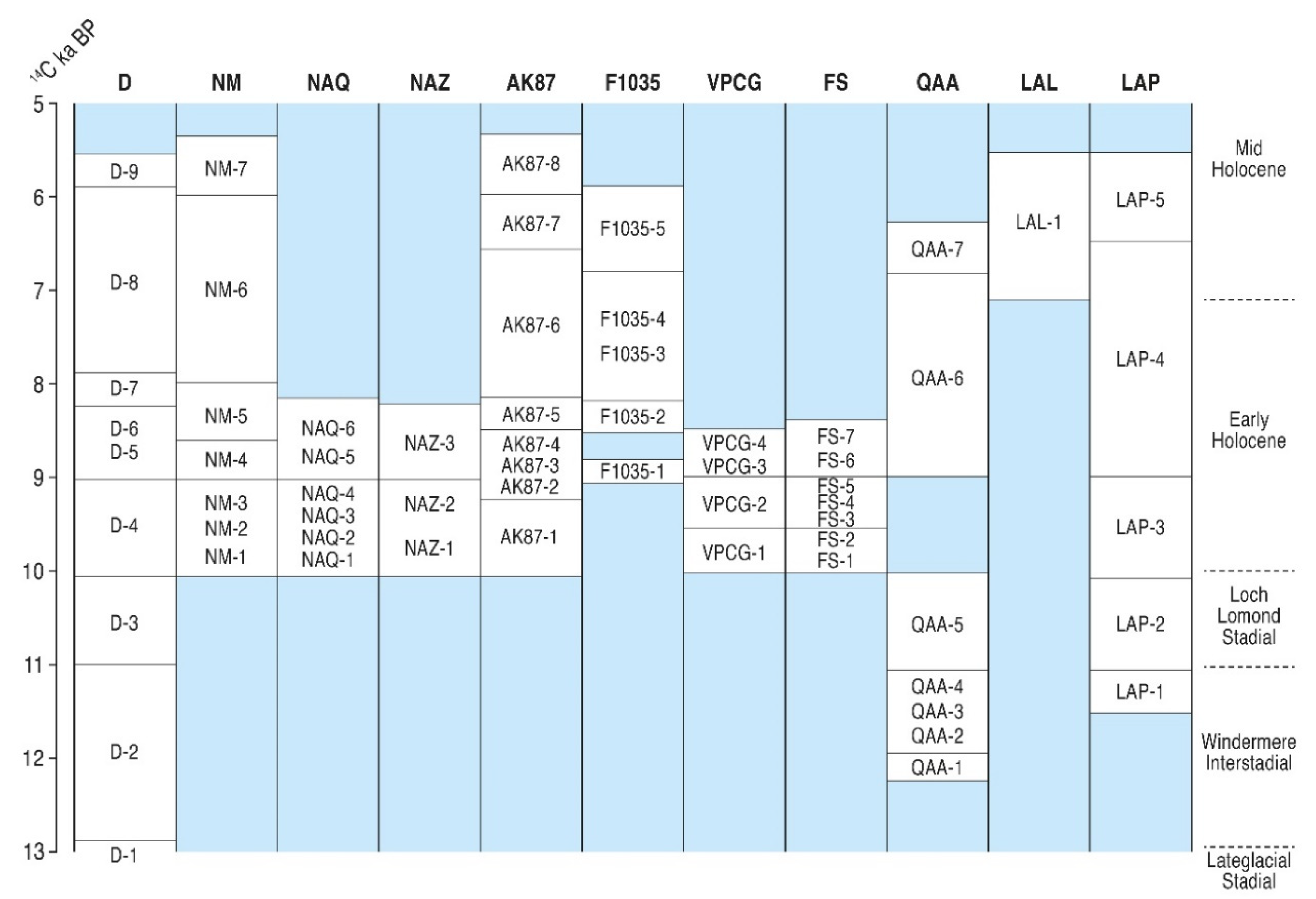
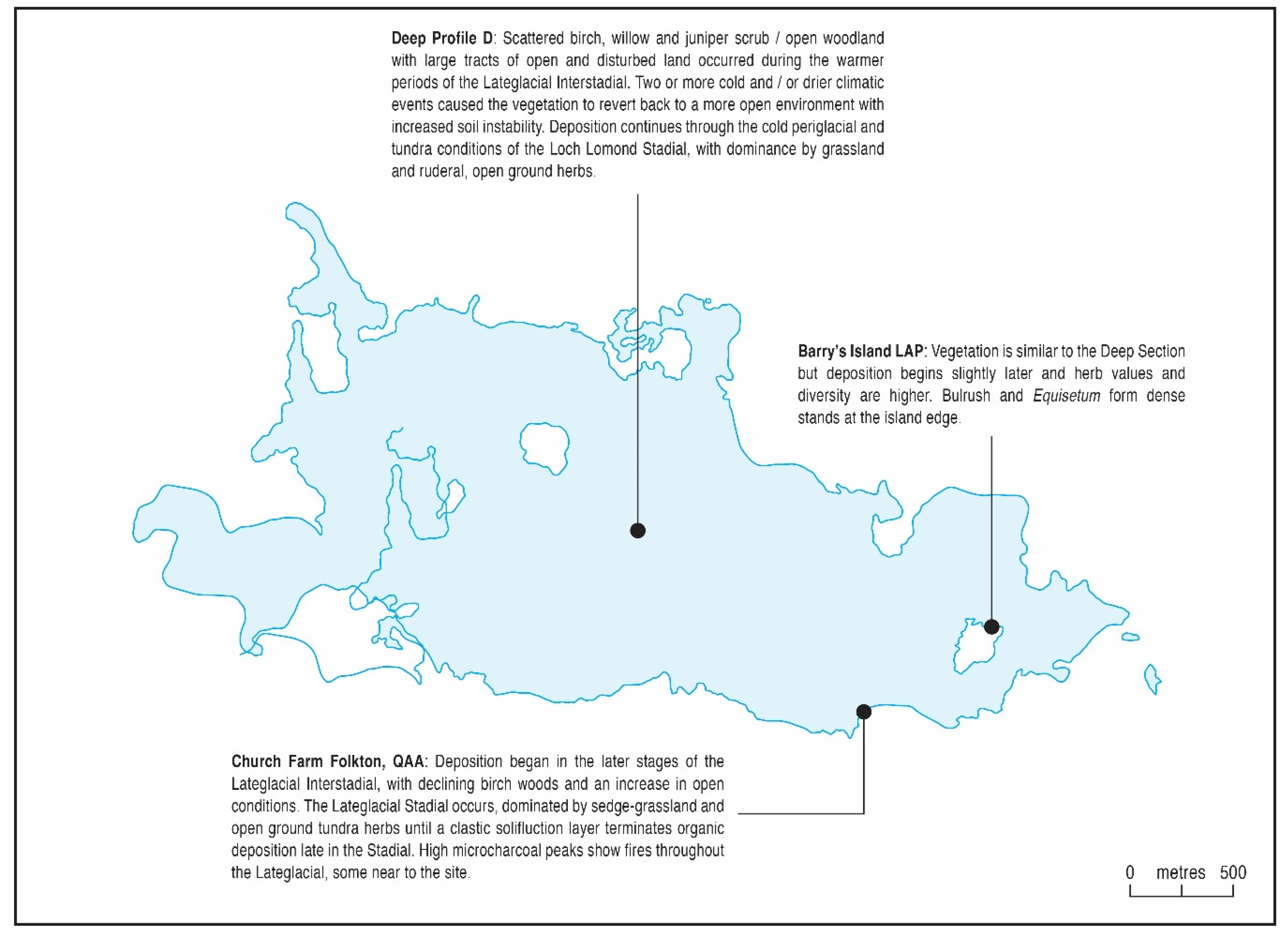
5.2.1. The Lateglacial Period (>10,000 14C BP; zones D 1–4, QAA 4–5 and LAP 1–2)
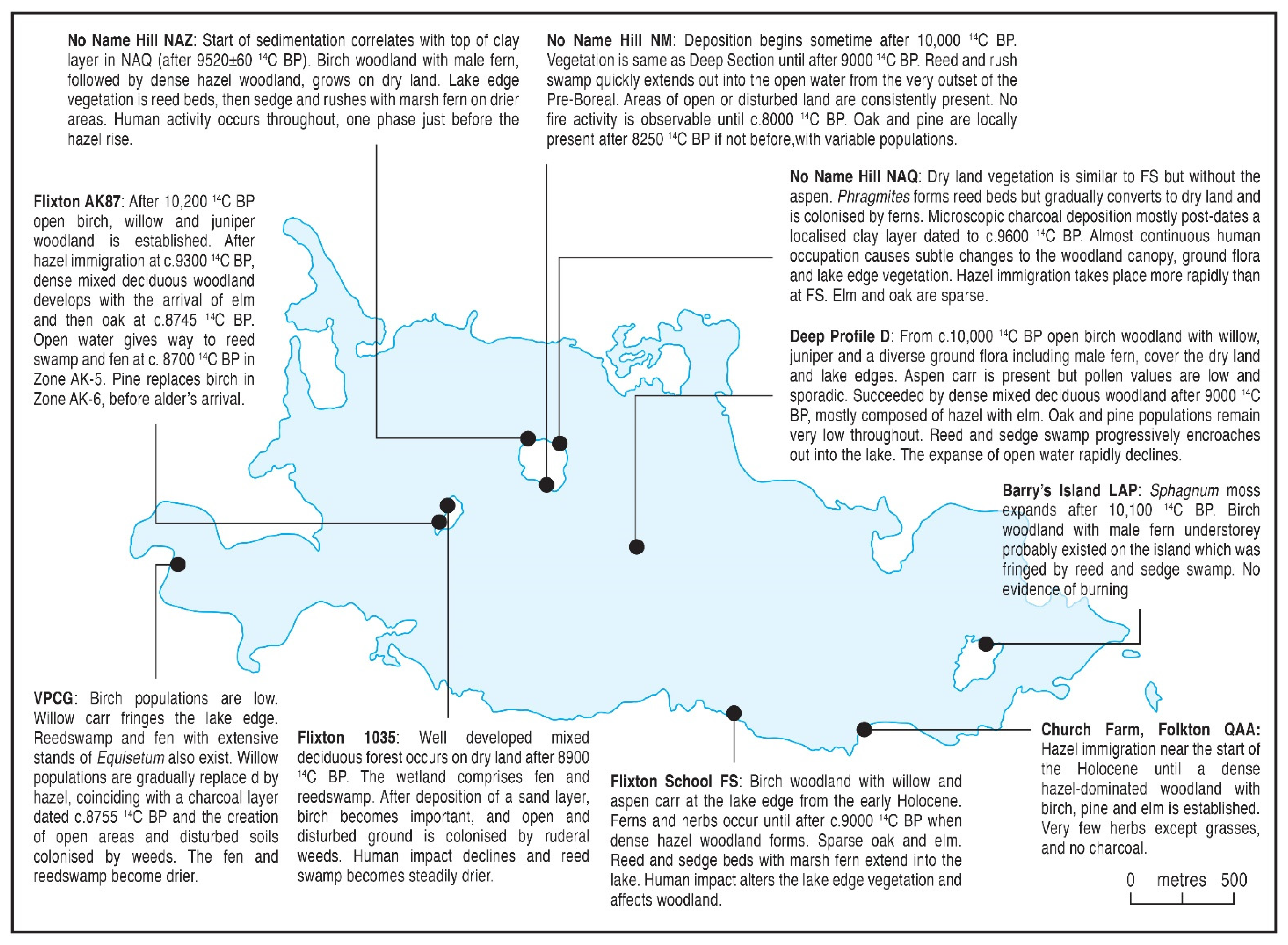
5.2.2. Early to Mid-Holocene (c.10,000 to 7000 14C BP; cf. Pre-Boreal and Boreal)
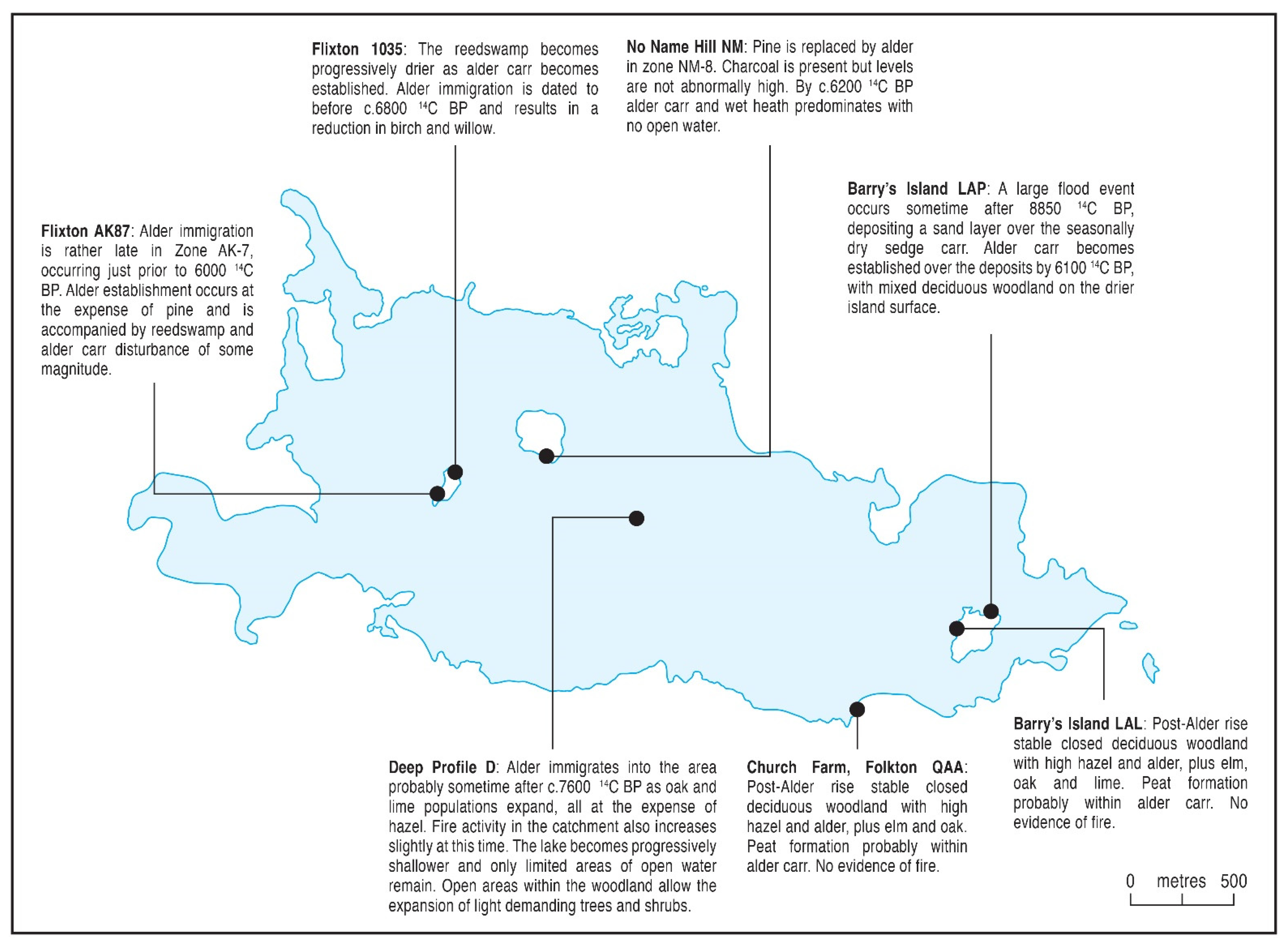
5.2.3. Mid-Holocene c.7000 to 5000 14C BP (cf. Atlantic; Zones D 8, NM 7, AK87 8, 1035 5, QAA 7, LAL 1 and LAP 4–5)
5.2.4. Later Holocene < 5000 14C BP (cf. Sub-Boreal)
5.3. Vegetation History Drivers around Lake Flixton
5.3.1. Climate
5.3.2. Human Impacts
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellars, P.; Dark, S.P. Star Carr in Context; McDonald Institute: Cambridge, UK, 1998. [Google Scholar]
- Innes, J.B.; Blackford, J.J.; Simmons, I.G. Mesolithic environments at Star Carr, the eastern vale of Pickering and environs: Local and regional contexts. J. Wetl. Archaeol. 2011, 11, 85–108. [Google Scholar] [CrossRef]
- Dark, P. Pollen and preservation at Star Carr: A 60-year perspective. J. Archaeol. Sci. Rep. 2017, 13, 240–248. [Google Scholar] [CrossRef]
- Milner, N.; Lane, P.; Taylor, B.; Conneller, C.; Schadla-Hall, T. Star Carr in a postglacial landscape: 60 years of research. J. Wetl. Archaeol. 2011, 11, 1–19. [Google Scholar] [CrossRef]
- Milner, N.; Taylor, B.; Conneller, C.; Schadla-Hall, T. Star Carr: Life in Britain after the Ice Age; Council for British Archaeology: York, UK, 2013. [Google Scholar]
- Milner, N.; Taylor, B.; Conneller, C. Star Carr Volume 1: A Persistent Place in a Changing World; White Rose University Press: York, UK, 2018. [Google Scholar]
- Milner, N.; Taylor, B.; Conneller, C. Star Carr Volume 2: Studies in Technology, Subsistence and Environment; White Rose University Press: York, UK, 2018. [Google Scholar]
- Clark, J.G.D. Excavations at Star Carr; Cambridge University Press: Cambridge, UK, 1954. [Google Scholar]
- Conneller, C.; Milner, N.; Taylor, B.; Taylor, M. Substantial settlement in the European Early Mesolithic: New research at Star Carr. Antiquity 2012, 86, 1004–1020. [Google Scholar] [CrossRef]
- Walker, D.; Godwin, H. Lake stratigraphy, pollen-analysis and vegetational history. In Excavations at Star Carr; Clark, J.G.D., Ed.; Cambridge University Press: Cambridge, UK, 1954; pp. 25–69. [Google Scholar]
- Conneller, C.; Milner, N.; Schadla-Hall, T.; Taylor, B. The temporality of the Mesolithic landscape: New work at Star Carr. In Chronology and Evolution within the Mesolithic of North-West Europe; Crombé, P., Van Strydonck, M., Sergant, J., Boudin, M., Bats, M., Eds.; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2009; pp. 77–94. [Google Scholar]
- Conneller, C.; Milner, N.; Schadla-Hall, R.T.; Taylor, B. Star Carr in the new millenium. In From Bann Flakes to Bushmills. Papers in Honour of Professor Peter Woodman; Finlay, N., McCartan, S., Milner, N., Wickham-Jones, C., Eds.; Prehistoric Society: Oxford, UK, 2009; pp. 78–88. [Google Scholar]
- Bayliss, A.; Taylor, B.; Bronk Ramsey, C.; Dunbar, E.; Kromer, B.; Bamforth, M.; Conneller, C.; Elliott, B.; Knight, B.; Milner, N. Dating the archaeology and environment of the Star Carr embayment. In Star Carr Volume 2: Studies in Technology, Subsistence and Environment; Milner, N., Conneller, C., Taylor, B., Eds.; White Rose University Press: York, UK, 2018; pp. 33–112. [Google Scholar]
- Schadla-Hall, R.T.; Cloutman, E.W. ‘One cannot dig at random in a peat bog.’ The eastern Vale of Pickering and the archaeology of a buried landscape. In Archaeology from the Ploughsoil; Haselgrove, C.C., Millet, M., Smith, I.M., Eds.; University of Sheffield Department of Archaeology and Prehistory: Sheffield, UK, 1985; pp. 77–86. [Google Scholar]
- Schadla-Hall, R.T. Recent investigations of the early Mesolithic landscape in the Vale of Pickering. In Mesolithic Northwest Europe: Recent Trends; Rowley-Conwy, P.A., Zvelebil, M., Blankholm, H., Eds.; University of Sheffield Department of Archaeology and Prehistory: Sheffield, UK, 1987; pp. 46–54. [Google Scholar]
- Schadla-Hall, R.T. The Vale of Pickering in the early Mesolithic in context. In The Mesolithic in Europe; Bonsall, C., Ed.; John Donald: Edinburgh, UK, 1989; pp. 218–224. [Google Scholar]
- Schadla-Hall, R.T. The Vale of Pickering in the Mesolithic: Uncovering the early post-glacial landscape. Arch. Int. 2000, 4, 11–13. [Google Scholar]
- Day, S.P.; Mellars, P. ‘Absolute’ dating of Mesolithic human activity at Star Carr, Yorkshire: New palaeoecological studies and the identification of the 9600 BP radiocarbon ‘plateau’. In Proceedings of the Prehistoric Society; Cambridge University Press: Cambridge, UK, 1994; Volume 60, pp. 417–422. [Google Scholar] [CrossRef]
- Conneller, C.; Schadla-Hall, T. Beyond Star Carr: The Vale of Pickering in the 10th millennium BP. Proc. Prehist. Soc. 2003, 69, 85–106. [Google Scholar] [CrossRef]
- Dark, P. Palaeoenvironmental investigations. In Star Carr in Context: New Archaeological and Palaeoecological Investigations at the Early Mesolithic site of Star Carr, North Yorkshire; Mellars, P.A., Dark, P., Eds.; McDonald Institute for Archaeological Research: Cambridge, UK, 1998; pp. 111–181. [Google Scholar]
- Cloutman, E.W. Palaeoenvironments in the Vale of Pickering. Part 1: Stratigraphy and palaeogeography of Seamer Carr, Star Carr and Flixton Carr. Proc. Prehist. Soc. 1988, 54, 1–19. [Google Scholar] [CrossRef]
- Cloutman, E.W. Palaeoenvironments in the Vale of Pickering. Part 2: Environmental history at Seamer Carr. Proc. Prehist. Soc. 1988, 54, 21–36. [Google Scholar] [CrossRef]
- Cloutman, E.W.; Smith, A.G. Palaeoenvironments in the Vale of Pickering Part 3: Environmental history at Star Carr. Proc. Prehist. Soc. 1988, 54, 37–58. [Google Scholar] [CrossRef]
- Taylor, B. Early Mesolithic activity in the wetlands of the Lake Flixton basin. J. Wetl. Archaeol. 2011, 11, 63–84. [Google Scholar] [CrossRef]
- Taylor, B. Early Holocene wetland succession at Lake Flixton (UK) and its implications for Mesolithic settlement. Veget. Hist. Archaeobot. 2019, 28, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Abrook, A. Vegetation changes at Palaeolake Flixton during the Late-glacial and early Holocene periods. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 129–134. [Google Scholar]
- Blockley, S.; Matthews, I.; Palmer, A.; Candy, I.; Kearney, R.; Langdon, P.; Langdon, C.; Abrook, A.; Lincoln, P.; Farry, A.; et al. Climate Research. In Star Carr Volume 2: Studies in Technology, Subsistence and Environment; Milner, N., Conneller, C., Taylor, B., Eds.; White Rose University Press: York, UK, 2018; pp. 113–121. [Google Scholar]
- Day, S.P. Devensian Late-Glacial and Early Flandrian environmental history of The Vale of Pickering. J. Quat. Sci. 1996, 11, 9–24. [Google Scholar] [CrossRef]
- Innes, J.B. An interim report on stratigraphic and pollen analytical research in the Vale of Pickering. Val. Pick. Res. Trust Newslett. 1994, 1, 4–6. [Google Scholar]
- Cummins, G.E. Impacts of Hunter-Gatherers on the Vegetation History of the Eastern Vale of Pickering, Yorkshire. Ph.D. Thesis, Durham University, Durham, UK, 2003. [Google Scholar]
- Spikins, P. GIS models of past vegetation: An example from northern England, 10,000–5000 BP. J. Archaeol. Sci. 2000, 27, 219–234. [Google Scholar] [CrossRef]
- Taylor, B. Early Holocene wetland succession at Like Flixton. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 148–154. [Google Scholar]
- Taylor, B.; Elliott, B.; Conneller, C.; Milner, N.; Bayliss, A.; Knight, B.; Bamforth, M. Resolving the issue of artefact deposition at Star Carr. Proc. Prehist. Soc. 2017, 83, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.; Blockley, S.; Candy, I.; Langdon, P.; Matthews, I.; Palmer, A.; Bayliss, A.; Milner, N. Climate, environment and Lake Flixton. In Star Carr Volume 1: A Persistent Place in a Changing World; Milner, N., Conneller, C., Taylor, B., Eds.; White Rose University Press: York, UK, 2018; pp. 41–54. [Google Scholar]
- Blockley, S. The Flixton area and Star Carr in the context of British Upper Palaeolithic and Early Mesolithic human occupation of Britain. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 32–41. [Google Scholar]
- Powell, J.H.; Ford, J.R.; Riding, J.B. Diamicton from the Vale of Pickering and Tabular Hills, north-east Yorkshire: Evidence for a Middle Pleistocene (MIS8) glaciation. Proc. Geol. Soc. 2016, 127, 575–594. [Google Scholar] [CrossRef] [Green Version]
- Bateman, M.D.; Evans, D.J.A.; Buckland, P.C.; Connell, E.R.; Friend, R.J.; Hartmann, D.; Moxon, H.; Fairburn, W.A.; Panagiotakopulu, E.; Ashurst, R.A. Last glacial dynamics of the Vale of York and North Sea lobes of the British and Irish Ice Sheet. Proc. Geol. Assoc. 2015, 126, 712–730. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.J.A.; Clark, C.D.; Mitchell, W.A. The last British ice sheet: A review of the evidence utilised in the compilation of the Glacial Map of Britain. Earth-Sci. Rev. 2005, 70, 253–312. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.J.A.; Bateman, M.D.; Roberts, D.H.; Medialdea, A.; Hayes, L.; Duller, G.A.T.; Fabel, D.; Clark, C.D. Glacial Lake Pickering: Stratigraphy and chronology of a proglacial lake dammed by the North Sea Lobe of the British-Irish ice sheet. J. Quat. Sci. 2017, 32, 295–310. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.R.; Hughes, L.; Burke, H.F.; Lee, J.R. The Vale of Pickering: An Initial Summary of the Quaternary/Superficial Geology and Data Holdings; British Geological Survey Open Report OR/15/064; British Geological Survey: Nottingham, UK, 2015. [Google Scholar]
- Catt, J. The Quaternary of East Yorkshire and adjacent areas. In East Yorkshire Field Guide; Ellis, S., Ed.; Quaternary Research Association: Cambridge, UK, 1987; pp. 1–14. [Google Scholar]
- Lincoln, P.; Matthews, I.; Palmer, A.; Blockley, S. Seamer Basin. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 87–91. [Google Scholar]
- Lincoln, P.; Matthews, I.; Palmer, A.; Blockley, S. Palaeoenvironmental reconstructions from the southern extension of Wykeham Quarry. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 103–113. [Google Scholar]
- Lincoln, P.; Matthews, I.P.; Palmer, A.P.; Blockley, S.; Staff, R.A.; Candy, I. Hydroclimatic changes in the British Isles through the Last-Glacial-Interglacial Transition: Multiproxy reconstructions from the Vale of Pickering, NE England. Quat. Sci. Rev. 2020, 249, 106630. [Google Scholar] [CrossRef]
- Matthews, I.; Abrook, A.; Lincoln, P.; Palmer, A. The Last Glacial Interglacial Transition (16–8 ka BP) in North East England. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 23–31. [Google Scholar]
- Palmer, A.P.; Matthews, I.P.; Candy, I.; Blockley, S.P.E.; MacLeod, A.; Darvill, C.M.; Milner, N.; Conneller, C.; Taylor, B. The evolution of palaeolake Flixton and the environmental context of Star Carr, NE. Yorkshire: Stratigraphy and sedimentology of the Last Glacial-Interglacial Transition (LGIT) lacustrine sequences. Proc. Geol. Assoc. 2015, 126, 50–59. [Google Scholar] [CrossRef]
- Candy, I.; Farry, A.; Darvill, C.M.; Palmer, A.; Blockley, S.P.E.; Matthews, I.P.; MacLeod, A.; Deeprose, L.; Farley, N.; Kearney, R.; et al. The evolution of Palaeolake Flixton and the environmental context of Star Carr: An oxygen and carbon isotopic record of environmental change for the early Holocene. Proc. Geol. Assoc. 2015, 126, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Day, S.P. Preliminary results of high-resolution palaeoecological analyses at Star Carr. Cambr. Archaeol. J. 1993, 3, 129–133. [Google Scholar] [CrossRef]
- Brown, A.G.; Bradley, C.; Grapes, T.; Boomer, I. Hydrological assessment of Star Carr and the Hertford catchment, Yorkshire, UK. J. Wetl. Archaeol. 2011, 11, 36–55. [Google Scholar] [CrossRef]
- Milner, N.; Conneller, C.; Elliott, B.; Koon, B.; Panter, H.; Penkman, K.; Taylor, B.; Taylor, M. From riches to rags: Organic deterioration at Star Carr. J. Archaeol. Sci. 2011, 38, 2818–2832. [Google Scholar] [CrossRef]
- Albert, B.M.; Innes, J.B.; Blackford, J.J.; Taylor, B.; Conneller, C.; Milner, N. Degradation of the wetland sediment archive at Star Carr: An assessment of current palynological preservation. J. Arch. Sci. Rep. 2016, 6, 488–495. [Google Scholar] [CrossRef] [Green Version]
- Cummins, G.E.; Simmons, I.G. Palaeoecological context of a Bos skeleton from peats at Flixton School House Farm, Vale of Pickering. FORUM Yorks. New Ser. 2013, 2, 21–28. [Google Scholar]
- Taylor, B.; Gray Jones, A. Definitely a pit, possibly a house? Recent excavations at Flixton School House Farm in the Vale of Pickering. Mesolith. Misc. 2009, 20, 21–26. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell: Oxford, UK, 1991. [Google Scholar]
- Stockmarr, J. Tablets with spores used in absolute pollen analysis. Pollen Spores 1971, 13, 871–873. [Google Scholar]
- Edwards, K.J. The separation of Corylus and Myrica pollen in modern and fossil samples. Pollen Spores 1981, 23, 205–218. [Google Scholar]
- Grimm, E.C. TILIA Software; Illinois State Museum: Springfield, MA, USA, 1993. [Google Scholar]
- Grimm, E.C. TGview; Illinois State Museum: Springfield, MA, USA, 2004. [Google Scholar]
- Troels-Smith, J. Characterisation of unconsolidated sediments. Danm. Geol. Unders. Række IV 1955, 3, 1–73. [Google Scholar]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk Ramsey, C.; Grootes, P.M.; Guilderson, T.P.; Haflidason, H.; Hajdas, I.; et al. IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 Years cal. BP. Radiocarbon 2013, 55, 1869–1877. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, G.L.; Bradshaw, R.H.W. The selection of sites for palaeovegetational studies. Quat. Res. 1981, 16, 80–96. [Google Scholar] [CrossRef]
- Beckett, S.C. Pollen diagrams from Holderness, North Humberside. J. Biogeogr. 1981, 8, 177–198. [Google Scholar] [CrossRef]
- Björk, S.; Walker, M.J.C.; Cwynar, L.C.; Johnsen, S.; Knudsen, K.-L.; Lowe, J.J.; Wohlfarth, B. Members INTIMATE An event stratigraphy for the Last Termination in the North Atlantic Region based on the Greenland Ice-Core Record: A proposal by the Intimate Group. J. Quat. Sci. 1998, 13, 283–292. [Google Scholar] [CrossRef]
- Walker, M.J.C.; Coope, G.R.; Lowe, J.J. The Devensian (Weichselian) Lateglacial palaeoenvironmental record from Gransmoor, East Yorkshire, England: A contribution to the ‘North Atlantic seaboard programme’ of IGCP-253, ‘Termination of the Pleistocene’. Quat. Sci. Rev. 1993, 12, 659–680. [Google Scholar] [CrossRef]
- Lowe, J.J.; Ammann, B.; Birks, H.H.; Björck, S.; Coope, G.R.; Cwynar, L.; de Beaulieu, J.-L.; Mott, R.J.; Peteet, D.M.; Walker, M.J.C. Climatic changes in areas adjacent to the North Atlantic during the last glacial-interglacial transition (14-9 ka BP): A contribution to IGCP-253. J. Quat. Sci. 1994, 9, 185–198. [Google Scholar] [CrossRef]
- Jones, R.L. Late Quaternary vegetational history of the North York Moors IV: Seamer Carrs. J. Biogeogr. 1976, 3, 397–406. [Google Scholar] [CrossRef]
- Walker, M.J.C. Climatic changes in Europe during the Last Glacial/Interglacial Transition. Quat. Int. 1995, 28, 63–76. [Google Scholar] [CrossRef]
- Walker, M.J.C. Quaternary Dating Methods; Wiley: Chichester, UK, 2005. [Google Scholar]
- Innes, J.B. The Late-glacial record of northern England: Introduction. In Quaternary of Northern England. Geological Conservation Review; Huddart, D., Glasser, N.F., Eds.; Joint Nature Conservation Review Committee: Peterborough, UK, 2002; Volume 25, pp. 211–220. [Google Scholar]
- Innes, J.B.; Bridgland, D.R.; Mitchell, W.A.; Rutherford, M.M.; O’Brien, C.; Long, A.J. Late Devensian environments in the Vale of Mowbray, North Yorkshire, UK: Evidence from palynology. Proc. Geol. Assoc. 2009, 120, 199–208. [Google Scholar] [CrossRef]
- Gearey, B.R.; Lillie, M.C. Routh Quarry. In The Quaternary of East Yorkshire and North Lincolnshire. Field Guide; Bateman, M.D., Buckland, P.C., Frederick, C.D., Whitehouse, N.J., Eds.; Quaternary Research Association: London, UK, 2001; pp. 69–72. [Google Scholar]
- Moore, J.W. Mesolithic sites in the neighbourhood of Flixton, north-east Yorkshire. Proc. Prehist. Soc. 1950, 16, 101–108. [Google Scholar] [CrossRef]
- Milner, N.; Taylor, B.; Conneller, C. Excavations at Flixton Island. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 140–147. [Google Scholar]
- Innes, J.B. Introduction to the Holocene (Flandrian) history and record of Northern England. In Quaternary of Northern England; Huddart, D., Glasser, N.F., Eds.; Geological Conservation Review Series 25; Joint Nature Conservation Committee: Peterborough, UK, 2002; pp. 351–365. [Google Scholar]
- Parker, A.G.; Goudie, A.S.; Anderson, D.E.; Robinson, M.A.; Bonsall, C. A review of the mid-Holocene elm decline in the British Isles. Prog. Phys. Geogr. 2002, 26, 1–45. [Google Scholar] [CrossRef]
- Bridgland, D.R.; Innes, J.B.; Long, A.J.; Mitchell, W.A. Late Quaternary Landscape Evolution of the Swale-Ure Washlands, North Yorkshire; Oxbow Books: Oxford, UK, 2011. [Google Scholar]
- Keen, D.H.; Jones, R.L.; Evans, R.A.; Robinson, J.E. Faunal and floral assemblages from Bingley Bog, West Yorkshire and their significance for Late Devensian and early Flandrian environmental changes. Proc. Yorks. Geol. Soc. 1988, 47, 125–138. [Google Scholar] [CrossRef]
- Young, D.S.; Green, C.P.; Batchelor, C.R.; Austin, P.; Elias, S.A.; Athersuch, J.; Lincoln, P. Macrofossil evidence of alder (Alnus sp.) in Britain early in the Late Glacial Interstadial: Implications for the northern cryptic refugia debate. J. Quat. Sci. 2021, 36, 40–55. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Emerson, B.C.; Hewitt, G.M. Cryptic or mystic? Glacial tree refugia in northern Europe. TREE 2013, 28, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.L. Late Devensian deposits from Kildale, north-east Yorkshire. Proc. Yorks. Geol. Soc. 1977, 41, 185–188. [Google Scholar] [CrossRef]
- Jones, R.L. Late Quaternary vegetational history of the North York Moors. V. The Cleveland Dales. J. Biogeogr. 1977, 4, 353–362. [Google Scholar] [CrossRef]
- Giles, J.R.A. Late Devensian and early Flandrian environments at Dishforth Bog, north Yorkshire. Proc. Yorks. Geol. Soc. 1992, 49, 1–10. [Google Scholar] [CrossRef]
- Hughes, P.D.M.; Mauquoy, D.; Barber, K.E.; Langdon, P.G. Mire-development pathways and palaeoclimatic records from a full Holocene peat archive at Walton Moss, Cumbria, England. Holocene 2000, 10, 465–479. [Google Scholar] [CrossRef]
- Bunting, M.J.; Gaillard, M.-J.; Sugita, S.; Middleton, R.; Broström, A. Vegetation structure and pollen source area. Holocene 2004, 14, 651–660. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W. Modern pollen representation factors for woods in south-east England. J. Ecol. 1981, 69, 45–70. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W. Spatial scale in the pollen record. In Modelling Ecological Change; Harris, D.R., Thomas, K.D., Eds.; Institute of Archaeology, University of London: London, UK, 1991; pp. 41–52. [Google Scholar]
- Sugita, S. Pollen representation of vegetation in Quaternary sediments: Theory and method in patchy vegetation. J. Ecol. 1994, 82, 881–897. [Google Scholar] [CrossRef]
- Bonny, A.P. The effect of pollen recruitment processes on pollen distribution over the sediment surface of a small lake in Cumbria. J. Ecol. 1978, 66, 385–416. [Google Scholar] [CrossRef]
- Pennington, W. The origin of pollen in lake sediments: An enclosed lake compared with one receiving inflow streams. New Phytol. 1979, 83, 189–213. [Google Scholar] [CrossRef]
- Davis, M.B. Pollen grains in lake sediments: Redeposition caused by seasonal water circulation. Science 1968, 162, 796–799. [Google Scholar] [CrossRef]
- Davis, M.B. Redeposition of pollen grains in lake sediments. Limnol. Oceanogr. 1973, 18, 44–52. [Google Scholar] [CrossRef]
- Pennington, W.; Brubaker, L.B. Differential sedimentation of pollen grains in lakes. Limnol. Oceanogr. 1973, 18, 635–646. [Google Scholar]
- Bonny, A.P. Recruitment of pollen to the seston and sediment of some lake district lakes. J. Ecol. 1976, 64, 859–887. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Bigler, M.; Blockley, S.P.; Blunier, T.; Buchardt, S.L.; Clausen, H.B.; Cvijanovic, I.; Dahl-Jensen, D.; Johnsen, S.J.; Fischer, H.; et al. A stratigraphic framework for abrupt climatic changes during the last glacial period based on three synchronized Greenland ice-core records: Refining and extending the INTIMATE event stratigraphy. Quat. Sci. Rev. 2014, 106, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.P.; Matthews, I.; Candy, I.; Blockley, S.; MacLeod, A.; Darvill, C.; Milner, N.; Conneller, C.; Taylor, B. The bathymetry and lithostratigraphy of sediments in Palaeolake Flixton, North Yorkshire. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 124–128. [Google Scholar]
- Tipping, R.M. The prospects for establishing synchroneity in the early postglacial pollen peak of Juniperus in the British Isles. Boreas 1987, 16, 155–163. [Google Scholar] [CrossRef]
- Chambers, F.M.; Elliott, L. Spread and expansion of Alnus Mill. In the British Isles: Timing, agencies and possible vectors. J. Biogeogr. 1989, 16, 541–550. [Google Scholar] [CrossRef]
- Bennett, K.D.; Birks, H.J.B. Postglacial history of alder (Alnus glutinosa (L.) Gaertn.) in the British Isles. J. Quat. Sci. 1990, 5, 123–133. [Google Scholar] [CrossRef]
- Bush, M.B.; Hall, A.R. Flandrian Alnus: Expansion or immigration? J. Biogeogr. 1987, 14, 479–481. [Google Scholar] [CrossRef]
- Chambers, F.M.; Price, S.-M. palaeoecology of Alnus (alder): Early postglacial rise in a valley mire, north-west Wales. New Phytol. 1985, 101, 333–344. [Google Scholar] [CrossRef]
- Croft, S.; Colonese, A.C.; Lucquin, A.; Craig, O.E.; Conneller, C.; Milner, N. Pine traces at Star Carr: Evidence on residues on stone tools. J. Archaeol. Sci. Rep. 2018, 21, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.D. Post-glacial dynamics of pine (Pinus sylvestris L.) and pinewoods in Scotland. In Our Pinewood Heritage; Aldhous, J.R., Ed.; Forestry Commission, The Royal Society for the Protection of Birds, Scottish Natural Heritage: Inverness, UK, 1995; pp. 23–29. [Google Scholar]
- Pigott, C.D.; Huntley, J.P. Factors controlling the distribution of Tilia cordata at the northern limits of its geographical range. II. History in North-West England. New Phytol. 1980, 84, 145–164. [Google Scholar] [CrossRef]
- Griffiths, S. A Bayesian radiocarbon chronology of the early Neolithic of Yorkshire and Humberside. Archaeol. J. 2014, 171, 2–29. [Google Scholar] [CrossRef]
- Bartley, D.D.; Chambers, C.; Hart-Jones, B. The vegetational history of parts of south and east Durham. New Phytol. 1976, 77, 437–488. [Google Scholar] [CrossRef]
- Simmons, I.G.; Innes, J.B. The ecology of an episode of prehistoric cereal cultivation on the North York Moors, England. J. Archaeol. Sci. 1996, 23, 613–618. [Google Scholar] [CrossRef]
- Innes, J.B.; Blackford, J.J.; Simmons, I.G. Late Mesolithic and early Neolithic forest disturbance: A high-resolution palaeoecological test of human impact hypotheses. Quat. Sci. Rev. 2013, 77, 80–100. [Google Scholar] [CrossRef]
- Innes, J.B.; Blackford, J.J. Palynology and the study of the Mesolithic-Neolithic transition in the British Isles. In The Archaeological and Forensic Applications of Microfossils: A Deeper Understanding of Human History; Williams, M., Hill, T., Boomer, I., Williamson, I.P., Eds.; Geological Society: London, UK, 2017; pp. 55–78. [Google Scholar]
- Birks, H.J.B.; Birks, H.H. The rise and fall of forests. Science 2004, 305, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Birks, H.J.B.; Tinner, W. Past forests of Europe. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Office EU: Luxembourg, 2016; pp. 36–39. [Google Scholar]
- Abrook, A.; Matthews, I.P.; Candy, I.; Palmer, A.P.; Francis, A.P.; Turner, L.; Brooks, S.J.; Self, A.E.; Milner, A.M. Complexity and asynchrony of climatic drivers and environmental responses during the last Glacial-Interglacial Transition (LGIT) in northwest Europe. Quat. Sci. Rev. 2020, 250, 106634. [Google Scholar] [CrossRef]
- Clark, C.D.; Gibbard, P.L.; Rose, J. Pleistocene glacial limits in England, Scotland and Wales. In Quaternary Glaciations—Extent and Chronology: Part 1: Europ; Ehlers, J., Gibbard, P.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 47–82. [Google Scholar]
- Pennington, W. Lags in adjustment of vegetation to climate caused by the pace of soil development: Evidence from Britain. Vegetatio 1986, 67, 105–118. [Google Scholar] [CrossRef]
- Feurdean, A.; Bhagwat, S.A.; Willis, K.J.; Birks, H.J.B.; Lischke, H.; Hickler, T. Tree migration-rates: Narrowing the gap between inferred post-glacial rates and projected rates. PLoS ONE 2013, 8, e71797. [Google Scholar] [CrossRef]
- Lowe, J.J.; Coope, R.G.; Lemdahl, G.; Walker, M.J.C. The Younger Dryas climate signal in land records from NW Europe. In The Younger Dryas; Troelstra, S.R., van Hinte, J.E., Ganssen, G.M., Eds.; Koninklijke Nederlandse Akademie van Wetenschappen: Oxford, UK, 1995; pp. 3–25. [Google Scholar]
- Hunt, C.O.; Hall, A.R.; Gilbertson, D.D. The palaeobotany of the Late Devensian sequence at Skipsea Withow Mere. In Late Quaternary Environments and Man in Holderness; Gilbertson, D.D., Ed.; British Archaeological Reports, British Series; BAR Publishing: Oxford, UK, 1984; pp. 81–108. [Google Scholar]
- Bartley, D.D. The stratigraphy and pollen analysis of lake deposits near Tadcaster, Yorkshire. New Phytol. 1962, 61, 277–287. [Google Scholar] [CrossRef]
- Coope, G.R.; Lemdahl, G.; Lowe, J.J.; Walking, A. Temperature gradients in northern Europe during the last glacial-Holocene transition (14–9 14C kyr BP) interpreted from coleopteran assemblages. J. Quat. Sci. 1998, 13, 419–433. [Google Scholar] [CrossRef]
- Jones, R.T.; Marshall, J.D.; Crowley, S.F.; Bedford, A.; Richardson, N.; Bloemendal, J.; Oldfield, F. A high resolution, multiproxy Late-glacial record of climate change and intrasystem responses in northwest England. J. Quat. Sci. 2002, 17, 329–340. [Google Scholar] [CrossRef]
- Bedford, A.; Jones, R.T.; Lang, B.; Brooks, S.; Marshall, J.D. A Late-glacial chironomid record from Hawes Water, northwest England. J. Quat. Sci. 2004, 19, 281–290. [Google Scholar] [CrossRef]
- Elias, S.A.; Matthews, I.P. A comparison of reconstructions based on aquatic and terrestrial beetle assemblages: Late glacial to Early Holocene temperature reconstructions for the British Isles. Quat. Int. 2014, 341, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Edwards, K.J.; Whittington, G.; Tipping, R.M. The incidence of microscopic charcoal in late glacial deposits. Palaeogeogr. Palaeoclimat. Palaeoecol. 2000, 164, 247–262. [Google Scholar] [CrossRef]
- Blockley, S.; Candy, I.; Matthews, I.; Langdon, P.; Palmer, A.; Lincoln, P.; Abrook, A.; Taylor, B.; Conneller, C.; Bayliss, A.; et al. The resilience of postglacial hunter-gatherers to abrupt climate change. Nat. Ecol. Evol. 2018, 2, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.J.; Birks, H.H.; Brooks, S.J.; Coope, G.R.; Harkness, D.D.; Mayle, F.E.; Sheldrick, C.; Turney, C.S.M.; Walker, M.J.C. The chronology of palaeoenvironmental changes during the last glacial-Holocene transition: Towards an event stratigraphy for the British Isles. J. Geol. Soc. Lond. 1999, 156, 397–410. [Google Scholar] [CrossRef]
- Marshall, J.D.; Jones, R.T.; Crowley, S.F.; Oldfield, F.; Nash, S.; Bedford, A. A high-resolution Lateglacial isotopic record from Hawes Water, Northwest England: Climatic oscillations, calibration and comparison of palaeotemperature proxies. Palaeogeogr. Palaeoclimat. Palaeoecol. 2002, 185, 25–40. [Google Scholar] [CrossRef]
- Lowe, J.J.; Rasmussen, S.O.; Bjorck, S.; Hoek, W.Z.; Steffensen, J.P.; Walker, M.J.C.; Yu, Z.C.; Grp, I. Synchronisation of palaeoenvironmental events in the North Atlantic region during the Last Termination: A revised protocol recommended by the INTIMATE group. Quat. Sci. Rev. 2008, 27, 6–17. [Google Scholar] [CrossRef]
- Candy, I.; Palmer, A.; Blockley, S.; Matthews, I.; MacLeod, A.; Farley, N.; Farry, A.; Kearney, R.; Abrook, A.; Darvill, C. δ13C and δ18O analysis of lacustrine marls from palaeolake Flixton. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 135–139. [Google Scholar]
- Lincoln, P.; Matthews, I.; Palmer, A.; Blockley, S. Synthesis of records from Wykeham Quarry. In The Quaternary of the Vale of Pickering: Field Guide; Lincoln, P., Eddey, L., Matthews, I., Palmer, A., Bateman, M., Eds.; Quaternary Research Association: London, UK, 2017; pp. 114–123. [Google Scholar]
- Mitchell, F.J.G. How open were European primeval forests? J. Ecol. 2005, 93, 168–177. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Vinther, B.M.; Clausen, H.B.; Andersen, K.K. Early Holocene climate oscillations recorded in three Greenland ice cores. Quat. Sci. Rev. 2007, 26, 1907–1914. [Google Scholar] [CrossRef] [Green Version]
- van der Plicht, J.; van Geel, B.; Bohncke, S.J.P.; Bos, J.A.A.; Blaauw, M.; Speranza, A.O.M.; Muscheler, R.; Björck, S. The Preboreal climate reversal and a subsequent solar-forced climate shift. J. Quat. Sci. 2004, 19, 263–269. [Google Scholar] [CrossRef]
- Fleitmann, D.; Mudelsee, M.; Burns, S.J.; Bradley, R.S.; Kramers, J.; Matter, A. Evidence for a widespread climatic anomaly at around 9.2 ka before present. Paleoceanography 2008, 23, PA1102. [Google Scholar] [CrossRef]
- Björck, S.; Rundgren, M.; Ingolfsson, O.; Funder, S. The Preboreal oscillation around the Nordic Seas: Terrestrial and lacustrine responses. J. Quat. Sci. 1997, 12, 455–466. [Google Scholar] [CrossRef]
- Björk, S.; Muscheller, R.; Kromer, B.; Andresen, C.S.; Heinemeier, J.; Johnsen, S.J.; Conley, D.; Koç, N.; Spurk, M.; Veski, S. High-resolution analyses of an early Holocene climatic event may imply decreased solar forcing as an important trigger. Geology 2001, 29, 1107–1110. [Google Scholar] [CrossRef]
- Conneller, C.; Higham, T. The early Mesolithic colonization of Britain: Preliminary results. In No Stone Unturned: Papers in Honour of Roger Jacobi; Ashton, N., Harris, C.R.E., Eds.; Lithic Studies Society: London, UK, 2015; pp. 157–166. [Google Scholar]
- Edwards, K.J.; Langdon, P.G.; Sugden, H. Separating climatic and possible human impacts in the early Holocene: Biotic response around the time of the 8200 cal. BP event. J. Quat. Sci. 2007, 22, 77–84. [Google Scholar] [CrossRef]
- Ghilardi, B.; O’Connell, M. Early Holocene vegetation and climate dynamics with particular reference to the 8.2 ka event: Pollen and macrofossil evidence from a small lake in western Ireland. Veget. Hist. Archaeobot. 2013, 22, 99–114. [Google Scholar] [CrossRef]
- Wicks, K.; Mithen, S. The impact of the abrupt 8.2 ka cold event on the Mesolithic population of western Scotland: A Bayesian chronological analysis using ‘activity events’ as a population proxy. J. Archeol. Sci. 2014, 45, 240–269. [Google Scholar] [CrossRef]
- Griffiths, S.; Robinson, E. The 8.2 ka bp Holocene climate change event and human population resilience in northwest Atlantic Europe. Quat. Int. 2018, 465, 251–257. [Google Scholar] [CrossRef]
- Waddington, C.; Wicks, K. Resilience or wipe out? Evaluating the convergent impacts of the 8.2 ka event and Storegga tsunami on the Mesolithic of northeast Britain. J. Archeol. Sci. Rep. 2017, 14, 692–714. [Google Scholar] [CrossRef]
- Smith, A.G. Problems of inertia and threshold related to post-glacial habitat changes. Proc. Roy. Soc. Lond. B 1965, 161, 331–342. [Google Scholar]
- Von Holle, B.; Delcourt, H.R.; Simberloff, D. The importance of biological inertia in plant community resistance to invasion. J. Veget. Sci. 2003, 14, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Hibbert, F.A.; Switsur, V.R.; West, R.G. Radiocarbon dating of Flandrian pollen zones at Red Moss, Lancashire. Proc. R. Soc. Lond. B Biol. Sci. 1971, 177, 161–176. [Google Scholar]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps For Europe: 0-13,000 Years Ago; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Birks, H.J.B. Holocene isochrone maps and patterns of tree-spreading in the British Isles. J. Biogeogr. 1989, 16, 503–540. [Google Scholar] [CrossRef]
- Huntley, B. Glacial and Holocene vegetation history: Europe. In Vegetation History; Huntley, B., Webb, T., III, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 341–383. [Google Scholar]
- Huntley, B. European vegetation history: Palaeovegetation maps from pollen data—13 000 yr BP to present. J. Quat. Sci. 1990, 5, 103–122. [Google Scholar] [CrossRef]
- Huntley, B. The post-glacial history of British woodlands. In Woodland in the Landscape: Past and Future Perspectives; Atherden, M., Butlin, R.A., Eds.; PLACE Research Centre: York, UK, 1998; pp. 9–25. [Google Scholar]
- Linnman, G. Some aspects of the colonization of Corylus avellana L. in north-west Europe during early Flandrian times. Striae 1981, 14, 72–75. [Google Scholar]
- Huntley, B. Rapid early-Holocene migration and high abundance of hazel (Corylus avellana L.): Alternative hypotheses. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman and Hall: London, UK, 1993; pp. 205–216. [Google Scholar]
- Tallantire, P.A. The early-Holocene spread of hazel (Corylus avellana L.) in Europe north and west of the Alps: An ecological hypothesis. Holocene 2002, 12, 81–96. [Google Scholar] [CrossRef]
- Seppä, H.; Schurgers, G.; Miller, P.A.; Bjune, A.E.; Giesecke, T.; Kühl, N.; Renssen, H.; Salonen, J.S. Trees tracking a warmer climate: The Holocene range shift of hazel (Corylus avellana) in northern Europe. Holocene 2015, 25, 53–63. [Google Scholar] [CrossRef]
- Tipping, R.M. Microscopic charcoal records, inferred human activity and climate change in the Mesolithic of northernmost Scotland. In The Early Prehistory of Scotland; Pollard, T., Morrison, A.M., Eds.; Edinburgh University Press: Edinburgh, UK, 1996; pp. 39–61. [Google Scholar]
- Tipping, R.M.; Milburn, P. Mid-Holocene charcoal fall in southern Scotland—Temporal and spatial variability. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 164, 177–193. [Google Scholar] [CrossRef]
- Smith, A.G.; Pilcher, J.R. Radiocarbon dates and vegetational history of the British Isles. New Phytol. 1973, 72, 903–914. [Google Scholar] [CrossRef]
- Birks, H.J.B. Holocene (Flandrian) chronostratigraphy of the British Isles: A review. Striae 1982, 16, 99–105. [Google Scholar]
- Barber, K.E.; Chambers, F.M.; Maddy, D. Holocene palaeoclimates from peat stratigraphy: Macrofossil proxy climate records from three oceanic raised bogs in England and Ireland. Quat. Sci. Rev. 2003, 22, 521–539. [Google Scholar] [CrossRef]
- Gieseke, T.; Brewer, S. Notes on the postglacial spread of abundant European tree taxa. Veget. Hist. Archaeobot. 2018, 27, 337–349. [Google Scholar] [CrossRef]
- Langdon, P.G.; Barber, K.E.; Hughes, P.D.M. A 7500-year, peat-based palaeoclimatic reconstruction and evidence for an 1100-year cyclicity in bog surface wetness from Temple Hill Moss, Pentland Hills, SE Scotland. Quat. Sci. Rev. 2003, 22, 259–274. [Google Scholar] [CrossRef]
- Bennett, K.D.; Lamb, H.F. Holocene pollen sequences as a record of competitive interactions among tree populations. TREE 1988, 3, 141–144. [Google Scholar] [CrossRef]
- Brown, A.G. Clearances and clearings: Deforestation in Mesolithic/Neolithic Britain. Ox. J. Archaeol. 1997, 16, 133–146. [Google Scholar] [CrossRef]
- Conneller, C. Moving beyond sites: Mesolithic technology in the landscape. In Mesolithic Studies at the Beginning of the 21st Century; Milner, N., Woodman, P., Eds.; Oxbow: Oxford, UK, 2005; pp. 42–55. [Google Scholar]
- Taylor, B.; Milner, N.; Conneller, C. Dryland structures. In Star Carr Volume 1: A Persistent Place in a Changing World; Milner, N., Conneller, C., Taylor, B., Eds.; White Rose University Press: York, UK, 2018; pp. 57–121. [Google Scholar]
- Bamforth, M.; Taylor, M.; Taylor, B.; Robson, H.K.; Radini, A.; Milner, N. Wooden structures. In Star Carr Volume 1: A Persistent Place in a Changing World; Milner, N., Conneller, C., Taylor, B., Eds.; White Rose University Press: York, UK, 2018; pp. 69–122. [Google Scholar]
- Bishop, R.R.; Church, M.J.; Rowley-Conwy, P.A. Firewood, food and human niche construction: The potential role of Mesolithic hunter-gatherers in actively structuring Scotland’s woodlands. Quat Sci. Rev. 2015, 108, 51–75. [Google Scholar] [CrossRef] [Green Version]
- Patterson, W.A.; Edwards, K.J.; Maguire, D.J. Microscopic charcoal as a fossil indicator of fire. Quat. Sci. Rev. 1987, 6, 3–23. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a fire proxy. In Tracking Environmental Change Using Lake Sediments, Vol 3., Terrestrial, Algal, and Siliceous Indicators; Smol, J.P., Birks, H.J.B., Last, W.M., Eds.; Kluwer: Dordrecht, The Netherlands, 2001; pp. 75–97. [Google Scholar]
- Bennett, K.D.; Simonson, A.B.; Peglar, S.M. Fire and man in postglacial woodlands of eastern England. J. Archaeol. Sci. 1990, 17, 635–642. [Google Scholar] [CrossRef]
- Law, C. The uses and fire-ecology of reedswamp vegetation. In Star Carr in Context; Mellars, P.A., Dark, P., Eds.; McDonald Institute for Archaeological Research: Cambridge, UK, 1998; pp. 197–206. [Google Scholar]
- Cummins, G.E. Fire! Accidental or strategic use of fire in the early Mesolithic of the eastern Vale of Pickering. In Mesolithic Lifeways. Current Research in Britain and Ireland; Young, R., Ed.; Leicester Archaeology Monographs No. 7; University Of Leicester: Leicester, UK, 2000; pp. 75–84. [Google Scholar]
- Bos, J.A.A.; Urz, R. Late Glacial and early Holocene environment in the middle Lahn river valley (Hessen, central-west Germany) and the local impact of early Mesolithic people—pollen and macrofossil evidence. Veget. Hist. Archaeobot. 2003, 12, 19–36. [Google Scholar] [CrossRef]
- Bos, J.A.A.; van Geel, B.; Groenewoudt, B.J.; Lauwerier, R.C.G.M. Early Holocene environmental change, the presence and disappearance of early Mesolithic habitation near Zutphen (The Netherlands). Veget. Hist. Archaeobot. 2005, 15, 27–43. [Google Scholar] [CrossRef]
- Rowley-Conwy, P.A. To the upper lake: Star Carr revisited—by birchbark canoe. In Economic Zooarchaeology: Studies in Hunting, Herding and Early Agriculture; Rowley-Conwy, P.A., Serjeantson, D., Halstead, P., Eds.; Oxbow Books: Oxford, UK, 2017; pp. 197–207. [Google Scholar]
- Simmons, I.G. The Environmental Impact of Later Mesolithic Cultures; Edinburgh University Press: Edinburgh, UK, 1996. [Google Scholar]
- Bush, M.B. Early Mesolithic disturbance: A force on the landscape. J. Arch. Sci. 1988, 15, 453–462. [Google Scholar] [CrossRef]
- Moore, J. Forest fire and human interaction in the early Holocene woodlands of Britain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 164, 125–137. [Google Scholar] [CrossRef]
- Tweddle, J.C. Regional vegetational history. In The Quaternary of East Yorkshire and North Lincolnshire Field Guide; Bateman, M.D., Buckland, P.C., Frederick, C.D., Whitehouse, N.J., Eds.; Quaternary Research Association: London, UK, 2001; pp. 35–46. [Google Scholar]
- Barnett, C. The chronology of Early Mesolithic occupation and environment impact at Thatcham Reedbeds, southern England. In Chronology and Evolution within the Mesolithic of North-West Europe; Crombé, P., van Strydonck, M., Sergant, J., Boudin, M., Bats, M., Eds.; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2009; pp. 57–76. [Google Scholar]
- Grant, M.J.; Stevens, C.J.; Whitehouse, N.J.; Norcott, D.; Macphail, R.I.; Langdon, C.; Cameron, N.; Barnett, C.; Langdon, P.G.; Crowder, J.; et al. A palaeoenvironmental context for terminal Upper Palaeolithic and Mesolithic activity in the Colne Valley: Offsite records contemporary with occupation at Three Ways Wharf, Uxbridge. Environ. Arch. 2014, 19, 131–152. [Google Scholar] [CrossRef]
- Lewis, H.T. Fire technology and resource management in aboriginal North America and Australia. In Resource Managers: North American and Australian Hunter-Gatherers; Williams, N.M., Hunn, E.S., Eds.; Westview Press: Boulder, CO, USA, 1982; pp. 45–67. [Google Scholar]
- Scherjon, F.; Bakels, C.; MacDonald, K.; Roebroeks, W. Burning the land: An ethnographic study of off-site fire use by current and historically documented foragers and implications for the interpretation of past fire practices in the landscape. Curr. Anthropol. 2015, 56, 299–326. [Google Scholar] [CrossRef]
- Coughlan, M.R.; Magi, B.I.; Derr, K.M. A global analysis of hunter-gatherers, broadcast fire use, and lightning-fire-prone landscapes. Fire 2018, 1, 41. [Google Scholar] [CrossRef]
- Mellars, P. Fire ecology, animal populations and man: A study of some ecological relationships in prehistory. Proc. Prehist. Soc. 1976, 42, 15–45. [Google Scholar] [CrossRef]
- Holst, D. Hazelnut economy of early Holocene hunter-gatherers: A case study from Mesolithic Duvensee, northern Germany. J. Archaeol. Sci. 2010, 37, 2871–2880. [Google Scholar] [CrossRef]
- Innes, J.B.; Simmons, I.G. Disturbance and diversity: Floristic changes associated with episodes of pre Ulmus-decline woodland recession in north-east Yorkshire. In Archaeology and the Flora of the British Isles; Jones, M., Ed.; Oxford University Press: Oxford, UK, 1988; pp. 7–20. [Google Scholar]
- Innes, J.B.; Blackford, J.J.; Simmons, I.G. Woodland disturbance and possible land-use regimes during the Late Mesolithic in the English uplands: Pollen, charcoal and non-pollen palynomorph evidence from Bluewath Beck, North York Moors, UK. Veget. Hist. Archaeobot. 2010, 19, 439–452. [Google Scholar] [CrossRef]
- Innes, J.B.; Simmons, I.G. Mid-Holocene charcoal stratigraphy, fire history and palaeoecology at North Gill, North York Moors, UK. Palaeogeogr. Palaeoclimat. Palaeoecol. 2000, 164, 151–165. [Google Scholar] [CrossRef]
- Simmons, J.B.; Innes, J.B. Tree remains in a North York Moors peat profile. Nature 1981, 294, 76–78. [Google Scholar] [CrossRef]
- Simmons, I.G.; Innes, J.B. Late Quaternary vegetational history of the North York Moors. X. Investigations on East Bilsdale Moor. J. Biogeogr. 1988, 15, 299–324. [Google Scholar] [CrossRef]
- Simmons, I.G.; Innes, J.B. Mid-Holocene adaptations and Later Mesolithic forest disturbance in Northern England. J. Archaeol. Sci. 1987, 14, 385–403. [Google Scholar] [CrossRef]
- Jones, R.L. The activities of Mesolithic man: Further palaeobotanical evidence from north-east Yorkshire. In Geoarchaeology: Earth Science and the Past; Davidson, D.A., Shackley, M.L., Eds.; Duckworth: London, UK, 1976; pp. 355–367. [Google Scholar]
- Jacobi, R.M. Northern England in the eighth millennium BC: An essay. In The Early Post-Glacial Settlement of Northern Europe; Mellars, P., Ed.; Duckworth: London, UK, 1978; pp. 295–332. [Google Scholar]
- Bunting, M.J.; Tipping, R.M. Complex hydroseral vegetation succession and ‘dryland’ pollen signals: A case study from northwest Scotland. Holocene 2004, 14, 53–63. [Google Scholar] [CrossRef]
- Binney, H.A.; Waller, M.P.; Bunting, M.J.; Armitage, R.A. The interpretation of fen-carr pollen diagrams: The representation of the dry land vegetation. Rev. Palaeobot. Palynol. 2005, 134, 197–218. [Google Scholar] [CrossRef]
- Bunting, M.J.; Farrell, M. Seeing the wood for the trees: Recent advances in the reconstruction of woodland in archaeological landscapes using pollen data. Environ. Arch. 2018, 23, 228–239. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | UK Grid Ref. | Latitude/Longitude | Geographical Location |
|---|---|---|---|
| Lake Centre profile D | TA04918091 | 54.21319° N: 0.39278° W | lake centre south-east of No Name Hill |
| Profile NM | TA04008120 | 54.21598° N: 0.40662° W | adjacent to south edge of No Name Hill |
| Profile NAQ | TA04028061 | 54.21068° N: 0.40652° W | north-east corner of No name Hill |
| Profile NAZ | TA03988149 | 54.21859° N: 0.40683° W | north-west corner of No name Hill |
| Profile AK87 | TA03568104 | 54.21463° N: 0.41342° W | adjacent to Flixton Island, to west |
| Profile F1035 | TA03548107 | 54.21491° N: 0.41372° W | western edge of Flixton Island |
| Profile VPCG | TA02208080 | 54.21275° N: 0.43435° W | extreme west of Lake Flixton |
| Profile FS | TA04848013 | 54.20620° N: 0.39412° W | at southern edge of Lake Flixton |
| Profile QAA | TA05608000 | 54.20488° N: 0.38253° W | at south-eastern edge of Lake Flixton |
| Profile LAL | TA06108040 | 54.20837° N: 0.37472° W | western edge of Barry’s Island |
| Profile LAP | TA61308042 | 54.20854° N: 0.37425° W | northern edge of Barry’s Island |
| Depth (cm) | Lab Code | 14C Date (yr. BP) | Age Range a (cal. BP) | Mean Age a (cal. BP) |
|---|---|---|---|---|
| Regional profile D | ||||
| 202–209 | Beta-104479 | 5740 ± 50 | 6413–6426; 6436–6656 | 6535 ± 122 |
| 321–330 | Beta-104478 | 8370 ± 60 | 9149–9168; 9250–9522 | 9336 ± 186 |
| Profile NM | ||||
| 12–13 | Beta-86147 | 6160 ± 50 b | 6906–7175; 7221–7234 | 7070 ± 164 |
| 70–71.2 | Beta-86146 | 8250 ± 50 b | 9033–9053; 9081–9407 | 9220 ± 187 |
| 109–110 | Beta-86145 | 8610 ± 60 b | 9487–9704; 9724–9731 | 9609 ± 122 |
| 156–156.6 | Beta-86144 | 11,400 ± 60 b c | 13,135–13,397 | 13,265 ± 131 |
| 155–157.1 | Beta-86143 | 11,410 ± 60 b d | 13,141–13,406 | 13,274 ± 132 |
| Profile AK87 | ||||
| 2–4 | Hv-17821 | 5300 ± 85 | 5920–6223; 6230–6278 | 6099 ± 179 |
| 13–15 | Hv-18296 | 5990 ± 90 | 6637–7070; 7079–7157 | 6897 ± 260 |
| 74–76 | Hv-17822 | 8710 ± 215 | 9256–10,276 | 9766 ± 510 |
| 98–100 | Hv-17823 | 8745 ± 380 | 8769–10,789; 11,037–11,059 | 9914 ± 1145 |
| 114–116 | Hv-17824 | 9395 ± 215 | 10,182–11,234 | 10,708 ± 525 |
| 118–120 | Hv-17825 | 9255 ± 135 | 10,175–10,792; 10,967–11,065 | 10,620 ± 445 |
| 124–126 | Hv-17826 | 10,275 ± 125 | 11,411–11,550; 11,602–12,533 | 11,972 ± 561 |
| Profile F1035 | ||||
| 15–17 | Hv-17827 | 6815 ± 110 | 7484–7867; 7901–7924 | 7704 ± 220 |
| 28–30 | Hv-17828 | 8340 ± 105 | 9035–9050; 9087–9528 | 9281 ± 247 |
| 49–51 | OxA-3734 | 8930 ± 85 | 9744–9756; 9761–10,236 | 9990 ± 245 |
| Profile VPCG | ||||
| 35–37 | Hv-17829 | 8755 ± 210 | 9307–10,298; 10,333–10,372 | 9840 ± 532 |
| 40–42 | Hv-17830 | 8435 ± 195 | 8796–8907; 8978–10,127 | 9461 ± 665 |
| Profile NAQ | ||||
| 154–156 | Beta-104483 | 9810 ± 160 | 10,711–11,815 | 11,263 ± 552 |
| 164–166 | Beta-104482 | 9570 ± 130 | 10,565–11,224 | 10,895 ± 330 |
| Profile NAZ | ||||
| 9–11 | Beta-104486 | 8850 ± 50 e | 9737–10,169 | 9953 ± 216 |
| 34–37 | Beta-104485 | 9250 ± 60 f | 10,258–10,570 | 10,414 ± 156 |
| 46 | Beta-104484 | 9510 ± 60 g | 10,591–10,628; 10,649–11,091 | 10,841 ± 250 |
| Profile LAP | ||||
| 164–167 | Beta-94438 | 6140 ± 60 | 6809–6812; 6859–7239 | 7024 ± 215 |
| 182.5–185 | Beta-94437 | 8850 ± 50 | 9737–10,169 | 9953 ± 216 |
| 237–239 | Beta-94436 | 10,140 ± 100 h | 11,320–12,113 | 11,717 ± 396 |
| 258–259 | Beta-94435 | 11,740 ± 130 | 13,322–13,844 | 13,584 ± 261 |
| Profile FS | ||||
| 262–264 | Beta-104481 | 9020 ± 60 b | 9919–10,081; 10,116–10,272 | 10,096 ± 176 |
| 265–267 | Beta-104480 | 9030 ± 60 b | 9919–10,071; 10,117–10,366 | 10,142 ± 224 |
| 268–269 | Beta-94434 | 9220 ± 100 b | 10,219–10,607; 10,618–10,657 | 10,438 ± 219 |
| 282–984 | Beta-94433 | 9900 ± 100 b | 11,157–11,764 | 11,460 ± 304 |
| 298–299 | Beta-94432 | 10,230 ± 100 b | 11,410–11,435; 11,479–12,398 | 11,904 ± 494 |
| 303–305 | Beta-94431 | 11,430 ± 100 b | 13,106–13,481 | 13,294 ± 188 |
| c. 7200–5000 14C BP (Atlantic) |
|---|
| Alnus-Ulmus-Quercus-Corylus |
| Alnus is added to the tree flora. Its immigration is asynchronous varying from c. 7600 14C BP [28] to c. 6000 14C BP. Tilia appears late in the period. Vegetation disturbance is significant by the end of the mid-Holocene with heliophytic taxa forming a significant contribution to the local (and regional?) pollen rain. Late Mesolithic people were present and human occupation was probably an important factor affecting the development of the vegetation and in some places disturbance might be linked to the local rise of alder. The Ulmus Decline seems not to be recorded. Almost all of the former lake is covered in reed and/or sedge swamp and alder carr and only very small pockets of open water are left in the deepest areas. Microcharcoal is either absent or in very low frequencies and burning was not a significant influence in the damper, denser mid-Holocene forest. |
| c. 10,000–c.7200 14C BP (Pre-Boreal and Boreal) |
| Betula (c. 10,000–c.9000) Corylus (c. 9000–c.8000) Corylus-Pinus-Ulmus-Quercus (c. 8000–c. 7200) |
| Encroachment of lake edge vegetation and reed swamp was rapid and occurred from the earliest point of the Holocene. The importance of Salix and Populus carr is variable around the lake edge although its detection depends to a large extent on pollen preservation and sample location, relative to the water’s edge. After succession through Juniperus and Salix shrubs, Betula dominated the first Holocene millennium. Rapid replacement of Betula by Corylus occurred around 9000 14C BP, with hazel thickets established both locally and regionally. Pinus and Ulmus enter the woodland in places in the centuries leading up to 8000 14C BP. Immigration of Quercus is highly asynchronous, occurring as early as 8700 14C BP at Flixton Island (AK87) but well after 8000 14C BP elsewhere. Early Mesolithic people were present all around the lake and their occupation of the area is envisaged to be continuous throughout, although greatest in the Betula phase and later varying in intensity and location. Detection of vegetation disturbance occurs close to the cultural activity in the Betula phase and further afield during the later phases. Microcharcoal is common near cultural sites in the Betula phase, as well as at the time of the expansion of Corylus, with microcharcoal layers sometimes present as at profile VPCG. It is less prevalent in the later phases. |
| >10,000 14C BP (Lateglacial) |
| Poaceae and tundra herbs (>c.13,000) Betula-Poaceae (c.13,000–c.12,000) Juniperus-Poaceae-Betula (c.12,000) Betula (c.12,000–c.11,000) Poaceae-Cyperaceae-tundra herbs (11,000–10,000) |
| Organic sedimentation began as early as 13,000 14C BP, or even slightly earlier at profile D. Open, tundra-type vegetation was supplanted by open Betula woodland in the first part of the Lateglacial (Windermere) Interstadial. A reversion to Juniperus scrub and grassland occurred under colder conditions around 12,000 14C BP. Denser Betula woodland developed in the later Interstadial. After c.11,000 14C BP in the cold Lateglacial (Loch Lomond) Stadial grassland and tundra-type herbs dominated, with some localised scrub. Microcharcoal frequencies vary but are generally high throughout the Lateglacial, and fire (from whatever source) appears to have played an important part in the environment. Upper Palaeolithic people were present except for the very cold Lateglacial Stadial. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simmons, I.G.; Cummins, G.E.; Taylor, B.; Innes, J.B. Lateglacial to Mid-Holocene Vegetation History in the Eastern Vale of Pickering, Northeast Yorkshire, UK: Pollen Diagrams from Palaeolake Flixton. Quaternary 2022, 5, 52. https://doi.org/10.3390/quat5040052
Simmons IG, Cummins GE, Taylor B, Innes JB. Lateglacial to Mid-Holocene Vegetation History in the Eastern Vale of Pickering, Northeast Yorkshire, UK: Pollen Diagrams from Palaeolake Flixton. Quaternary. 2022; 5(4):52. https://doi.org/10.3390/quat5040052
Chicago/Turabian StyleSimmons, Ian G., Gaynor E. Cummins, Barry Taylor, and James B. Innes. 2022. "Lateglacial to Mid-Holocene Vegetation History in the Eastern Vale of Pickering, Northeast Yorkshire, UK: Pollen Diagrams from Palaeolake Flixton" Quaternary 5, no. 4: 52. https://doi.org/10.3390/quat5040052